Альбертозавр
| † Альбертозавр | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Клада: Клада: Клада: Клада: Клада: Клада: Клада: Клада: Надсемейство: Семейство: Подсемейство: Род: † Альбертозавр | |||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Albertosaurus Osborn, 1905 | |||||||||||||||||||||||||||||||||||||||||
| Единственный вид | |||||||||||||||||||||||||||||||||||||||||
Osborn, 1905 | |||||||||||||||||||||||||||||||||||||||||
| Геохронология 70,6—66,043 млн лет
◄ Наше время◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание◄ Массовоепермское вымирание ◄ Девонское вымирание◄ Ордовикско-силурийское вымирание◄ Кембрийский взрыв | |||||||||||||||||||||||||||||||||||||||||


Альбертоза́вр[1][2] (лат. Albertosaurus) — род тероподных динозавров из семейства тираннозаврид, обитавших в западной части Северной Америки (от Аляски до Техаса) во времена позднемеловой эпохи (70,6—66,043 млн лет назад[3]). Ареал типового вида Albertosaurus sarcophagus был ограничен областью современной канадской провинции Альберты, отсюда и произошло его название.
Как и все тираннозавриды, альбертозавр был двуногим хищником с небольшими двупалыми передними конечностями и массивной головой с десятками больших острых зубов. Считается, что альбертозавр был доминирующим хищником в своей среде обитания. Будучи относительно крупным для теропода, по размерам он значительно уступал своему более известному родственнику тираннозавру — средняя масса особей альбертозавра была менее двух тонн.
Изучение вида ведётся с 1884 года, всего были обнаружены ископаемые остатки более тридцати особей, что позволило учёным изучить анатомию альбертозавра лучше других представителей семейства. Открытие двадцати особей различного возраста на одном участке (что может являться свидетельством стайного поведения) позволяет исследовать онтогенез и популяционную биологию альбертозавра, в отличие от большинства других динозавров.
Описание
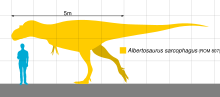
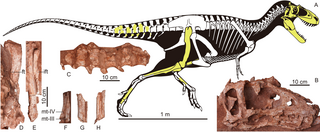
По размеру альбертозавр был меньше других тираннозаврид, например, тарбозавра и тираннозавра. Типичная взрослая особь достигала 9 метров в длину[4][5], некоторые старые особи могли превышать 10 метров[6]. Вес взрослых альбертозавров, вычисленный на основе ископаемых остатков разными методами, был от 1,2[7] до 1,7 тонны[8].
Массивный череп альбертозавра, достигавший 1 метра в длину, поддерживала короткая S-образная шея[9]. Широкие отверстия в черепе (окна) снижали вес головы, а также предоставляли пространство для челюстных мышц и сенсорных органов. Длинные челюсти альбертозавра содержали более 60 длинных, похожих на бананы зубов, более крупные тираннозавриды имели меньшее количество зубов. В отличие от большинства других теропод, альбертозавр и другие тираннозавриды были гетеродонтами, то есть имели разные виды зубов, в зависимости от их расположения во рту. Зубы, находившиеся на предчелюстной кости, были меньше остальных, располагались более тесно и имели D-образную форму в сечении[5]. Над глазами имелись короткие костяные гребни, которые при жизни могли быть яркого цвета и использоваться при ухаживании для привлечения партнёра[10].
Все тираннозавриды, в том числе альбертозавр, имеют сходное строение тела. Как большинство теропод, альбертозавр был бипедальным и уравновешивал тяжёлую голову и корпус очень длинным хвостом. Передние конечности тираннозаврид были чрезвычайно маленьких размеров и сохранили только два когтя. Задние конечности были длинными и заканчивались четырьмя пальцами с мощными когтями. Короткий первый палец задней конечности не участвовал в передвижении, только три других соприкасались с землёй, третий (средний) был больше, чем остальные[5]. Считается, что альбертозавр мог развивать скорость от 40 до 50 км/ч[10].
Систематика
Альбертозавр был представителем семейства тираннозаврид, подсемейства альбертозаврин. Его самый близкий «родственник» — обнаруженный в породах более позднего возраста горгозавр[11] (иногда называемый Albertosaurus libratus[12]). Эти два рода — единственные известные представители подсемейства альбертозаврин, но могли существовать и другие[13]. Палеонтолог Томас Хольц в 2004 году предположил, что аппалачиозавр тоже относился к альбертозавринам[5], но в более поздней его работе аппалачиозавр не рассматривался как представитель этого подсемейства[14], эта точка зрения поддерживается и другими учёными[12].
Бо́льшая же часть систематиков не выделяют подсемейство альбертозаврин[3], включая род непосредственно в семейство тираннозаврид, что подтверждено в 2014 году в исследовании Люя и других[15].
| Кладограмма тираннозаврид | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Другим подсемейством тираннозаврид являются тираннозаврины, к которым могут относиться дасплетозавр, тарбозавр и тираннозавр. По сравнению с тираннозавринами, альбертозавр имел более стройное тело, пропорционально меньший череп и кости таза и более длинные большеберцовые и таранные кости, а также кости плюсны и пальцев[9][11].
История изучения
Этимология названия
Название этому виду динозавров дал известный американский геолог, евгеник и палеонтолог Генри Осборн в 1905 году в одностраничном комментарии в конце своего описания Tyrannosaurus rex. Название «альбертозавр» происходит от названия канадской провинции Альберта, на территории которой впервые были найдены кости. Родовое название также включает в себя греческое слово σαῦρος (saurus) — ящер. Название типового вида «sarcophagus» означает «хищник», «плотоядный» и имеет то же происхождение, что и слово «саркофаг» (от древнегреческого σάρξ (sarx) — плоть и φαγεῖν (phagein) — поедать)[16]. Науке известно более тридцати скелетов этого динозавра всех возрастов[6][13].
Открытие

Типовым экземпляром является неполный череп, обнаруженный в 1884 году в отложениях карьера Хорсшу одноимённого каньона, расположенного рядом с рекой Ред-Дир на территории современной провинции Альберта. Этот экземпляр и меньший череп, вместе с некоторым скелетным материалом, были обнаружены геологической экспедицией во главе с известным геологом Джозефом Тирреллом. Эти два черепа были отнесены к ранее описанному виду Laelaps incrassatus Эдвардом Коупом в 1892 году[17], несмотря на то, что название Laelaps было ранее дано роду клещей и было изменено на Dryptosaurus в 1877 году Отниелом Маршем. Коуп отказался признать новое название, созданное его главным научным противником; несмотря на это, Лоуренс Ламб в 1904 году описал эти черепа, отнеся их к виду Dryptosaurus incrassatus, а не Laelaps incrassatus[18]. Чуть позже Генри Осборн указал на то, что описание Dryptosaurus incrassatus было основано на нескольких основных зубах тираннозаврид, поэтому два черепа из Альберты не могут с уверенностью быть отнесены к этому виду. Эти черепа также значительно отличались от ископаемых остатков типового вида дриптозавра — Dryptosaurus aquilunguis. Таким образом, Осборн в 1905 году выделил на основании этих черепов новый вид — Albertosaurus sarcophagus. При классификации он полагался на описание, созданное Ламбом на год раньше[16]. Оба экземпляра (NMC 5600 и NMC 5601[note 1]) были выставлены в Канадском музее природы в Оттаве.
Экспедиция Брауна
В 1910 году американский палеонтолог Барнум Браун обнаружил множество костей, принадлежавших большой группе альбертозавров в другом карьере, рядом с рекой Ред-Дир. Из-за большого количества образцов и недостатка времени экспедиция Брауна не откопала все экземпляры, но всё же ей удалось собрать из костеносного слоя все особи, которые можно было идентифицировать на тот момент. В тот год было обнаружено семь разных наборов костей стопы, а также два образца, не соответствующих другим по размерам и, возможно, принадлежавших детёнышам. Все они находятся в коллекции Американского музея естественной истории. Королевский Тиррелловский палеонтологический музей возобновил экспедицию в этот район (Сухой остров Буффало-Джамп[англ.]) в 1997 году[19]. Раскопки проводились с 1997 по 2005 год. В результате были обнаружены кости ещё 13 особей, среди них двухлетнего детёныша и очень старого альбертозавра, длина которого была оценена в 10 метров. Ни один скелет из найденных экземпляров не был полным. Кости были выставлены в Тирелловском музее и Американском музее естественной истории[6][7].
Горгозавр

В 1913 году палеонтолог Чарльз Штернберг обнаружил скелет тираннозаврида в формации Парк динозавров в Альберте. В следующем 1914 году Лоуренс Ламб дал этому динозавру название Gorgosaurus libratus[20]. Позже в Альберте и американском штате Монтана были найдены другие ископаемые остатки горгозавров. Обнаружив, что горгозавр и альбертозавр имеют слишком мало отличий, Дейл Рассел предложил считать горгозавра младшим синонимом альбертозавра, а так как альбертозавр был открыт и назван первым, в 1970 году горгозавр был переименован в Albertosaurus libratus, из-за чего временной диапазон обитания альбертозавра увеличился на несколько миллионов лет, а ареал простирался на сотни километров южнее, чем это считалось раньше[4].
В 2003 году палеонтолог Фил Карри изучил черепа нескольких тираннозаврид, в том числе и горгозавра и альбертозавра, и выяснил, что они имеют всё же несколько больше различий, чем считалось ранее. На сегодняшний день вопрос о валидности этих таксонов, не решён окончательно, так как горгозавр и альбертозавр являются очень близкими таксонами[11]. Признавая это, Карри, однако, рекомендовал, чтобы альбертозавр и горгозавр считались отдельными родами, поскольку отличий между ними не меньше, чем между дасплетозавром и тираннозавром, которых почти всегда можно отличить друг от друга. Кроме того, недавно были обнаружены новые скелеты альбертозаврин в штате Нью-Мексико и на Аляске, и Карри предположил, что вопрос о альбертозавре-горгозавре может быть полностью разъяснён, как только они будут изучены[13]. Большинство исследователей поддерживают точку зрения Карри[5], но есть и противники этой гипотезы[12].
Другие находки
В 1928 году Уильям Паркс описал новый вид — Albertosaurus arctunguis на основании частичного скелета, раскопанного у реки Ред-Дир[21], но с 1970 года этот вид считается идентичным Albertosaurus sarcophagus[5]. Этот экземпляр (ROM 807) хранится в Королевском музее Онтарио в Торонто. С тех пор в Альберте было обнаружено ещё шесть экземпляров, отнесённых к этому виду. Ископаемые, причисленные к Albertosaurus sarcophagus, были также обнаружены в американских штатах Монтана, Нью-Мексико и Вайоминг[5], но они, вероятно, не относятся к этому виду, а возможно, и к роду альбертозавр[12].
Albertosaurus megagracilis был описан на основании небольшого скелета, найденного в геологической формации Хелл-Крик[22]. В 1995 году он был выделен в отдельный род — Dinotyrannus[23], который позже стали описывать как принадлежащий подростковой особи тираннозавра рекса[9].
Палеобиология
Модель роста

Альбертозавр —
До нашего времени сохранились ископаемые остатки почти всех возрастных групп альбертозавра. С помощью гистологического исследования можно определить возраст животного в момент его смерти. Сравнив остатки особей одного вида, но разного возраста, можно определить среднюю скорость роста этих животных. Самый молодой из известных альбертозавров — двухлетний, он был обнаружен в Баффало-Джамп, весил приблизительно 50 килограммов, а в длину достигал чуть более двух метров. Десятиметровый экземпляр, обнаруженный в том же карьере, является самым старым из известных — он умер в возрасте двадцати восьми лет. Наибольший темп роста альбертозавра приходился на период от четырёх до шестнадцати лет, как и у некоторых других тираннозаврид. В этот период жизни альбертозавры набирали 122 килограмма ежегодно, пока не достигали 1,3 тонны. Впрочем, согласно другим исследованиям, масса взрослых альбертозавров был больше, и темпы роста тоже могли быть больше. У тираннозаврид, близких по размерам к альбертозавру, темпы роста были схожи, а более крупные, такие как тираннозавр, наоборот, росли почти в пять раз быстрее (601 килограмм ежегодно)[6]. Конец фазы быстрого роста у альбертозавров предполагал достижение половой зрелости; хотя они и продолжали расти до конца жизни, но гораздо медленнее[6][7]. Раннее половое созревание типично для мелких[24] и крупных динозавров[25], как и для большинства млекопитающих, таких как люди и слоны[25]. С другой стороны, современные динозавры, птицы, достигают половой зрелости лишь по завершении процесса роста[25].
Жизненный цикл

Чаще всего встречаются ископаемые остатки особей альбертозавров в возрасте четырнадцати лет и старше. Более молодых животных находят редко по нескольким причинам — в основном из-за того что кости молодых животных сохраняются хуже, чем кости взрослых, а также из-за того, что искателям окаменелостей их сложнее заметить[26]. Несмотря на то, что молодые альбертозавры были достаточно крупных размеров, их остатков обнаружено гораздо меньше, чем взрослых. Предполагается, что такая ситуация сложилась вследствие хороших условий жизни для детёнышей, из-за чего они умирали реже взрослых[6].
Согласно общей теории, уже по достижении всего двух лет детёныши альбертозавра были крупнейшими хищниками своего ареала, они также были гораздо быстрее, чем все их жертвы. Из-за этого даже у молодых альбертозавров не было врагов. Это и привело к уменьшению смертности детёнышей и, соответственно, к уменьшению количества их остатков. Остатки двенадцатилетних альбертозавров встречаются в два раза чаще, чем более молодых, а достигших половой зрелости четырнадцатилетних и старше — ещё в два раза больше. Возможно, по достижении половой зрелости усиливалась внутривидовая борьба за самку или территорию. Более высокая смертность взрослых объясняет большее количество сохранившихся остатков. Очень крупные особи также встречаются очень редко, поскольку лишь немногим удавалось прожить столь долго, чтобы достигнуть таких размеров. Схема, в которой высокий уровень смертности наблюдается среди детёнышей, низкий среди подростков, повышенный среди взрослых особей и крайне небольшое количество животных способно дожить до глубокой старости, свойственна большинству современных крупных животных, таких как носороги, африканский буйвол или слон, а также некоторым тираннозавридам, за исключением, разумеется, альбертозавра, детёныши которого умирали редко. В связи с пробелами в палеонтологической летописи, выводы об уровне смертности не могут быть более точными, в том числе и потому что более двух третей всех остатков альбертозавров были найдены в одном месте[6][27].
Стайность


В Баффало-Джамп были обнаружены ископаемые остатки двадцати двух альбертозавров, что является самым крупным массовым захоронением теропод после массового захоронения аллозавров в Карьере динозавров Кливленд-Ллойд[англ.] в Юте. В захоронении присутствуют остатки одной старой особи, восьми взрослых между семнадцатью и двадцатью тремя годами, семи подростков в возрасте от двенадцати до шестнадцати лет и шести детёнышей между двумя и одиннадцатью годами, которые ещё не достигли фазы быстрого роста[6].
Почти полное отсутствие остатков растительноядных животных и одинаковый уровень сохранности остатков альбертозавров в карьере навели Фила Карри на мысль, что это захоронение не было ловушкой для хищников, как смоляные ямы на ранчо Ла-Брея в Калифорнии, и что все сохранившиеся животные умерли в одно время. Карри считает, что это свидетельство стайного поведения[19]. Другие учёные более скептичны, отмечая, что, возможно, животных согнала вместе засуха или их тела были снесены в одно место наводнением[6][26].
Есть многочисленные свидетельства стадного образа жизни среди растительноядных динозавров, в том числе цератопсов и гадрозавров[28]. Однако массовые захоронения хищных динозавров редки. Маленькие тероподы, такие как дейноних[29], целофиз и мегапнозавр (синтарс)[30], были найдены в больших скоплениях, как и крупные хищники аллозавр и мапузавр[31]. Есть также доказательства стайного поведения среди других тираннозаврид. Фрагментарные остатки детёнышей были найдены рядом с тираннозавром по прозвищу «Сью», хранящимся в музее естественной истории имени Филда в Чикаго, а в костеносном слое геологической формации Ту-Медисин в Канаде были найдены ископаемые остатки трёх дасплетозавров рядом с несколькими гадрозаврами[32]. Эти находки могут подтвердить, что альбертозавр также вёл социальный образ жизни, хотя некоторые вышеупомянутые скопления могут быть временными или неестественными[19]. Но этому может быть и другое объяснение. По крайней мере, некоторые из этих находок представляют собой проявления массового каннибализма, как у комодского варана[26].
Карри также считает, что доказательством стайного образа жизни альбертозавров может быть их способ охоты. Пропорции ног молодых альбертозавров были сопоставимы с таковыми у орнитомимид, которые, пожалуй, были самыми быстрыми среди динозавров. Они, по всей видимости, были быстрее любых растительноядных своего времени. Карри выдвинул гипотезу, что младшие члены стаи отбивали жертву от стада и загоняли её к взрослым особям, которые были сильнее детёнышей, но медлительней[19]. Животные подросткового возраста, предположительно, вели образ жизни, отличавшийся от поведения своих взрослых родственников, заполняя экологическую нишу между ними и мелкими тероподами, крупнейший из которых весил вдвое меньше взрослого альбертозавра[5]. Подобная ситуация наблюдается у современных комодских варанов, которые начинают жизнь как маленькие насекомоядные хищники, а потом становятся доминирующими хищниками[33]. Тем не менее, полностью воссоздать поведение вымерших животных на основании окаменелостей невозможно.

Все идентифицированные окаменелости Albertosaurus sarcophagus были обнаружены в формации каньона Хорсшу. Эту геологическую формацию датируют маастрихтским ярусом верхнемеловой системы, возрастом 73—70 миллиона лет. Непосредственно ниже этой формации находятся сланцы морской формации Бирпо, которая является частью более обширной морской формации Сиуэй. В конце мелового периода море отступало, климат становился холоднее, уровень моря понижался, обнажая землю, которая раньше находилась под водой. Этот процесс, однако, не был постоянным: из-за сильных бурь на море часть суши в районе каньона Хорсшу частично затоплялась, но в конце концов море всё-таки отступило. Из-за постоянно изменяющегося уровня моря в формации Хорсшу было много прибрежных лагун, болот и илистых осушек. Многочисленные угольные пласты представляют древние торфяные болота. Как и в случае большинства других позвоночных в этой формации, ископаемые остатки альбертозавров были найдены в отложениях доисторических дельт и пойм больших рек[34].
Доисторическая фауна формации Хорсшу хорошо известна, поскольку окаменелости позвоночных, включая динозавров, присутствуют здесь в массовых количествах. Ихтиофауну представляли акулы, скаты, осетры, ильные рыбы, панцирникообразные и подобные панцирниковым аспидохиноморфы. Фауна млекопитающих включала в себя многобугорчатых и сумчатых, таких как дидельфодон. В морских отложениях был обнаружен плезиозавр леуроспондил, а в отложениях пресноводной среды были найдены черепахи, хампсозавры и крокодилы, такие как лейдиозух и стангерохампс. Доминирующими представителями фауны были динозавры, особенно распространены были гадрозавры, которые составляют половину всех найденных здесь динозавров, к ним относились эдмонтозавр, зауролоф и гипакрозавр. Цератопсы и орнитомимиды также были очень распространены, составляя треть найденных здесь динозавров. Наряду с намного более редко встречающимися анкилозаврами и пахицефалозаврами, эти растительноядные динозавры были добычей для множества плотоядных теропод, таких как троодонтиды, дромеозавриды и ценагнатиды. Взрослые альбертозавры был доминирующими хищниками в этой среде и находились на вершине пищевой пирамиды, а промежуточные ниши между мелкими тероподами и взрослыми альбертозаврами, возможно, занимали юные альбертозавры[34].
В массовой культуре
- Альбертозавр показан в последнем эпизоде сериала ВВС «Доисторический парк».
- Альбертозавр присутствует и в мультфильме «Поход динозавров».
- Альбертозавр фигурирует в сериале «Портал юрского периода: Новый мир».
- Альбертозавр присутствует в кроссплатформенной on-line игре Jurassic Park Builder.
- Альбертозавр присутствует в игре Jurassic World Alive.
Примечания
Комментарии
- ↑ Первые буквы — сокращение, образованное первыми буквами названия музея на языке оригинала, в котором хранится окаменелость, а цифровой индекс — номер этой окаменелости в коллекции музея.
Источники
- ↑ Бейли Дж., Седдон Т. Доисторический мир / П. К. Чудинов. — Росмэн, 1995. — С. 39. — 160 с. — ISBN 5-7519-0097-9.
- ↑ Хоун Д. Хроники тираннозавра: Биология и эволюция самого известного хищника в мире = The Tyrannosaur Chronicles: The Biology of the Tyrant Dinosaurs : ориг. изд. 2016 : [пер. с англ.] / науч. ред. А. Аверьянов. — М. : Альпина нон-фикшн, 2017. — С. 69. — 358 с. : ил. — ISBN 978-5-91671-744-0.
- ↑ 1 2 Albertosaurus (англ.) информация на сайте Paleobiology Database. (Дата обращения: 29 июня 2019).
- ↑ 1 2 Dale A. Russell. Tyrannosaurs from the Late Cretaceous of western Canada (англ.) // National Museum of Natural Sciences Publications in Paleontology. — 1970. — P. 1—34.
- ↑ 1 2 3 4 5 6 7 8 Thomas R. Holtz. Tyrannosauroidea // The Dinosauria / David B. Weishampel, Peter Dodson & Halszka Osmólska. — 2. — Berkeley, CA: University of California Press, 2004. — P. 111—136. — 861 p. — ISBN 0-520-24209-2.
- ↑ 1 2 3 4 5 6 7 8 9 Gregory M. Erickson; Philip. J. Currie; Brian D. Inouye; Alice A. Wynn. Tyrannosaur life tables: an example of nonavian dinosaur population biology (англ.) // Science. — AAAS, 2006. — Vol. 313. — P. 213—217. — ISSN 0036-8075. — doi:10.1126/science.1125721. Архивировано 28 февраля 2009 года.
- ↑ 1 2 3 Gregory M. Erickson; Peter J. Makovicky; Philip J. Currie; Mark A. Norell; Scott A. Yerby & Christopher A. Brochu. Gigantism and Comparative Life-History Parameters of Tyrannosaurid Dinosaurs (англ.) // Nature. — NPG, 2004. — Vol. 430. — P. 772—775. — ISSN 0028-0836. — doi:10.1038/nature02699. Архивировано 5 сентября 2009 года.
- ↑ Per Christiansen & Richard A. Fariña. Mass Prediction in Theropod Dinosaurs (англ.) // Historical Biology. — Taylor & Francis, 2004. — Vol. 16. — P. 85—92. — ISSN 1029-2381. — doi:10.1080/08912960412331284313.
- ↑ 1 2 3 Philip J. Currie. Allometric Growth in Tyrannosaurids (Dinosauria: Theropoda) from the Upper Cretaceous of North America and Asia (англ.) // Canadian Journal of Earth Sciences. — NRC Research Press, 2003. — Vol. 40. — P. 651—665. — ISSN 1480-3313. — doi:10.1139/e02-083. Архивировано 22 февраля 2016 года.
- ↑ 1 2 Peter Dodson, Brooks Britt, Kenneth Carpenter, Catherine A. Forster, David D. Gillette, Mark A. Norell, George Olshevsky, J. Michael Parrish, David B. Weishampel. Albertosaurus // The Age of Dinosaurs. — Lincolnwood, IL: Publications International, Ltd, 1993. — P. 106—107. — 192 p. — ISBN 0-785-30443-6.
- ↑ 1 2 3 Philip J. Currie; Jørn H. Hurum; Karol Sabath. Skull Structure and Evolution in Tyrannosaurid Phylogeny (англ.) // Acta Palaeontologica Polonica. — Институт палеобиологии — Польская академия наук, 2003. — Vol. 48, no. 2. — P. 227—234. — ISSN 0567-7920. Архивировано 26 марта 2009 года.
- ↑ 1 2 3 4 Thomas D. Carr; Thomas E. Williamson; David R. Schwimmer. A New Genus and Species of Tyrannosauroid from the Late Cretaceous (Middle Campanian) Demopolis Formation of Alabama (англ.) // Journal of Vertebrate Paleontology. — SVP, 2005. — Vol. 25. — P. 119—143. — ISSN 0272-4634. — doi:10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2. Архивировано 12 апреля 2016 года.
- ↑ 1 2 3 Philip J. Currie. Cranial Anatomy of Tyrannosaurids from the Late Cretaceous of Alberta (англ.) // Acta Palaeontologica Polonica. — Институт палеобиологии — Польская академия наук, 2003. — Vol. 48, no. 2. — P. 191—226. — ISSN 0567-7920. Архивировано 23 мая 2011 года.
- ↑ Thomas R. Holtz. Burpee Conference (англ.). Cleveland Museum of Natural History (20 сентября 2005). Дата обращения: 15 февраля 2011. Архивировано 29 января 2012 года.
- ↑ Lü J., Yi L., Brusatte S. L., Yang L., Li H. and Chen L. A new clade of Asian Late Cretaceous long-snouted tyannosaurids // Nature Communications. — 2014. — P. 1—10. — doi:10.1038/ncomms4788.
- ↑ 1 2 Henry F. Osborn. Tyrannosaurus and Other Cretaceous Carnivorous Dinosaurs (англ.) // Bulletin of the American Museum of Natural History. — New York, NY: AMNH, 1905. — Vol. 21. — P. 259—265. — ISSN 0001-0196.
- ↑ Edward D. Cope. On the Skull of the Dinosaurian Laelaps Incrassatus Cope (англ.) // Proceedings of the American Philosophical Society. — Philadelphia, PA: APS, 1892. — Vol. 30. — P. 240—245. — ISSN 0003-049X. Архивировано 20 октября 2013 года.
- ↑ Lawrence M. Lambe. On Dryptosaurus Incrassatus (Cope) from the Edmonton Series of the Northwest Territory // Contributions to Canadian Palaeontology. — Ottawa: Government Printing Bureau, 1904. — Vol. 3. — 40 p.
- ↑ 1 2 3 4 Philip J. Currie. Possible Evidence of Gregarious Behavior in Tyrannosaurids (англ.) // Gaia. — Lisbon: Universidade de Lisboa, 1998. — Vol. 15. — P. 271—277. — ISSN 0871-5424. Архивировано 11 января 2006 года.
- ↑ Lawrence M. Lambe. On a New Genus and Species of Carnivorous Dinosaur from the Belly River Formation of Alberta, with a Description of the Skull of Stephanosaurus Marginatus from the Same Horizon (англ.) // Ottawa Naturalist. — 1914. — Vol. 28. — P. 13—20. — ISSN 0008-3550.
- ↑ William A. Parks. Albertosaurus arctunguis, a new species of therapodous dinosaur from the Edmonton Formation of Alberta (англ.) // Geological Series. — University of Toronto Studies, 1928. — Vol. 21. — P. 1—42.
- ↑ Gregory S. Paul. Predatory Dinosaurs of the World. — New York, NY: Simon & Schuster, 1988. — 464 p. — ISBN 978-0671619466.
- ↑ George Olshevsky. The Origin and Evolution of the Tyrannosaurids (яп.) // Kyoryugaku Saizensen (Dino Frontline). — 1995. — Vol. 9. — P. 92—119.
- ↑ Gregory M. Erickson; Kristi Curry Rogers; David J. Varricchio; Mark Norell; Xing Xu. Growth Patterns in Brooding Dinosaurs Reveals the Timing of Sexual Maturity in Non-avian Dinosaurs and Genesis of the Avian Condition (англ.) // Biology Letters. — Лондонское королевское общество, 2007. — Vol. 3. — P. 558—561. — ISSN 1744-9561. — doi:10.1098/rsbl.2007.0254. Архивировано 14 октября 2022 года.
- ↑ 1 2 3 Andrew H. Lee; Sarah Werning. Sexual Maturity in Growing Dinosaurs Does not Fit Reptilian Growth Models (англ.) // Proceedings of the National Academy of Sciences. — NAS, 2008. — Vol. 105. — P. 582—587. — ISSN 0027-8424. — doi:10.1073/pnas.0708903105. Архивировано 16 октября 2022 года.
- ↑ 1 2 3 Brian T. Roach; Daniel T. Brinkman. A Reevaluation of Cooperative Pack Hunting and Gregariousness in Deinonychus Antirrhopus and Other Nonavian Theropod Dinosaurs (англ.) // Bulletin of the Peabody Museum of Natural History. — Музей естественной истории Пибоди, 2007. — Vol. 48. — P. 103—138. — ISSN 0079-032X. Архивировано 13 октября 2008 года.
- ↑ Robert E. Ricklefs. Tyrannosaur Ageing (англ.) // Biology Letters. — Лондонское королевское общество, 2007. — Vol. 3. — P. 214—217. — ISSN 1744-9561. — doi:10.1098/rsbl.2006.0597. Архивировано 12 апреля 2016 года.
- ↑ John R. Horner. Behavior // Encyclopedia of Dinosaurs / Philip J. Currie, Kevin Padian. — San Diego, CA: Academic Press, 1997. — P. 44—50. — 869 p. — ISBN 0-122-26810-5.
- ↑ W. Desmond Maxwell; John H. Ostrom. Taphonomy and Paleobiological Implications of Tenontosaurus-Deinonychus Associations (англ.) // Journal of Vertebrate Paleontology. — Taylor & Francis, 1995. — Vol. 15. — P. 707—712. — ISSN 0272-4634. — doi:10.1080/02724634.1995.10011256.
- ↑ Michael A. Raath. Morphological Variation in Small Theropods and Its Meaning in Systematics: Evidence from Syntarsus Rhodesiensis // Dinosaur Systematics: Approaches and Perspectives / Kenneth Carpenter, Philip J. Currie. — 1. — Cambridge, MA: Cambridge University Press, 1992. — P. 91—105. — 318 p. — ISBN 0-521-43810-1.
- ↑ Rodolfo A. Coria; Philip J. Currie. A New Carcharodontosaurid (Dinosauria, Theropoda) from the Upper Cretaceous of Argentina (англ.) // Geodiversitas. — Muséum national d'Histoire naturelle, 2006. — Vol. 28. — P. 71—118. — ISSN 1280-9659. Архивировано 26 марта 2009 года.
- ↑ Philip J. Currie; David Trexler; Eva B. Koppelhus; Kelly Wicks; Nate Murphy. An Unusual Multi-individual Tyrannosaurid Bonebed in the Two Medicine Formation (Late Cretaceous, Campanian) of Montana (USA) // The Carnivorous Dinosaurs / Kenneth Carpenter. — Bloomington, IN: Indiana University Press, 2005. — P. 313—324. — 371 p. — ISBN 0-253-34539-1.
- ↑ Walter Auffenberg. The Behavioral Ecology of the Komodo Monitor. — Gainesville, FL: University Press of Florida, 1981. — 406 p. — ISBN 0-813-00621-X.
- ↑ 1 2 David A. Eberth. Edmonton Group // Encyclopedia of Dinosaurs / Philip J. Currie, Kevin Padian. — San Diego, CA: Academic Press, 1997. — P. 199—204. — 869 p. — ISBN 0-122-26810-5.
Ссылки
- Альбертозавр в архиве Американского музея естествознания онлайн (англ.)
- Список экземпляров из базы данных по тероподам (англ.). Дата обращения: 14 февраля 2011. Архивировано 29 января 2012 года.
- Экземпляр FR 5218 (англ.). Дата обращения: 14 февраля 2011. Архивировано 29 января 2012 года.
- Экземпляр FR 5226 (англ.). Дата обращения: 14 февраля 2011. Архивировано 29 января 2012 года.
- Экземпляр FR 5234 (англ.). Дата обращения: 14 февраля 2011. Архивировано 29 января 2012 года.
- Экземпляр FR 5235 (англ.). Дата обращения: 14 февраля 2011. Архивировано 29 января 2012 года.
- Реконструкции скелетов из Буффало-Джамп (англ.). Дата обращения: 14 февраля 2011. Архивировано 8 января 2007 года.
- Раскопки останков динозавров, в том числе и альбертозавров в Нью-Мексико (англ.). Дата обращения: 14 февраля 2011. Архивировано 29 января 2012 года.
- Профиль альбертозавра на динопедии. Дата обращения: 14 февраля 2011.