Га́зовая гангре́на — омертвение тканей организма, вызванное инфекцией, обусловленной ростом и размножением в тканях клостридиальной микрофлоры. Рост этой микрофлоры возможен только при отсутствии кислорода (анаэробно), однако споры возбудителей заболевания могут длительное время сохраняться и в кислородной среде.

Золоти́стый стафилоко́кк — вид шаровидных грамположительных бактерий из рода стафилококков. Приблизительно 25—40 % населения являются постоянными носителями этой бактерии, которая может сохраняться на кожных покровах и слизистых оболочках верхних дыхательных путей.

Фосфолипаза A2 — тип фосфолипазы, гидролизующей ацильную цепь фосфолипида в sn-2 положении. При действии фосфолипазы А2 на фосфолипид образуется 2-лизофосфолипид и жирная кислота. В sn-2 положении фосфолипида могут находиться различные жирные кислоты, включая арахидоновую и эйкозапентаеновую кислоты.
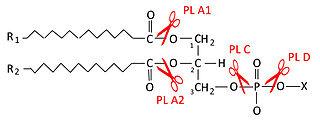
Фосфолипаза D — фосфолипаза, гидролизующая фосфодиэфирную связь между остатком фосфатидной кислоты фосфолипида и головной группой полярной части. Относится к фосфодиэстеразам также как и фосфолипаза C. Существует 2 изоформы этой фосфолипазы D1 и D2. Обе изоформы специфически расщепляют фосфатидилхолин на фосфатидную кислоту и холин, высвобождая последний в цитоплазму. Изоформы имеют различную внутриклеточную локализацию. Фосфолипаза D1 расположена в аппарате Гольджи, эндосомах, лизосомах и в секреторных гранулах, тогда как фосфолипаза D2 локализуется в липидных рафтах плазматической мембраны.

Клостри́дии — род грамположительных, облигатно анаэробных бактерий, способных продуцировать эндоспоры.
Cry-токсины — белковые токсины, продуцируемые бактерией Bacillus thuringiensis.

Clostridium perfringens (лат.) — вид грамположительных, облигатно (строго) анаэробных спорообразующих бактерий рода клостридий. Возбудитель пищевых отравлений человека, один из возбудителей газовой гангрены. Является санитарно-показательным организмом. Открыта в 1892 году Уэлчем и Нетталом.
Протеинкина́зы — подкласс ферментов киназ (фосфотрансфераз). Протеинкиназы модифицируют другие белки путём фосфорилирования остатков аминокислот, имеющих гидроксильные группы или гетероциклической аминогруппы гистидина.
Фактор активации тромбоцитов — сильный фосфолипидный медиатор воспаления. Синтезируется многими типами клеток: нейтрофилами, базофилами, тромбоцитами и эндотелиальными клетками. Участвует в воспалении, агрегации тромбоцитов, играет роль в патогенезе анафилактического шока.
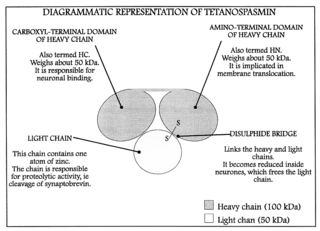
Тетанотоксин — чрезвычайно сильнодействующий нейротоксин, продуцируемый вегетативными клетками Clostridium tetani в анаэробных условиях, является причиной мышечных спазмов при столбняке. Неизвестно, какую функцию выполняет этот токсин в естественных для бактерии условиях (грунте).
Для людей смертельная одноразовая доза токсина составляет 0,2-0,3 мг.
Холерный токсин — мультисубъединичный белковый токсин, вырабатываемый холерным вибрионом. Вибрион (вирулентный штамм Vibrio cholerae[1] [2] секретирует ХТ после попадания бактерии в организм человека. Действие ХТ является причиной интенсивного обезвоживания после начала активной фазы холерной инфекции. При проникновении в клетки заражённого организма одна из субъединиц этого токсина катализирует АДФ-рибозилирование Gsα-компонента аденилатциклазы, что ведёт к её гиперактивации. Повышенная активность адениталциклазы приводит к нарушению транспорта ионов через мембрану клеток кишечника: снижается поступление ионов натрия и увеличивается отток анионов. В результате происходит быстрая потеря воды клетками кишечника, которая в некоторых случаях доходит до двух литров в час.
Лейкотриен B4 — липидный медиатор семейства эйкозаноидов из группы лейкотриенов, является медиатором воспаления.
Лакто́зный репре́ссор — ДНК-связывающий белок, который ингибирует экспрессию генов, кодирующих белки лактозного оперона. Кодируется геном lacI. Белки лактозного оперона участвуют в метаболизме лактозы в клетках бактерий. Эти гены подавляются, когда лактоза недоступна клеткам, гарантируя, что бактериальная клетка не будет тратить энергию на синтез белков, метаболизирующих лактозу, в условиях её отсутствия. Когда лактоза становится доступной, она преобразуется в аллолактозу, которая ингибирует способность лактозного репрессора связываться с ДНК. Когда репрессор не связан с оператором лактозного оперона, начинаются его транскрипция и последующий синтез ферментов метаболизма лактозы.

Систе́ма токси́н-антитокси́н — набор двух и более тесно связанных генов, которые в совокупности кодируют и белок-«яд», и соответствующее ему «противоядие». Когда такая система локализована на плазмиде, то в результате деления исходной клетки, содержащей плазмиду, дочерняя клетка выживет только в том случае, если унаследует плазмиду. Если дочерняя клетка лишена плазмиды, то нестабильный антитоксин, унаследованный с цитоплазмой матери, разрушается, а стабильный токсичный белок убивает клетку; это явление получило название «постсегрегационное убийство». Системы токсин-антитоксин широко распространены среди прокариот, и зачастую один прокариотический организм содержит множество копий таких систем.
Clostridium histolyticum (лат.) — вид грамположительных, факультативно анаэробных, спорообразующих палочковидных бактерий рода Clostridium. Является одним из возбудителей газовой гангрены у человека и млекопитающих. Её разрушающие способности, воздействующие на мягкие ткани так велики, что по истечении 12 часов после заражения, могут просматриваться кости.

АДФ-рибозили́рование — химическая реакция присоединения одного или нескольких остатков АДФ-рибозы к белку. Это обратимая посттрансляционная модификация, которая играет важную роль во многих клеточных процессах, таких как передача сигнала, репарация ДНК, регуляция экспрессии генов и апоптоз. Неправильное АДФ-рибозилирование наблюдается при некоторых формах рака. Многие бактериальные токсины, такие как холерный токсин и дифтерийный токсин, влияют на АДФ-рибозилирование.

Бактериа́льная кле́тка обычно устроена наиболее просто по сравнению с клетками других живых организмов. Бактериальные клетки часто окружает капсула, которая служит защитой от внешней среды. Для многих свободноживущих бактерий характерно наличие жгутиков для передвижения, а также ворсинок.
Бактериа́льные систе́мы секре́ции — белковые комплексы, расположенные в клеточной мембране бактерий и служащие для секреции различных белков. В частности, их используют патогенные бактерии для выделения факторов вирулентности. На основании состава, структуры и действия системы секреции делят на несколько типов. Наиболее фундаментальные различия наблюдаются между системами секреции грамположительных и грамотрицательных бактерий. Существует по меньшей мере шесть типов систем секреции, специфичных для грамотрицательных бактерий, четыре типа систем секреции уникальны для грамположительных бактерий, а два типа систем секреции имеются у обеих групп бактерий.

Вне́шняя бактериа́льная мембра́на, или нару́жная бактериа́льная мембра́на — биологическая мембрана, располагающаяся поверх слоя пептидогликана у грамотрицательных бактерий. По составу она отличается от внутренней, клеточной мембраны. На её поверхности находятся липополисахариды, являющиеся антигенами грамотрицательных патогенных бактерий.

Вези́кулы вне́шней бактериа́льной мембра́ны, или бле́ббинг-вези́кулы — липидные везикулы, которые отпочковываются от внешней мембраны грамотрицательных бактерий. С их помощью бактерии «общаются» с бактериями своего и других видов, а также окружающей средой. Эти везикулы содержат разнообразные сигнальные молекулы, в числе которых могут быть ДНК, РНК, белки, эндотоксины и другие факторы вирулентности.