Анкилозавры
| † Анкилозавры | |
|---|---|
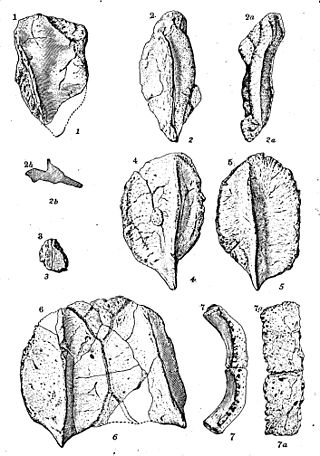
 Разнообразие анкилозавров, слева направо. | |
| Научная классификация | |
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Клада: Клада: Клада: Клада: Отряд: Подотряд: Инфраотряд: † Анкилозавры | |
| Международное научное название | |
| Ankylosauria Osborn, 1923 | |
| Распространение ископаемых остатков | |
 | |
Анкилозавры[1], или панцирные динозавры[2] (лат. Ankylosauria, от названия рода Ankylosaurus) — инфраотряд вымерших динозавров отряда птицетазовых, характерной особенностью которых являются костные образования на туловище[3].
Все представители данной группы динозавров были четвероногими. Обладали широким туловищем, короткими и массивными конечностями, сравнительно небольшим черепом. Длина тела варьировалась от 2 до 9 метров, масса от 300 кг до 6 тонн[4]. По образу жизни анкилозавров рассматривают как наземных животных, предпочитавших влажные среды обитания с богатой растительностью, такие как районы рек и озёр, прибрежные равнины. В строении анкилозавров проявляются морфологические признаки строения тела, которые в комплексе могут указывать на их тесную связь с водой[5]. Простая зубная система говорит о питании анкилозавров мягкой растительностью. Вероятно могли также питаться плодовыми телами и маленькими беспозвоночными, хотя растительность, несомненно, составляла значительную часть рациона[6].
Находки окаменелостей дают возможность предполагать, что анкилозавры произошли от четвероногих предков[7]. Самые ранние ископаемые остатки анкилозавров известны из верхней юры[8] и достигли своего пика в меловом периоде[9], расселившись по всем континентам. В России остатки обнаружены за Уралом[10]. Как и все птицетазовые динозавры, вымерли в конце мезозойской эры (мел-палеогеновое вымирание)[11].
Клада анкилозавров была впервые предложена Генри Фэйрфилдом Осборном в 1923 году для обширной группы родов, объединённых общим наличие костных пластин и шипов[12]. По степени выраженности определённых морфологических особенностей анкилозавры делятся на два основных семейства — нодозавриды и анкилозавриды[13]. Основные различия заключаются в форме и пропорциях черепа, характере костных щитков на поверхности черепа, величине зубов. В общей сложности описано около 57 видов анкилозавров, относящихся к 50 родам[14].
Благодаря своим запоминающимся внешним отличиям, анкилозавры являются частью популярной культуры всего света. Гилеозавр был одним из первых динозавров, для которого была предпринята попытка полноразмерной реконструкции в Хрустальном дворце Лондона в 1854 году[15]. Известности анкилозаврам очень поспособствовала и реконструкция рода анкилозавр в натуральную величину, фигурировавшая на Всемирной выставке 1964 года в Нью-Йорке[16].
Этимология
Название данной группы динозавров связано с рядом анатомических особенностей, в которых маленькие и большие костные щитки срослись вместе, полностью покрывая их спину и бока, на черепе эти щитки срастались с нижележащими костями, а спинные рёбра плотно прилегали к позвонкам. Латинское название Ankylosauria образовано от др.-греч. ἀγκύλος [ankylos] — «кривой», «согнутый» с анатомическим значением «жёсткий» или «слитый» и σαῦρος [sauros] — «ящер, ящерица»)[17].
В описании образца рода анкилозавр 1908 года Барнум Браун охарактеризовал семейство анкилозаврид как группу представителей, имеющих «жёсткий позвоночник», однако отметил широкую, изогнутую форму рёбер, предполагающих «сильно изогнутую» спину[18] (ошибка, основанная на предполагаемом сходстве со стегозаврами и глиптодонтами — у анкилозавров оказались довольно плоские спины). Таким образом, в дополнение к значению «жёсткая ящерица», «изогнутая ящерица» вероятно также следует читать в названии анкилозавров[17].
Общее описание

Анкилозавры — своеобразная группа динозавров, распространённых практически по всему земному шару. Большинство анкилозавров достигали длины от четырёх до шести метров[19], самые крупные представители, такие как анкилозавр, были до восьми-десяти метров в длину[20], а самые маленькие представители, такие как Struthiosaurus и Minmi, достигали всего двух-трёх метров[21]. Хвост составлял около 49—57 % общей длины от передней части шейного ряда до дистального конца хвоста, без учёта головы, и мог равняться 3 метрам[прим. 1][20]. Масса тела варьировалась от 300 кг, у небольших видов[21], до 6 тонн у самых крупных представителей[22]. Виктория Меган Арбор предположила, что максимальная масса могла достигать 8 тонн[20]. Эти животные внешне напоминали современных черепах или броненосцев. У всех анкилозавров было широкое коренастое туловище, передвигались исключительно на четырёх коротких массивных конечностях, задние конечности были длиннее передних[13]. Вследствие сильной изогнутости передних конечностей голова и передняя часть туловища были несколько более приближены к земле, задние конечности были более выпрямлены, но в целом туловище было поставлено очень низко[23][24]. Низ туловища находился на высоте от 30 до 50 сантиметров от земли[23]. Остатки передних конечностей плохо сохранились до наших дней, но по имеющимся окаменелостям установлено, что у примитивных видов анкилозавров было пять пальцев на передних конечностях, а у продвинутых — четыре. На задних конечностях у нодозаврид, по-видимому, было четыре пальца, тогда как у продвинутых анкилозаврид обычно три (у полакантид это число неизвестно)[25]. Пальцы передних и задних конечностей были толстыми и оканчивались тупыми когтями. Известен единичный случай наличия острых когтей[26]. Шея обычно достигала полуметра в длину и была довольно гибкой, учитывая толстые межпозвоночные диски[24], максимально позволяя обеспечить вертикальный охват головы на высоту до 1 метра[20]. Движения головы в горизонтальной плоскости были более свободны, чем в вертикальной, что было важно при почти боковом расположении глаз на черепе[24]. Тело анкилозавров покрывал панцирь, состоящий из сросшихся костных щитков, шипов или спинных поясов, которые являются характерной особенностью данной группы динозавров[3]. Щитки и шипы, срастаясь определённым образом, составляли различные конструкции, однако панцирь анкилозавров никогда не достигал степени консолидации черепашьего панциря, а скорее напоминал защитные образования броненосцев[19]. На хвосте некоторых видов имелся костный вырост, который мог использоваться для самозащиты[27].

Морфология
Костные пластины


Панцирь анкилозавров обычно состоит из плоских (либо имеющих гребень) костных пластин, называемых остеодермами, овальной или прямоугольной формы[28]. Размер пластин мог варьироваться от 1[29] до 35 сантиметров в диаметре[30], а толщина могла достигать 5 см[31]. Они шли вдоль спины, многочисленными продольными и поперечными рядами, и располагались в слоях кожи (отсюда и название остеодерма — «окостеневшая кожа»[28]). Остеодермы встречаются и в других группах динозавров, но пика развития они достигли у анкилозавров[32]. У некоторых видов эти остеодермы дополняются шипами на теле и хвосте, а также «булавой» на кончике хвоста[28].
Почти у всех дошедших до нас окаменелостей кожа не сохранилась, поэтому точное расположение остеодерм на теле часто неизвестно. Образец сколозавра сохранил четыре или пять поперечных рядов разделённых кожными складками, образец пинакозавра сохранил четыре продольных ряда боковых остеодерм. Исследователи предполагают, что примерно таким образом могли располагаться пластины и у других анкилозавров. Остеодермы расположенные непосредственно на спине за вторым шейным полукольцом обычно имитируют форму и расположение таковых в полукольце на шее. Между большими пластинами находились маленькие, промежуточные остеодермы, прикрывавшие зазоры[20]. Благодаря комбинированию отдельных элементов туловище анкилозавров, лишённое сплошного панциря, подобного черепашьему, оказывалось практически «бронированным» на всей поверхности[33].
У полакантид панцирь состоял как минимум из двух рядов пластин на шее, овальных килевидных пластин, расположенных чередующимися рядами вдоль спины, поперечных параллельных рядов на хвосте и атласных треугольных пластин с широким основанием или выступающих шипов по бокам шеи, тела и хвоста[34]. Панцирь нодозаврид состоял из трёх рядов больших пластинчатых остеодерм на шее, но в остальном он сильно варьируется и может включать длинные шипы в области шеи и плеч. Например у рода Edmontonia имелась пара выступающих вперёд шипов у основания шеи, тогда как у Sauropelta были два ряда шипов, самые большие из которых, в области предплечий, остриями направленные вверх. Нодозавриды также имели толстые треугольные пластины, выступающих по бокам тела и параллельные ряды килевидных овальных пластин на спине. У анкилозаврид было два ряда шейных пластин и треугольные шипы или пластины, выступающих по бокам шеи, тела и хвоста. Спина была покрыта овальными килевидными пластинами, расположенными параллельно (Euoplocephalus) или поперечно параллельными и чередующимися рядами (Saichania)[34][35][36]. Находки молодых анкилозаврид рода Pinacosaurus показывает, что нательный панцирь почти отсутствовал у молодых особей (около 1 метра в длину), ограничиваясь поясами шейного панциря. Длинные шипы у анкилозаврид отсутствуют, однако у некоторых видов на спине и плечах есть высокие тонкостенные шишки, внешне напоминающие шипы[33].
Находка рода Borealopelta, сделанная в Канаде в 2011 году, дала один из самых сохранных образцов среди анкилозавров, давая возможность разглядеть расположение шипов и остеодерм в том виде, в каком они был при жизни[37]. Эта находка, в совокупности с несколькими другими образцами, с сохранившимися костными пластинами в их естественном, прижизненном положении, позволили проводить довольно точные реконструкции внешнего вида анкилозавров[38]. Различные особенности расположения костных пластин на поверхности тела и морфология отдельных остеодерм могут быть информативными в плане видового разнообразия, в то же время могут быть значительные различия в морфологии остеодермы одной особи[3]. Гистологические исследования остеодерм анкилозавров показывают, что они различны у трёх групп семейств[39]. Остеодермы сцелидозавра и полакантид имеют утолщённый однородный внешний слой, полностью окружающий внутреннюю губчатую кость с небольшим количеством коллагеновых волокон. Остеодермы нодозаврид помимо внешнего слоя имеют достаточно сложные переплетённые коллагеновые структуры, наделяя пластины, а также шипы, очень высокой прочностью. Наконец у анкилозаврид внешний слой остеодерм достаточно тонкий, они также имеют укрепляющие коллагеновые волокна, хотя и не расположены так аккуратно, и в целом менее прочные, чем у нодозаврид[34][40].

Пластины также покрывали и боковые стороны ног. У пинакозавра и сколозавра на боковых сторонах передних конечностей имеются многочисленные яйцевидные, килевидные, а иногда и каплевидные остеодермы[20]. Маленькие круглые чешуйки на ногах заполняли пространство между крупными элементами, а также защищали живот[20][41]. Иногда те, что на задней части туловища, сливались в единый жёсткий бедренный щит, например у базальных анкилозавров Gastonia и Mymoorapelta, а также у многих нодозаврид — например Taohelong и Polacanthus. Остеодермы таза также сохранились у нескольких поздних североамериканских — сколозавр, и азиатских — пинакозавр и тархия, экземпляров. Образец сколозавра имеет смесь малых и больших кольцевых остеодерм на тазу, другой образец того же рода имеет три или четыре поперечных ряда кольцевых остеодерм, которые относительно однородны по размеру. Боковые остеодермы таза, в отличие от спинных остеодерм, имеют высокий киль и имеют треугольный профиль при виде сверху[20].
У азиатских и североамериканских видов, например Pinacosaurus, Euoplocephalus, Dyoplosaurus и Zuul, сохранившиеся хвостовые остеодермы обычно килевидные или треугольные, и расположены рядами, как и на остальной части тела. Азиатские анкилозаврины сохраняют треугольные боковые остеодермы вдоль задней части хвоста, у североамериканских образцов треугольные остеодермы хвоста пока известны только у рода Zuul[20].


У некоторых видов анкилозаврид конец хвоста дополнен большим костным диском в виде «булавы», за исключением некоторых базальных видов, например Minmi. Он состоял из остеодерм, сросшихся с хвостовыми позвонками, поддерживался и укреплялся окостеневшими сухожилиями[42]. Хвостовая «булава» появлялась только в более зрелом возрасте, когда особь вырастает более чем наполовину (около 2,5 метров)[43]. Реконструкция хвоста Euoplocephalus указывает на то, что при движении «булава» держалась чуть выше земли, но не волочилась[42]. В то время как задняя часть хвоста была жёсткой и неподвижной, подвижная, предположительно сильно мускулистая, передняя часть хвоста позволяла «булаве» раскачиваться из стороны в сторону[44]. Морфология хвостовых позвонков ограничивала вертикальные движения хвостовой «булавы»[23][42]. У относительно полных анкилозавров, неподвижная часть хвоста составляет около 55-64 % от общей длины хвоста, у рода Zuul достигает длины 210 см[20]. Наличие хвостовой «булавы» ранее было важным диагностическим признаком анкилозаврид, но ряд исследований показали, что хвостовые «булавы» могли присутствовать только у более поздних представителей, относящихся к кладе анкилозаврин и в настоящее время известны только у позднемеловых видов[45][46]. Образец анодонтозавра имеет одну из самых больших в ширину костных «булав» — 59 см[20], «булава» хвоста неопределённого анкилозавра имеет ширину 62 см[47]. Исследователи также предполагают, что был верхний предел массы «булавы», которую могли поддерживать позвонки хвоста[20]. Окаменелая «булава» анкилозаврин очень тяжёлая из-за наличия минералов, заполняющих очень пористую внутреннюю структуру. При жизни органические ткани заполняли эти пространства, делая её очень прочной[43]. «Булава» значительно различается по форме у разных родов и даже у особей одного и того же вида. При виде сверху у анкилозавра он имеет полукруглый вид, похож на таковую у родов Euoplocephalus и Scolosaurus, у анодонтозавра она имела заострённой вид, у диоплозавра была узкой и удлинённой[20].

Шея и надплечевая область, как наиболее уязвимые места, у большинства анкилозавров также были наделены своеобразной защитой[33][48]. На шее обычно было два (у анкилозаврид) или три (у нодозаврид)[46] ряда больших килевидных пластин, вместе образовывающих форму полукольца, покрывающих верхнюю часть шейного отдела и надплечевую область[45]. Пластины, составляющие плечевой пояс, значительно крупнее, чем пластины шейного пояса[33]. Нижняя частью шеи была открыта[48]. Обычно шейные полукольца состоят из нижележащей полосы неостеодермальной кости и вышележащего набора из шести больших остеодерм[20], по три на каждую сторону[48]. Иногда вокруг этих основных остеодерм присутствуют многочисленные более мелкие остеодермы. Нижняя часть шейного полукольца состоит из сравнительно толстых и плотных сегментов, квадратной или прямоугольных формы. Каждый сегмент полосы увенчан остеодермой[20][33]. У разных видов эти остеодермы различались. Рода Euoplocephalus и Anodontosaurus обычно имеют овальную форму у основания и киль размером от низкого до среднего; Scolosaurus имел круглую остеодерму с низкими коническими точками; Ziapelta имеет прямоугольные остеодермы с высокими килями. Ещё больше отличаются остеодермы рода Shamosaurus, который имеет сильно направленные вперёд вершины на дистальных остеодермах[20]. Между полукольцами, и особенно между первым полукольцом и черепом, были полосы кожи с очень мелкими ороговевшими чешуйками, часто называемые «косточками», в результате чего шея не была скована в движениях и имела относительную гибкость[48]. Промежуток между шеей и головой у анкилозаврид защищали шипы на задней части черепа, а также в области щёк. У представителей семейства нодозаврид на шее и в области плеч последние (нижние) остеодермы полуколец имели форму шипов[33][48]. Направленность вершин краевых шипов является предметом споров. На реконструкции палеосцинка[18] длинные острые краевые шипы направлены вперёд и в стороны. Таким же образом, по мнению Острома, были повёрнуты шипы плечевой области Sauropelta edwardsi[49]. Тереза Марианская считает, что у сайхании свободные шипы, располагавшиеся по бокам туловища продольными рядами, были обращены в одном ряду остриём назад, килем вперёд, а в следующем, наоборот, — остриём вперёд, килем назад[23]
Череп и зубы



Черепа анкилозавров обладали сильным окостенением и массивностью. Длина черепа взрослых особей варьируется от 20 до 60 см. Ширина в области глазниц может превышать длину[50][51]. Самый большой из когда-либо найденных черепов анкилозавров в ширину достигает 66,2 см (образец рода анкилозавр)[20], череп рода Cedarpelta имеет длину примерно 60 см[52].
Общая синапоморфия черепа анкилозавров заключается в наличии хорошо развитого черепного «орнамента», а также в присутствии только одного височного окна[3]. В отличие от других черепов динозавров, два черепных окна — предглазничное и верхнее височное — были закрыты[50]. Существовало предположение об их зарастании остеодермами[53], однако их нет и на молодом экземпляре пинакозавра[54]. Мозговая коробка почти прямая, низкая. Верхняя часть черепа, а иногда и область щёк обладали остеодермальными щитками — caputegulae[3] или «черепной орнамент». Традиционно считалось, что поверхность черепа анкилозавров имеет костную «броню», срастающуюся у взрослых с подстилающими костями крыши черепа[55][56]. Однако новые данные указывают на то, что «орнамент» частично[57] или полностью[58] является результатом ремоделирования (образование новой костной ткани) поверхности черепа. Следовательно, термин «орнамент» предпочтительнее «брони» в отношении этой текстуры. По всей видимости, орнамент соответствует вышележащим чешуйкам[57]. У нодозаврид орнамент состоит из нескольких крупных узоров, симметрично расположенных на верхней и боковой сторонах черепа, а также нижней челюсти[13][59]. Напротив, орнамент у полакантид и анкилозаврид состоял из множества мелких асимметрично расположенных чешуйчатых узоров[59]. Формы орнаментов и их узоры могут быть как таксономически информативными, так и в высокой степени похожи в пределах одного вида[20][60]. Костные пластины на черепах у некоторых видов были очень своеобразны. Например у Euoplocephalus были обнаружены окостеневшие веки[61], а у Panoplosaurus и Edmontonia были обнаружены пластины в области щёк[62].
У анкилозаврид также имелись роговые выросты: в верхней части черепа за глазницами — верхний заглазничный шип, в области щёк — нижний заглазничный шип[20][63], у нодозаврид они отсутствуют[64]. Глазницы анкилозавров средних размеров, расположенные по бокам черепа, направленные у большинства несколько вперёд, а у шамозавра — вбок[50]. Глаза в процессе эволюции передвинулись несколько назад. За счёт подобного поворота плоскости глазниц увеличивалась площадь обзора и появлялась основа для зрения с перекрытием полей обзора[6]. В конструкции черепа у многих анкилозавров имелись околоносовые пазухи, функция которых окончательно не определена. У более поздних анкилозавров они используются для формирования костного нёба[65]. Костные ноздри, как правило, крупные. Наружные ноздри у большинства анкилозавров расположены спереди, но у некоторых видов (например у анкилозавра[20]) наружные ноздри сдвинуты назад[66]. Носовая область анкилозавров была очень обширной, занимая по длине более половины черепа[67]. Общая форма черепов двух семейств заметно различается. У нодозаврид он был довольно узким и удлинённым, в то время как у анкилозаврид — шире и массивнее, что позволяет делать выводы о различных диетах данных групп[35].

Зубы анкилозавров были относительно небольшими, имели листообразный вид, сжатые с боков, с зазубренными по краю коронками[68][69]. Щёчные зубы нодозаврид обычно пропорционально больше, чем у анкилозаврид. У ранних представителей нодозаврид зубы ещё присутствовали на предчелюстной кости (передняя часть верхней челюсти), но они по-видимому были не очень функциональны и у более поздних представителей семейства остаются лишь гребни, соединяющие предчелюстную кость с началом каждого зубного ряда, а у анкилозаврид исчезающие вовсе[35][70]. Например они встречаются у примитивных Cedarpelta, Gargoyleosaurus, Pawpawsaurus, Silvisaurus и присутствуют у развитых Gastonia и Edmontonia[71]. Наличие у ранних нодозаврид предчелюстных зубов, вероятно, является архаичным признаком[70]. Зубы располагались в два ряда, смещённые внутрь, что позволяет предположить наличие щёк. Наличие щёчных пластинок, образованных внутри кожи, указывает на то, что у них была мясистая щека, вероятно, без мускулатуры[72]. Число верхнечелюстных зубов у большинства анкилозавров составляет около 20: у анкилозавра наибольшее количество верхних зубов 34-35, тогда как у Liaoningosaurus наименьшее число около 10. Количество зубов, вероятно, увеличивается во время роста, а количество взрослых зубов также варьируется между видами[46]. На зубах анкилозавров не было обнаружено явных следов стирания, подобных сильным повреждениям на коронках протоцератопсов или гадрозавров. Однако слабое стесывание зубчиков, похожее больше на заточенность края, чем на следы стирания, на некоторых зубах верхней и нижней челюсти тархии имеется[73].
Скелет

Позвоночный столб у анкилозавров состоит из 7 или 8 шейных позвонков, от 12 до 19 спинных, 3 или 4 крестцовых, 20 хвостовых у анкилозаврид и 40 или более у нодозаврид[3]. Шейные позвонки короткие, часто бывают равны по ширине и длине, спинные позвонки обычно длиннее, в результате чего область шеи небольшая, а туловище достаточно длинное. Кроме того, рёбра, в местах крепления к позвонкам, направлены вверх под углом между 30 и 50 градусами. Этот высокий угол заставляет рёбра выгибаться наружу, образуя широкое бочкообразное тело. Последние четыре (иногда больше) ребра часто неподвижно срастаются с позвонками. Это слияние, по-видимому, связано с чрезвычайно широким тазом[74]. Подвздошная кость повёрнута в сторону и создаёт почти горизонтальную структуру, предположительно, чтобы освободить больше места для объёмной пищеварительной системы. Чтобы обеспечить большую стабильность этого таза, задние три-шесть спинных позвонка и один-три передних хвостовых позвонка сливаются с крестцом, образуя сложно устроенный крестец — synsacrum. Эти сросшиеся позвонки предотвращают вращение таза вокруг позвоночника[13][55]. Хвостовые кости обычно короткие около таза, и первые один или два могут фактически срастаться с крестцом. Хвостовые кости быстро удлиняются, за исключением области около таза, где они намного больше в ширину, чем в длину. У анкилозаврид дистальная треть хвоста может сливаться, обеспечивая жёсткую основу для терминальной «булавы» хвоста; у нодозаврид и полакантид хвостовые кости остаются длинными до тех пор, пока не заканчивается хвост, где последние три (или около того) кости быстро укорачиваются[75]. В целом у нодозаврид был более длинный хвост, чем у анкилозаврид[76].
Кости конечностей довольно крепкие, чтобы выдерживать вес динозавра. Большая часть веса тела приходилась на задние лапы; следовательно, бедро прямое и столбовидное, а большеберцовая и малоберцовая кости короткие и толстые. Бедренная кость у нодозаврид и полакантид более тонкая, чем у анкилозаврид[13]. Нижняя часть передних конечностей состоит из большой крепкой локтевой кости и меньшей лучевой кости[25]. Плечевая кость анкилозавров имеет хорошо развитый дельтовидный гребень, служивший местом крепления нескольких основных мышц, используемых при передвижении[13]. Плечевая кость пропорционально более тонкая у нодозаврид, чем у анкилозаврид[77]. Особенности строения плечевых костей говорят о том, что предплечья держались не прямыми, как у современных слонов, а были согнуты. В области надплечья бросается в глаза удлинённая лопатка, которая — особенно у нодозаврид — имела выпуклый вырост (акромион), он служит местом крепления передней плечевой мышцы. Олекранон локтевой кости у анкилозавров хорошо развит и обеспечивает высокий рычаг для мышц-разгибателей локтя[25]. Таз необычен для птицетазовых динозавров, потому что подвздошная кость расширилась по горизонтали, образуя широкую поверхность, охватывая большую кишку. Кроме того, вертлужная впадина или тазобедренная впадина не открытая, как у остальных динозавров, а имеет чашеобразный вид; лобковая кость, расположенная ниже подвздошной кости, небольшая или может вообще отсутствовать. Напротив, седалищная кость остаётся выступающей и выступает вниз, что более типично для ящеротазовых динозавров. Седалищная кость у анкилозаврид довольно прямая, в то время как у нодозаврид и полакантид она изогнута или изогнута в середине[25].
Происхождение и эволюция

Происхождение анкилозавров до сих пор плохо изучено, поскольку известно лишь несколько образцов из средней юры, когда по-видимому и появились первые анкилозавры[60]. Предков анкилозавров долгое время искали среди стегозавров, это наиболее близкая к анкилозаврам группа по сравнению с другими динозаврами. В настоящее время анкилозавры являются сестринской группой стегозаврам внутри клады Eurypoda[78]. Их объединяет наличие остеодермальных элементов в коже, узкий треугольный череп стегозавров сходен с таковым нодозаврид, некоторое сходство обнаруживается и в строении нёба[79]. Поскольку стегозавры известны из средней юры[80], анкилозавры должны быть как минимум того же возраста. Возможно они разделились во время аалена, более 170 миллионов лет назад. Однако хорошо сохранившиеся остатки анкилозавров того возраста отсутствуют. Известны лишь неполные лучевая и локтевая кости с острова Скай в Шотландии байоского возраста, точная принадлежность которых к анкилозаврам или стегозаврам не установлена[81]. Скорее всего анкилозавры пошли по иному эволюционному пути, чем стегозавры. У последних остеодермы становятся поднятыми, а боковая защита исчезает[13]. Анкилозавры же эволюционировали в сторону развития остеодермального слоя верха черепа, к усилению «бронированности» и дальнейшей консолидации панциря[23], что предполагает наличие у предка панциря, состоявшего из отдельных не сраставшихся друг с другом остеодермальных элементов[82].

Также нет возможности точно определить, откуда впервые произошли анкилозавры. На сегодняшний день самыми ближайшими предками анкилозавров считаются сцелидозавр из нижней юры Англии и Emausaurus из нижней юры Германии и дают возможность предполагать, что анкилозавры впервые появились в Европе в раннем юрском периоде и произошли от некоторых примитивных тиреофор[83][84]. Тиреофоры, это группа «бронированных» растительноядных динозавров, чьи несколько более поздних эволюционных ветвей вероятно перешли с двуногого на четвероногий способ передвижения. Гипотетических предков анкилозавров вероятнее всего предполагать четвероногоходящими[79]. Тереза Марианская также предполагает, что предки анкилозавров уже были, по всей вероятности, четвероногоходящими в отличие от орнитопод и цератопсов, исходная бипедальность которых очевидна[23]. Будучи полностью наземными животными, анкилозавры совершали межконтинентальные миграции во времена, когда континенты были соединены между собой[85]. В ранней юре все континенты ещё более или менее объединялись в суперконтинент Пангея. Таким образом анкилозавры, зародившись в Европе, легко могли распространиться по всему земному шару. В этом плане примечательны находки анкилозавра Австралии — Minmi. Согласно анализу Виктории Меган Арбор данный род является представителем основной и самой старой ветви эволюционного древа среди известных анкилозавров. Однако Minmi относится всего лишь к меловому периоду и не является самым ранним из известных анкилозавров, что даёт возможность предполагать о существовании более старой и обширной ветви эволюционного древа, которая претерпела свой собственный процесс эволюции в Гондване. Однако имеющихся данных недостаточно для более точной проверки этой гипотезы[60].
Ранние анкилозавры
Считается, что анкилозавры существовали на планете в течение 100 миллионов лет и были довольно успешны с точки зрения эволюции[16], пока не вымерли в конце мелового периода 65 миллионов лет назад[11]. Юрские анкилозавры были в основном относительно небольшие по размеру, длина тела не превышает 4 метров. Например Mymoorapelta и Gargoyleosaurus в Северной Америке, Tianchisaurus в Азии, и Cryptodraco и Dracopelta в Европе были около 3 метров в длину[46][86]. Их вес мог колебаться от трёхсот килограммов до двух тонн.
Самые ранние из известных анкилозавров средней юры представлены фрагментарными остатками. Они мало что дают об их родстве с более поздними анкилозаврами. Их фрагменты известны из келловея и оксфорда Англии, между 165 и 160 миллионами лет. Большинство из них настолько фрагментарны (состоят из отдельных костей или зубов), что их невозможно отличить от других анкилозавров, и они считаются недействительными видами — nomen dubium. Например Sarcolestes описан по части нижней челюсти, Priodontognathus по части верхней челюсти, Cryptodraco по бедренной кости[35][87]. Ещё более древним анкилозавром, относящемуся к байосу—бату, возрастом 168 миллионов лет, может быть скелет Tianchisaurus, остатки которого более полные. По мнению Донг Чжимина, описавшего данный род, Tianchisaurus может являться самым ранним анкилозавром[8]. Однако род плохо описан, датирован ненадёжно и отсутствует большая часть скелета для более детального описания[60]. Основная гипотеза распространения анкилозавров предполагает, что они впервые появились в Европе в период нижней юры[88]. Самый ранний хорошо сохранившийся анкилозавр европейского континента относится к роду Dracopelta. Примитивные анкилозавры распространились из Европы в Северную Америку, куда они попали в конце юры. Наиболее полные остатки из ранних анкилозавров, которые были обнаружены в Северной Америки, принадлежат родам Mymoorapelta и Gargoyleosaurus, жившие более 155—150 миллионов лет назад. Они относятся к семейству нодозаврид[89]. В настоящее время нет никаких известных записей о скелетных остатках анкилозавров из юрского периода Гондваны[90].
В юрский период анкилозавры, по-видимому, представляли лишь небольшую часть фауны. Конкуренцию им составляли стегозавры, которые вместе с анкилозаврами были основными поедателями низкорослых растений[91]. Однако во время перехода от юры к мелу флора изменилась. Стегозавры постепенно уменьшаются в численности вместе с их пищевыми растениями. Наоборот анкилозавры, которые обладали более широкими челюстями и огромной брюшной полостью, смогли диверсифицироваться и стать более многочисленными за счёт потребления большего количества некачественной пищи[92][93].
Расцвет и вымирание в меловом периоде
Наибольшая эволюция анкилозавров произошла в меловом периоде (142—65 млн лет назад), особенно в северном полушарии[94]. В качестве окаменелостей анкилозавры наиболее многочисленны в слоях от баррема до альба (примерно 127—98,2 млн лет) и в слоях от кампана до маастрихта (примерно 83,5—65 млн лет)[95]. Эти интервалы также отмечают и наибольшее видовое разнообразие. Образцы из верхнего мела включают несколько исключительно хорошо сохранившихся скелетов, при этом большая часть костных пластин сохранилась в исходном положении[23][76]. Они становятся важными фаунистическими компонентами в лавразийских экосистемах, но оставались редкими в Гондване[3]. В меловом периоде их размеры увеличиваются, большинство меловых анкилозавров имеют длину тела более 5 метров[46]. Например, примитивный анкилозаврид Cedarpelta имел предполагаемую длину тела 7,5—8,5 метров[58]. Sauropelta из Северной Америки является одним из самых крупных нодозаврид раннего мела, его длина составляет 5,5 метров, такие крупные нодозавриды неизвестны в меловых отложениях Европейского архипелага, из-за чего Переда-Субербиола назвал европейских нодозаврид «карликовыми островными формами»[96].
Ранний мел был особенно важен для эволюции анкилозавров, поскольку в летописи окаменелостей впервые появились два семейства анкилозавров — нодозавриды и анкилозавриды[35]. Расхождение нодозаврид и анкилозаврид произошло предположительно в юре, поскольку раннемеловые представители этих ветвей имеют уже типичное для своего семейства строение[82]. Их синапоморфии указывают на гораздо более длительную эволюционную историю с призрачными линиями, возможно, восходящими к средней юре[35]. Поскольку у нодозаврид наблюдаются черты, которые с большим основанием можно считать исходными, то, по-видимому, общего предка анкилозавров можно назвать нодозавроподобным. Существенные различия в морфологии анкилозаврид вызваны, возможно, их ранним ответвлением от предкового ствола и приспособлением к несколько иным условиям обитания и образу жизни[97]. Предполагается что нодозавриды впервые появились в Европе. Питер Гальтон сделал вывод о европейском происхождении нодозаврид из-за экземпляров средней юры (Hylaeosaurus), относящихся к этому семейству[88]. К началу мелового периода североамериканские нодозавриды уже хорошо известны — Sauropelta. У данного рода все признаки нодозаврид уже хорошо развиты, что указывает на существование ещё не найденных более примитивные форм[98]. Для многих раннемеловых и среднемеловых формаций Северной Америки нодозавриды составляют большую долю фауны птицетазовых динозавров[99]. Есть также убедительные доказательства присутствия нодозаврид в раннем мелу Азии. В середине мелового периода азиатские нодозавриды были заменены анкилозавридами[60]. Эта смена могла быть связана с достаточно крупной перестройкой фаунистических комплексов суши и моря, климатической картины, отмечаемых в этот период[100], либо лучшей приспособленностью анкилозаврид к давлению со стороны хищников[101].
Первые анкилозавриды известны по плохо сохранившимся остаткам раннего мела, около 145 миллионов лет назад. Самый ранний известный анкилозаврид — это Liaoningosaurus из нижнего мела (баррем) провинции Ляонин в Китае[102]. Предполагается, что именно в Азии происходило раннее разветвление родственных групп и появление новых видов анкилозаврид. В середине мелового периода (около 100 млн лет назад) геологические блоки, составляющие географический регион современной Аляски, достигли своего современного местоположения. Примерно в это время образовалось вероятное сухопутное соединение между Азией и Северной Америкой и было вероятным маршрутом расселения динозавров с одного континента на другой[103]. Между альбом и сеноманом примитивные анкилозавриды мигрировали в Северную Америку из Азии, где они превратились в кладу более развитых форм — анкилозаврин. У них были ещё более широкие головы, чем у ранних анкилозавров, и резко изогнутая морда[60]. В этот же период анкилозаврины приобрели свою знаменитую хвостовую булаву, толстое окостенение на конце хвоста, которое могло использоваться для отражения нападений хищных динозавров[104]. Самым раннем представителем, обладавшим булавой на хвосте является род Jinyunpelta, обитавший сто миллионов лет назад[47].

Небольшое временное перекрытие между ранними нодозавридами и группой примитивных анкилозавров (полакантиды)[105] на североамериканском континенте в формации Кедровая гора (англ. Cedar Mountain Formation; нижний мел), повышает вероятность конкурентного вымирания группы последних к концу баррема—апта[35]. К альбу они заменили семейство полакантид и в Европе. Представители нодозаврид становятся доминирующими в летописи североамериканских окаменелостей анкилозавров в апте—альбе (121—98,5 млн лет) и в сеномане—сантоне (98,5—83,5 млн лет) и сохраняют своё присутствие в Ларамидии на протяжении всего позднего мелового периода[35]. Напротив, мигрировавшие из Азии анкилозавриды подверглись локальному вымиранию одновременно с затоплением Западного внутреннего моря в сеномане и не показывают своего доминирования вплоть до кампана[106]. Многие находки нодозаврид происходят из морских отложений, анкилозавриды напротив в значительно меньшей степени представлены в морских отложениях, что и даёт возможность исследователям предположить о преобладании группы нодозаврид, поскольку не морские пласты были редки для среднего мела. Возможно, между этими двумя группами существовало какое-то нишевое разделение, в котором нодозавриды ели более качественную пищу[75].
Тем не менее, это преобладание изменилось в течение позднего мела (кампан—маастрихт; 83,5—65 млн лет назад), когда анкилозавриды увеличились в разнообразии и относительной численности. Особенно разнообразны они становятся в Северной Америке, а также в Азии. Североамериканский континент становится местом обитания как типичные местных видов анкилозавров, так и вновь мигрировавших азиатских форм[107]. К этому времени нодозавриды появляются в Антарктиде (кампан; 83—71 млн лет назад)[108], вероятно, через Южную Америку, где также известны нодозавриды позднего мела[109]. Что помешало миграции анкилозаврид в Южную Америку неизвестно[60]. Азиатские и североамериканские анкилозавриды известны как из засушливых (эоловых), так и из низинных прибрежных районов[75]. Некоторые виды анкилозаврид начинают достигать значительных размеров: самые большие ископаемые фрагменты (род анкилозавр) указывают на длину девять метров и вес в 6 тонн. В этот период в Северной Америки наблюдается общая тенденция увеличения размеров у многих групп динозавров — тираннозаврид, цератопсид, анкилозаврид и других. Это могло быть результатом эволюционной конкуренции «хищник—жертва» или расширения пищевой базы по мере того, как отступающее Западное внутреннее море соединяло восточную и западную части континента в большую сухопутную область, или же их комбинацией. Увеличение размеров анкилозаврид могло являться адаптацией для защиты от столь же крупных хищных динозавров. В то же время увеличение размеров у нодозаврид в это время не отмечается[110].
Вымирание анкилозавров в Северной Америке и Европе, по-видимому, произошло примерно в одно и то же время в конце маастрихта (примерно 67—66 млн лет назад), но задолго до конца маастрихта (65 млн лет назад) и исчезновения динозавров. Отсутствие анкилозавров в верхних слоях маастрихта считается подтверждённым фактом в результате тысячи человеко-часов, потраченных разными людьми на поиски их окаменелостей начиная с XIX века. Кеннет Карпентер и Брент Брейтхаупт задокументировали полное отсутствие остатков анкилозавров в более верхних слоях формаций Ланс и Хелл-Крик[11]. Вымирание как в Северной Америке, так и в Европе могло быть связано с потерей прибрежных местообитаний в результате регресса многих мелководных континентальных морей, произошедшего в то время по всему земному шару[111][112]. Полное вымирание анкилозавров в конце мелового периода не было внезапным. В течение последних трёх миллионов лет до мел-палеогенового вымирания происходит снижение их разнообразия в Северной Америке[11], предполагая, что по крайней мере в Северной Америке они вымерли до удара астероида[113]. Что касается азиатских анкилозавров, конец мелового вымирания всех динозавров слишком плохо задокументирован, чтобы знать точно, когда произошло окончательное вымирание анкилозавров[114].
История находок

Окаменелости анкилозавров — одни из самых ранних находок динозавров. Скелет Hylaeosaurus был обнаружен летом 1832 года на каменоломнях в Англии и состоял из относительно хорошо сохранившихся костей передней части туловища[115]. Находка поразила учёных необычным костным панцирем и многочисленными шипами. В 1842 году Ричард Оуэн включил его в число трёх родов, наряду с мегалозавром и игуанодонтом, на основе которых описал новую «трибу или подотряд» — динозавры[116]. Однако после этого многообещающего начала, описания новых видов анкилозавров приостановились. Странные особенности Hylaeosaurus, такие как плотный панцирь и длинные шипы, рассматривались как особенности только данного вида, не типичные для обширной коллекции родственных форм. Следующими важными находками стали скелет Polacanthus, описанный в 1865 году[117][118] и несколько фрагментарных остатков Acanthopholis, описанных в 1867 году[119]. Из-за ограниченного количества ископаемого материал и времени их описания, когда отсутствовали современные методы кладистики, данный род был отнесён к гораздо более известному сцелидозавру, описанному в 1861 году[120]. В какой-то степени связь этих динозавров была определена правильно, так как сцелидозавр вместе с анкилозаврами принадлежит к более широкой группе, которая ныне называется тиреофорами[121]. Начиная с 1870 года знания о динозаврах в целом были значительно улучшены благодаря обширным открытиям в Европе и Северной Америке[122]. Были описаны и несколько новых родов анкилозавров: в 1871 году Эмануэль Бунцель опубликовал научный труд, в котором описал род Struthiosaurus[123], а в 1879 году Гарри Говир Сили описал молодую особь Anoplosaurus[124]. Большинство находок динозавров данного периода в основном относились к юрским отложениям, в которых остатки анкилозавров довольно редки даже сейчас[60]. Тем не менее по фрагментам челюстей и бедренной кости в 1875 году были описаны рода Priodontognathus[125] и Cryptosaurus[126], в 1893 году Sarcolestes[127], однако точная их принадлежность к анкилозаврам была доказана только после девятнадцатого века.
Ранний период изучения панцирных динозавров отличался обсуждением вопроса самостоятельности группы, её статуса. В 1890 году профессор Отниэль Чарльз Марш после описания нового рода динозавров — нодозавра, определил и новую кладу — нодозавриды. Но в те времена ещё не было понимания того, что эта группа является одним из двух основным семейств панцирных динозавров; находка была слишком фрагментарной, Марш даже думал, что нодозавриды были частью группы Ceratopia[128]. Ко времени описания Stereocephalus[129] в 1902 году из формации Олдмэн Северной Америки анкилозавры были разбросаны в трёх семействах подотряда Stegosauria, будучи объединёнными в него со стегозаврами по наличию остеодермальных образований[130][131].

В начале 20-го века как в США, так и в Канаде были сделаны ряд крупных находок из верхнего мела. Они включали в себя как двух анкилозаврид, такие как анкилозавр и Euoplocephalus, так и остатки нодозаврида — эдмонтония. Только теперь открытия новых видов давали лучшую картину конструкции анкилозавров. Однако пробелы в знаниях оставались, потому что помимо более полных скелетов, было найдено много разрозненных черепов и костных пластин. Вместе с тем постепенно в пределах подотряда стегозавров обосабливались группы родов — будущие анкилозавровые семейства[68]. В 1908 году Барнум Браун на основе описания нового рода Ankylosaurus, установленного по черепу и неполному посткраниальному скелету, определил новую группу — анкилозавриды[18]. Таким образом, среди стегозавров обособилось ещё одно семейство. Затем на протяжении нескольких десятилетий существенных изменений в систематике анкилозавров в целом и анкилозаврид в частности не предлагалось. Наличие наиболее часто используемых сегодня названий семейств в классификации, ни в коей мере не означало, что существующие в те времена определения были общепринятыми. До начала 20-х годов XX века вокруг панцирных динозавров продолжалась большая путаница. Анкилозавры чаще всего оказывались объединёнными со стегозаврами в разного ранга группы в пределах последних[68], хотя основные идеи эволюционных линий были более-менее правильными. В 1923 году Генри Фэрфилд Осборн впервые использовал название анкилозавры для более-менее уже обширной группы панцирных динозавров[12]. В этом же году Франц Нопча предложил схему, преимуществом которой было уравнение рангов трёх групп четвероногих орнитисхий: стегозавров, анкилозавров и цератопсов, объединение их в подотряд тиреофор вызвало понижение на ранг этих групп[132]. Альфред Ромер в 1927 году принял и использовал идеи об изолированности анкилозавров в пределах орнитисхий и об обособленности от стегозавров. В его работе были подчёркнуты различия в строении таза, не имеющего подобий среди других орнитисхий[133]. Дальнейшие работы были направлены в основном на распределение родов среди анкилозавров[134][135]. В 1956 году Альфред Ромер разделил подотряд на две группы — Acanthopholidae и нодозавриды. В первую группу он включил примитивных представителей, в последнюю — более развитые формы. В этой работе Ромер сразу отметил временный характер предложенной им схемы из-за недостаточной изученности группы[136]. Ранг подотряда сохранён за группой анкилозавров и в работе Евгения Малеева «Основы палеонтологии»[1].
К 70-м годам XX века находки этих животных были представлены, как правило, щитками и шипами панциря или фрагментами посткраниального скелета, черепной материал отличался обычно плохой сохранностью. В силу этого изученность группы оставалась достаточно низкой, а описания носили в основном поверхностный характер. В это время появились монографии и отдельные статьи, в которых значительно детальнее рассматривается морфология азиатских и североамериканских панцирных динозавров. Из-за слабой изученности морфологии анкилозавров открытыми оставались вопросы об их происхождении, связях с родственными группами и о взаимоотношениях внутри группы[68]. В 1977 году польский палеонтолог Тереза Марианская предположила, что анкилозавры не были частью птицетазовых динозавров, и развились как отдельная ветвь этозавров[23]. Эта группа животных также развила в процессе конвергентной эволюции защищённую часть тела, внешне напоминающий панцирь анкилозавров, однако в более поздних своих работах Марианская всё же признала связь анкилозавров и птицетазовых динозавров[137].
В 1978 году Уолтер Престон Кумбс опубликовал статью «Семейства птицетазовых динозавров отряда анкилозавров»[13], в которой он обобщил результаты своей диссертации, которую защищал в 1971 году[138]. Кумбс сначала попытался дать анкилозаврам чёткое место в генеалогическом древе, как представителям птицетазовых динозавров, и ещё больше упростить их внутреннее разделение на анкилозаврид и нодозаврид. Он попытался разместить все известные на тот момент виды по этим двум группам. Более того, чтобы увеличить точность порядка, он скрупулёзно удалил каждый вид, который мог обозначать более молодой синоним другого вида[13]. Хотя его публикация по-прежнему считается важной вехой в исследованиях анкилозавров, она уже достаточно устарела. В конце 90-х годов и в начале 2000-х были описаны ещё несколько новых родов анкилозавров, такие как Gastonia, Gargoyleosaurus и Mymoorapelta, которые обладали сочетанием свойств двух семейств, стирая «традиционные» различия между нодозавридами и анкилозавридами[105]. Даже сегодня изучение анкилозавров пока не дало стабильных результатов. Первой причиной этого является появление в 1970-х годах метода кладистики, который подходит к вопросу об эволюционном развитии именно путём определения индивидуальных характеристик всех обнаруженных анкилозавров и последующего вычисления с помощью компьютерной программы генеалогических деревьев[105]. Стали появляться попытки кладистических анализов группы. Пол Серено[55], а также Уолтер Кумбс совместно с Терезой Марианской[139] представили кладограммы анкилозавров (первый включил в анализ только два рода и две составных группы — Panoplosauridae и Ankylosaurinae, вторые использовали в анализе только 14 родов), которые поддержали традиционную раздвоенность группы на нодозаврид—анкилозаврид и предоставили списки синапоморфий в поддержку их родства. Однако ни одно из этих исследований не было основано на численном анализе. Аналогичным образом Татьяна Туманова представила филогенетическое древо анкилозаврид[63], основанное на наличии общих характеристик, но более детального анализа не проводилось[105]. Первый матричный кладистический анализ анкилозавров провёл Ю. Ли в 1996 году, однако и он включил только 26 признаков (описывающие только черепные вариации) и восемь видов[105]. В кладистические анализы М. Викариуса[140] и Р. Хилла[141] также были включены только черепные признаки, что привело к исключению множества действительных видов, известных по посткраниальным скелетами, у которых отсутствуют черепа. Данные анализы, которые были не всегда полны, делали результаты кладистических исследований нестабильными. Всё это подкреплялось второй причиной — большое количество находок с описанием новых родов и видов.

Ещё в 1920-х годах американские экспедиции Ч. Берки и Ф. Морриса[142] в пустыню Гоби между Монголией и Китаем обнаружили череп панцирного динозавра, описанного Чарльзом Гилмором как Pinacosaurus grangeri в 1933 году[143]. Новые советско-монгольские, советско-китайские, польско-монгольские (1965—1971), канадско-китайские (1987—1990), японско-монгольские (1993—1998) и корейские палеонтологические экспедиции вызвали лавину находок новых ископаемых материалов со времён Второй мировой войны, часто прекрасно сохранившихся в песчаных отложениях пустыни[141]. Обнаруженных скелетов было слишком много, чтобы описать даже незначительную их часть, а многие так и остались в местах находок. Был открыт первый полный скелет анкилозавра — Pinacosaurus. Азиатские находки также позволили описать множество новых видов[141], различия между которыми было нелегко указать из-за сбивающего с толку разнообразия форм. Уолтер Кумбс предположил, что костные пластины анкилозавров разнятся до такой степени, что их трудно использовать для определения родовых характеристик[13]. Однако до сих пор существует множество комбинаций черт, которые позволяют всем окаменелостям определённого места или геологического образования быть обобщёнными. На основании этих черт Виктория Меган Арбор смогла объявить многие виды, которые Уолтер Кумбс объединил как синонимы, действительными (признанными в научной среде). Тот же метод она использовала для классификации азиатских находок[60]. Большинство из 50 известных на сегодня видов, были обнаружены после 1978 года[10].
География находок
В настоящее время можно говорить уже о повсеместном распространении анкилозавров. Наибольшее количество находок приходится на Северную Америку, Азию и, несколько в меньшей степени, на Европу[3][144]. Хорошо сохранившиеся ископаемые остатки известны в отложениях верхней юры и мела на территории современных Европы, Северной Америки и Центральной Азии, мела Австралии, Антарктиды[3](Antarctopelta был первым динозавром, обнаруженным на этом континенте[108]) и Южной Америки[145], известны фрагментарные остатки из Африки[146] и Новой Зеландии[147]. Есть некоторые свидетельства, что анкилозавры могли также обитать на территории нынешнего Мадагаскара[148].
Европа
Европа дала немалое количество ископаемых остатков анкилозавров, уступая только Северной Америке и Азии[144]. Тем не менее остатки этих четвероногих животных на европейском континенте не так часты и сравнительно мало изучены. Большинство остатков представлены в основном посткраниальными (часть скелета, за исключением черепа) костями, с отсутствием сопутствующего черепного материала. Череп или кости нижней челюсти, будучи наиболее важными элементами для уточнения родового различия и их систематики, во многих случаях являются либо изолированными образцами, либо фрагментарными остатками, что делает сравнение образцов находок весьма сомнительным[149]. В то время как западноевропейская летопись ископаемых характеризуется широким временным диапазоном (около 90 млн лет, с некоторыми временными разрывами), остатки анкилозавров к востоку от реки Рейн известны почти исключительно от сантона до маастрихта — временной промежуток около 20 млн лет. Кроме того у более поздних представителей европейского континента (особенно в его восточной части) идёт смещение в сторону эндемических характеристик (присущих только данной местности) — небольшие размеры и причудливые, своеобразные формы обитавшие на островах позднемеловой Европы[149].
Остатки анкилозавров из юрских отложений Европы чрезвычайно редки, и большинство из них представляют собой изолированные, фрагментарные кости. Самые ранние находки существования анкилозавров встречаются начиная со средней юры (келловей и оксфорд)[88][150] — Sarcolestes, Cryptodraco, Priodontognathus. Более поздний примитивный европейский анкилозавр — Dracopelta, относится к верхней юре (титон). В то время как Кеннет Карпентер[35] рассматривал всех их в качестве nomina dubia, М. Викариус[3] относил часть из них к действующим родам без ясной позиции внутри группы анкилозавров — incertae sedis. Помимо этих позднеюрских родов анкилозавров, некоторые остеодермы из киммериджа и хвостовой позвонок из титона Англии были отнесены к Ankylosauria indet[149].
Раннемеловая (берриас—апт) летопись по сравнению с юрской более многочисленна и разнообразна, включает лучший черепной материал и несколько частичных скелетов. С другой стороны, черепные остатки, определённо принадлежащие к этим видам, мало изучены. Нижнемеловая группа анкилозавров известна из Британии и представлена тремя действительными (признанными в научной среде) в настоящее время видами: Hylaeosaurus armatus (формация Гринстед-Клэй, Западный Суссекс), Polacanthus foxii (формация Уэссекс, остров Уайт) и Horshamosaurus rudgwickensis (группа Вельд-Клэй, Западный Суссекс). Также известны виды Anoplosaurus curtonotus, Europelta carbonensis[149].
Позднемеловая летопись анкилозавров не представлена каким-либо ископаемым материалов анкилозавров на континенте вплоть до сантона. Единственный материал анкилозавра из первой половины позднего мела был отнесён к различным видам Acanthopholis[149]. Переда-Субербиола и Баррет пришли к выводу, что все пять известных видов Acanthopholis являются nomina dubia, поскольку материал фрагментарен, недиагностичен и в некоторых случаях это возможно совокупность различных видов/особей[151]. Сантонская и маастрихтская летопись анкилозавров более полна, включая четыре различных вида, относящихся к двум родам. Материал не ограничивается Западной Европой и относительно распространён также в Центральной и Восточной Европе. Hungarosaurus tormai является самым изученным европейским анкилозавром позднемелового периода, описанный на основе многочисленных частичных скелетов из Западной Венгрии. Struthiosaurus — самый распространённый род европейских анкилозавров, известный по находкам от Испании до востока бассейна Хацега, Румыния. Остатки этого рода известны от сантонских до маастрихтских отложений, имеющие одно из самых длинных временных распределений (около 17 млн лет) среди родов анкилозавров[149].
| Известные рода и виды | |||||
|---|---|---|---|---|---|
| Род | Виды | Семейство | Возраст | Место находки | |
| Acanthopholis Huxley, 1867 | Acanthopholis horrida | Нодозавриды | Сеноман | Англия | |
| Anoplosaurus Seeley, 1879 | Anoplosaurus curtonotus | Нодозавриды | Альб | Англия | |
| Cryptodraco Seeley, 1869 | Cryptodraco eumerus | incertae sedis | Оксфорд | Англия | |
| Dracopelta Galton 1980 | Dracopelta zbyszewskii | incertae sedis | Киммеридж | Португалия | |
| Europelta Kirkland et al., 2013 | Europelta carbonensis | Нодозавриды | Альб | Испания | |
| Hungarosaurus Ősi, 2005 | Hungarosaurus tormai | Нодозавриды | Сантон | Венгрия | |
| Horshamosaurus Blows, 2015 | Horshamosaurus rudgwickensis | Нодозавриды | Баррем | Англия | |
| Hylaeosaurus Mantell, 1833 | Hylaeosaurus armatus | Нодозавриды | Валанжин | Англия, Германия | |
| Polacanthus Owen, 1865 | Polacanthus foxii | Нодозавриды | Баррем—Апт | Англия, Испания | |
| Priodontognathus Seeley, 1875 | Priodontognathus phillipsii | incertae sedis | Оксфорд | Англия | |
| Sarcolestes Lydekker, 1893 | Sarcolestes leedsi | incertae sedis | Келловей | Англия | |
| Struthiosaurus Bunzel, 1871 | Struthiosaurus austriacus Struthiosaurus languedocensis Struthiosaurus transylvanicus | Нодозавриды | Сантон—Кампан | Австрия, Венгрия, Испания, Румыния, Франция | |
Азия
Наиболее хорошо сохранившиеся находки окаменелостей анкилозавров относятся к Азии, где многие образцы представлены почти полными и сочленёнными скелетами[152]. Ископаемые остатки азиатских анкилозавров достаточно многочисленны. Основной описанный материал собран в аптских—маастрихтских отложениях на территории Монголии и Китая. Отдельные, большей частью разрозненные остатки найдены и на территории стран СНГ[153]. О находках азиатских анкилозавров имеются сообщения из Индии[154], возможный образец анкилозавра был обнаружен в сеноманских морских отложениях северной Японии (частичный череп и несколько зубов с остеодермами)[155].
О юрских анкилозаврах Азии практически ничего не известно. Из отложений данного периода найдены только два свидетельства существования анкилозавров. Самым ранним считается Tianchisaurus из формации Тутунхэ (бат—оксфорд)[8]. Вторая находка относится к позднеюрской формации Цигу в южной части Джунгарской впадины. Находки в некой степени дают возможность заполнить пробел в знаниях ранней эволюции панцирных динозавров Азии[156]. Ещё одна находка, сделанная в Индии, сомнительна и вызывает споры[90][154].
Остатки анкилозавров в нижнемеловых отложениях (апт—альб) Монголии также сравнительно редки, впервые они были найдены на местонахождении Хобур[157] в 1974 году. В прослоях песчаников, песков и алевролитов были найдены кости Psittacosaurus mongoliensis. Ещё одна находка нижнемелового анкилозавра Shamosaurus была сделана в 1977 году на местонахождении Хамрын-Ус[158]. Фауна нижнемеловых отложений является сходной с имеющейся на местонахождении Хобур, что свидетельствует об идентичности возрастов обоих местонахождений. Захоронение анкилозавров происходило в условиях, соответствующих формированию речных фаций. При этом имел место длительный перенос с сопутствующей мацерацией (размягчением), полное расчленение скелетов и соответственно захоронение преимущественно фрагментов костей[158]. На территории южного Китая провинции Чжэцзян известны два рода — Dongyangopelta и Jinyunpelta относящихся к альбу—сеноману. Они принадлежат к разным семействам анкилозавров. Это предполагает, что два семейства все ещё населяли и представляли собой важное дополнение к известному разнообразию анкилозавров в Азии до того, как нодозавриды были заменены анкилозавридами[47].
Самые обширные находки дошли до нас из верхнего мела. В это время анкилозавры Азии достигают наибольшего разнообразия, где они процветали почти всю вторую половину мелового периода. Из местонахождения Баин-Ширэ (Монголия) известен род и вид Talarurus plicatospineus (сеноман—сантон)[159][160]. Остатки таларуруса (крыша черепа с мозговой коробкой) были обнаружены и несколько юго-западнее, на местонахождении Байшин-Цав, откуда известен род и вид Maleevus disparoserratus. В Амтгае в баинширэинской свите также сделана находка анкилозавра, принадлежащего, судя по строению мозговой коробки, новому роду Amtosaurus. Генезис пород Байшин-Цава и Амтгая большей частью озёрный. Осадконакопление происходило в небольших мелководных водоёмах озёрного типа[161]. Из отложений верхнего мела (формация Джадохта, местонахождение Баин-Дзак) Монголии[143] известен вид Pinacosaurus grangeri. Остатки некоторых пинакозавров, часто образуют массовые скопления. Таковыми являются, например, расположенное несколько западнее Баин-Дзака местонахождение Алаг-Тег (где найдено около сотни скелетов, большинство из которых относятся к незрелыми особям) и лежащее восточнее — Бага-Тариач. Из более высоких горизонтов верхнего мела известны анкилозавриды сайхания и тахия. Оба скелета найдены в формации Баруун-Гоёот около Хулсана. Самый же верхний слой датируется маастрихтом (формация Нэмэгэт, местонахождение Хермин-Цав) и содержит остатки анкилозавров рода сайхания, который является самым последним и совершенным с точки зрения эволюции анкилозавров Азии. Таким образом, очевидно, находки анкилозавров являются неотъемлемым компонентом фаунистического комплекса мела Монголии[162].
Из меловых отложений (кампан; формация Баян Мандаху) Северного Китая известен род Pinacosaurus. В провинции Ляонин в Китае зарегистрировано четыре вида анкилозавров: Liaoningosaurus paradoxus из нижнего мела формации Исянь, Crichtonsaurus bohlini и Crichtonsaurus benxiensis из верхнего мела формации Сунцзявань, а также Chuanqilong chaoyangensis из нижнего мела формации Цзюфотан. Это свидетельствует об относительно высоком разнообразии анкилозавров в меловом периоде на территории современной провинции Ляонин[46].
| Известные рода и виды | |||||
|---|---|---|---|---|---|
| Род | Виды | Семейство | Возраст | Место находки | |
| Amtosaurus Kurzanov and Tumanova, 1978 | Amtosaurus magnus | incertae sedis | 99.7—84.9 | Монголия | |
| Bissektipelta Parish and Barrett, 2004 | Bissektipelta archibaldi | incertae sedis | 94.3—89.3 | Узбекистан | |
| Brachypodosaurus Chakravarti, 1934 | Brachypodosaurus gravis | incertae sedis | 70.6—66.0 | Индия | |
| Chuanqilong Han et al., 2014 | Chuanqilong chaoyangensis | Анкилозавриды | 125.4—112.6 | Китай | |
| Crichtonsaurus Dong, 2002 | Crichtonsaurus benxiensis | Анкилозавриды | 99.7—89.3 | Китай | |
| Gobisaurus Vickaryous et al., 2001 | Gobisaurus domoculus | Анкилозавриды | 94.3—89.3 | Китай | |
| Heishansaurus Bohlin, 1953 | Heishansaurus pachycephalus | Анкилозавриды | 130.0—112.6 | Китай | |
| Jinyunpelta Zheng et al., 2018 | Jinyunpelta sinensis | Анкилозавриды | 112.6—99.7 | Китай | |
| Liaoningosaurus Xu et al., 2001 | Liaoningosaurus paradoxus | Анкилозавриды | 130.0—122.5 | Китай | |
| Qijiangpus Xing et al., 2007 | Qijiangpus sinensis | incertae sedis | 130.0—99.7 | Китай | |
| Pinacosaurus Gilmore, 1933 | Pinacosaurus grangeri Pinacosaurus mephistocephalus | Анкилозавриды | 84.9—70.6 | Китай; Монголия | |
| Saichania Maryanska, 1977 | Saichania chulsanensis | Анкилозавриды | 84.9—70.6 | Монголия | |
| Shamosaurus Tumanova, 1983 | Shamosaurus scutatus | Анкилозавриды | 125.45—99.7 | Монголия | |
| Shanxia Barrett et al., 1998 | Shanxia tianzhenensis | Анкилозавриды | 99.7—66 | Китай | |
| Talarurus Maleev 1952 | Talarurus plicatospineus | Анкилозавриды | 99.7—84.9 | Монголия | |
| Taohelong Yang et al., 2013 | Taohelong jinchengensis | Нодозавриды | 145.5—99.7 | Китай | |
| Tarchia Maryanska, 1977 | Tarchia gigantea Tarchia kielanae Tarchia teresae | Анкилозавриды | 70.6—66 | Монголия | |
| Tianchiasaurus Dong, 1993 | Tianchiasaurus nedegoapeferima | incertae sedis | 175.6—161.2 | Китай | |
| Tianzhenosaurus Pang and Cheng, 1998 | Tianzhenosaurus youngi | Анкилозавриды | 99.7—70.6 | Китай | |
| Tsagantegia Tumanova, 1993 | Tsagantegia longicranialis | Анкилозавриды | 99.7—84.9 | Монголия | |
| Zaraapelta Arbour et al., 2014 | Zaraapelta nomadis | Анкилозавриды | 99.7—84.9 | Монголия | |
| Zhongyuanosaurus Li et al., 2007 | Zhongyuangosaurus luoyangensis | Анкилозавриды | 125.45—99.7 | Китай | |
Северная Америка
В Северной Америке возраст местонахождений анкилозавров лежит в пределах от кимериджа до маастрихта, причём представители семейства нодозаврид встречены начиная с самых нижних отложений киммериджа и до кампана, или самого нижнего маастрихта (формации Эдмонтон)[144]; имеется также единственный зуб из позднего маастрихта, который мог принадлежать нодозавриду[163]. Все находки анкилозаврид в этом регионе приурочены к кампану (формации Ту-Медисин, Джудит Ривер, Олдмэн) и верхнемаастрихтским отложениям[144].
Анкилозавры верхней юры (кимеридж) Колорадо и Вайоминга включают два рода — Gargoyleosaurus и Mymoorapelta, из богатой динозаврами формации Моррисон[95][164]. Эти два рода принадлежат к примитивным анкилозаврам[35]. Нижнемеловая (альб) летопись на североамериканском континенте представлена родами Silvisaurus, Pawpawsaurus и самым ранним анкилозавридом Cedarpelta, найденным на территории штата Юта. Sauropelta известна из нижней части формации Кловерли, возраст которой поздний апт (её остатки также присутствует в вышележащих частях формации Кловерли альбского века)[75]. Ещё один из представителей панцирных динозавров — Gastonia, возможно является самым многочисленным по находкам среди известных анкилозавров. Он представлен тысячами костных остатков, а также несколькими изолированными находками в формации Сидар-Маунтин в восточной части штата Юта. Одно моноспецифическое костное захоронение содержит минимум 22 особи[35][75].
В позднем мелу анкилозавры начинают составляют важный компонент фауны динозавров в северном полушарии, в том числе и в Северной Америке[35]. Анализ анкилозавров кампана и маастрихта из северной Ларамидии показывает большее разнообразие. Известны рода Anodontosaurus, Ankylosaurus, Dyoplosaurus, Euoplocephalus, Oohkotokia, Scolosaurus, Zuul. Все они найдены на территории штата Монтана, США и провинции Альберта, Канада[165]. Присутствие различных видов анкилозавров в тесной хронологической и географической близости в северной Ларамидии, учитывая различную палеосреду, предполагает, что близкородственные виды анкилозавров могли предпочитать разные среды обитания[166]. Свидетельства анкилозавров из южной Ларамидии были редки к моменту открытия Nodocephalosaurus. Однако за последние 15 лет после его описания число позднемеловых таксонов анкилозавров резко увеличилось. К новым южноларамидийским анкилозаврам кампана и маастрихта были отнесены Ahshislepelta, Glyptodontopelta, Invictarx и Ziapelta, остатки которых были найдены на территории штата Нью-Мексико, а также Akainacephalus найденный на территории Юты. Это привело к быстрому увеличению видового разнообразия в пределах южной Ларамидии в позднем мелу западной части Северной Америки. Несмотря на эти открытия из Нью-Мексико и Юты, позднемеловые южноларамидийские анкилозавры остаются редки, к тому же большинство видов представлены одним экземпляром[167]. В отличие от богатой летописи окаменелостей восточной части Ларамидии, анкилозавры западного побережья субконтинента почти неизвестны. Найдены окаменелости анкилозавра из Калифорнии — Aletopelta coombsi формации Пойнт-Лома, а также частичный череп анкилозавра Edmontonia из морской формации Матануска на юге центральной Аляски[168]. Остатки наиболее поздней формы Северной Америки относятся к эдмонтонии и встречаются в верхнемаастрихтских формациях Ланс, Хелл-Крик и Ларами[169].
Высокое разнообразие анкилозавров в кампане—маастрихте Ларамидии согласуется с высоким морфологическим и видовым разнообразием, наблюдаемым у других групп динозавров, особенно гадрозаврид и цератопсид, однако последние всё же значительно более разнообразны[166]. Присутствие анкилозаврид, тесно связанных с монгольскими видами, в позднем кампане Ларамидии, даёт возможность предполагать некоторую палеогеографическую связь и межконтинентальный обмен видами между Азией и западом Северной Америки в это время или несколько раньше[167][170].
| Известные рода и виды | |||||
|---|---|---|---|---|---|
| Род | Виды | Семейство | Возраст | Место находки | |
| Ahshislepelta Burns and Sullivan, 2011 | Ahshislepelta minor | Анкилозавриды | 84.9—70.6 | США (Нью-Мехико) | |
| Aletopelta Ford and Kirkland, 2001 | Aletopelta coombsi | incertae sedis | 84.9—70.6 | США (Калифорния) | |
| Akainacephalus Wiersma and Irmis, 2018 | Akainacephalus johnsoni | Анкилозавриды | 84.9—70.6 | США (Юта) | |
| Anodontosaurus Sternberg, 1929 | Anodontosaurus inceptus Anodontosaurus lambei | Анкилозавриды | 70.6—66 | Канада (Альберта) | |
| Ankylosaurus Brown, 1908 | Ankylosaurus magniventris | Анкилозавриды | 70.6—66 | Канада (Альберта, Саскачеван) США (Монтана, Нью-Мехико, Вайоминг) | |
| Cedarpelta Carpenter et al., 2001 | Cedarpelta bilbeyhallorum | Анкилозавриды | 122.46—109 | США (Юта) | |
| Dyoplosaurus Parks, 1924 | Dyoplosaurus acutosquameus | Анкилозавриды | 84.9—70.6 | Канада (Альберта) | |
| Euoplocephalus Lambe, 1910 | Euoplocephalus tutus | Анкилозавриды | 70.6—66 | Канада (Альберта); США (Монтана) | |
| Gastonia Kirkland, 1998 | Gastonia burgei Gastonia lorriemcwhinneyae | Полакантиды | 140.2—125.4 | США (Юта) | |
| Niobrarasaurus Carpenter et al., 1995 | Niobrarasaurus coleii | incertae sedis | 85.8—84.9 | США (Канзас) | |
| Nodocephalosaurus Sullivan, 1999 | Nodocephalosaurus kirtlandensis | Анкилозавриды | 84.9—70.6 | США (Нью-Мехико) | |
| Oohkotokia Penkalski, 2014 | Oohkotokia horneri | Анкилозавриды | 84.9—70.6 | США (Монтана) | |
| Palaeoscincus Leidy, 1856 | Palaeoscincus asper Palaeoscincus costatus Palaeoscincus latus Palaeoscincus magoder | incertae sedis | 84.9—70.6 | Канада (Альберта); США (Монтана) | |
| Platypelta Penkalski, 2018 | Platypelta coombsi | Анкилозавриды | 84.9—70.6 | Канада (Альберта) | |
| Scolosaurus Nopsca, 1928 | Scolosaurus cutleri Scolosaurus thronus | Анкилозавриды | 84.9—70.6 | Канада (Альберта) | |
| Stegopelta Williston, 1905 | Stegopelta landerensis | incertae sedis | 105.3—94.3 | США (Вайоминг) | |
| Ziapelta Arbour et al., 2014 | Ziapelta sanjuanensis | Анкилозавриды | 84.9—70.6 | США (Нью-Мехико) | |
| Zuul Arbour and Evans, 2017 | Zuul crurivastator | Анкилозавриды | 84.9—70.6 | США (Монтана) | |
Южная Америка
Ископаемые свидетельства присутствия анкилозавров в Южной Америке немногочисленны, но устойчивы. Несмотря на то, что в Южной Америке нет заявленных видов, количество окаменелостей анкилозавров увеличилось за последние годы. Самые раннее свидетельства существования анкилозавров в западной Гондване относятся к следам верхней юры формации Гуара (бассейн Парана, южная Бразилия)[171], а также к следам верхней юры—нижнего мела близ деревни Икла, в 100 км к юго-востоку от Сукре, Боливия. Остальные неопределённые экземпляры нодозаврид, некоторые изолированные остеодермы и множество следов известны только из верхнего мела (кампан—маастрихт)[172]. Свидетельствами существования анкилозавров позднего мела в Южной Америке являются обнаруженные следы в Боливии, близ города Сукре на горе Каль Орко[173]. На основе этих находок швейцарский палеонтолог Кристиан Майер выдвинул предположения, что анкилозавры могли прибыть сюда из Северной Америки в позднем юрском или раннем меловом периоде. Также помимо этого он выдвигает теорию, что анкилозавры могли прийти сюда в меловом периоде из Африки[174]. Помимо этого в Боливии, в местонахождении Торо-Торо, били найдены следы динозавров, предположительно относящиеся к анкилозаврам. Ихнотаксону было дано имя Ligabueichnium bolivianum[175]. Наличие разнообразия следов в Боливии даёт возможность исследователям предполагать, что они являются частью маршрута сезонной миграции вдоль береговой линии и древней озёрной системы существовавшей в позднемеловую эпоху[176]. Есть также задокументированные свидетельства существования анкилозавров позднего мела на территории нынешних Аргентины[177][178] и Бразилии[179].
Австралия и Антарктида
Австралия в настоящее время представлена наиболее сохранным и богатым ископаемым материалом анкилозавров среди южных континентов. Самые ранние находки анкилозавров в Австралии указывают на то, что эта группа динозавров присутствовала на континенте по крайней мере уже во времена позднего альба—сеномана. Раннемеловые свидетельства панцирных динозавров фрагментарны и известны из местонахождений в штатах Квинсленд и Южная Виктория[180]. Наиболее полные находки анкилозавров Австралии ограничиваются остатками из отложений нижнего мела западного Квинсленда — Minmi и Kunbarrasaurus. В раннем меловом периоде часть Квинсленда образовала большой остров, отдельный от остальной части Австралии. Окружающая среда интерпретируется как смесь пойм и лесов[181]. Окаменелости анкилозавра также были обнаружены и в Новой Зеландии в формации Тахора (англ. Tahora Formation; несколько позвонков и фрагмент ребра) относящейся к кампану. В это время Новая Зеландия была островом недалеко от Антарктиды, а динозавры данной местности представляли собой островную полярную фауну. Фауна отличается от других хорошо известных меловых фаун Гондваны, но похожа на фауну из нижнего мела Квинсленда. Предполагается, что происхождение этой фауны связано с заселением из Антарктиды в то время, когда Новая Зеландия ещё не покинула её[147][182].
Летопись окаменелостей анкилозавров Антарктиды является достаточно бедной. Малочисленность находок частично объясняется нахождением её поверхности подо льдом. Все находки анкилозавров Антарктиды относятся к морским и прибрежным отложениям верхнего мела (кампан—маастрихт)[183]. Известен единственный вид — Antarctopelta oliveroi[184]. Также в Антарктиде найдены относящиеся к анкилозаврам фрагменты нижней челюсти, зубы, остеодермы, фрагмент хвостовой «булавы». Предполагается, что расселение анкилозаров в Антарктиду могло произойти по существовавшему маршруту расселения динозавров из Южной Америки в позднем мелу (кампан—ранний маастрихт) или немного ранее — в туронском веке. Учитывая, что остеодерма неизвестного нодозаврида была найдена в верхних слоях маастрихта, она представляет собой самую молодую запись анкилозавров из Антарктиды, давая возможность предполагать, что панцирные динозавры сохранились на континенте до конца мелового периода[183].
| Известные рода и виды | |||||
|---|---|---|---|---|---|
| Род | Виды | Семейство | Возраст | Место находки | |
| Antarctopelta Salgado and Gasparini, 2006 | Antarctopelta oliveroi | Нодозавриды | Кампан—Маастрихт | Антарктида | |
| Kunbarrasaurus Leahey et al. 2015 | Kunbarrasaurus ieversi | incertae sedis | Альб | Австралия | |
| Minmi Molnar, 1980 | Minmi paravertebra | Анкилозавриды | Валанжин—Апт | Австралия | |
На территории России
На территории России нет заявленных родов и видов. Находки анкилозавров очень бедны, представлены исключительно фрагментарными остатками и в силу характера их сохранности они не могут быть идентифицированы на родовом уровне. Это связано с тем, что на протяжении длительной меловой истории Поволжье и значительная часть центральной России находились под водой. Лишь в конце меловой эпохи часть территории Поволжья освободилась от морских вод[185]. В связи с этим основные находки анкилозавров на территории России расположены за Уралом, где большая часть также труднодоступна — занята лесами, тундрой, болотами, зоной вечной мерзлоты. Самые ранние остатки панцирных динозавров относятся к нижнему мелу (берриас—баррем), обнаружены на территории Якутии на местонахождении Тээтэ[112]. В Волгоградской области близ хутора Полунино известны фрагменты щитков из верхнего мела (кампан—маастрихт)[100]. В местонахождении Кундур (ранний маастрихт), расположенном на юго-востоке Амурской области (Архаринский район) находки анкилозавров состоят из двух частично разрушенных зубов и шипообразной остеодермы[169]. Также о нахождении фрагментов на реке Амур, принадлежащих анкилозаврам, сообщает А. К. Рождественский, относя их к маастрихтскому возрасту[186]. Зубы, обнаруженные в местонахождении Каканаут (поздний маастрихт) на северо-востоке России в Чукотском автономном округе, очень напоминают зубы североамериканского нодозаврида — эдмонтонии, также найденного в высоких широтах на Аляске. Однако одних лишь зубов недостаточно, чтобы достоверно отнести находки к какому-либо роду, и потому зубы, обнаруженные в Каканауте, относятся к неопределённым анкилозаврам — Ankylosauria indet.[187]
Физиология
Питание и пищеварение

Исторически считалось, что рацион анкилозавров преобладает или состоит исключительно из растительного материала[129][188]. Из-за сочетания морфологии черепа и относительно небольших и простых зубов многие авторы также предполагали, что их диета была ограничена мягким, неабразивным растительным материалом[139][189]. Многочисленные исследования палеоэокологии анкилозавров дали относительно полные данные о специфике питания анкилозавров и их пищевых предпочтениях. Низкое расположение головы и телосложение, не позволявшее анкилозаврам подниматься на задних лапах, указывают на то, что пищу анкилозавры искали близко к земле — примерно до 1 метра в высоту[93][190]. По ширине челюстей анкилозавров можно делать выводы о различии диет разных семейств[191]. Заострённые морды нодозаврид указывают на выборочное поедании более высококачественных растений. Анкилозавриды обычно имели более широкие морды и челюсти, что предполагает поедание как можно большего количества корма, независимо от качества растений[69][192]. Существование анкилозавров около ста миллионов лет на большинстве, если не на всех, континентах даёт возможность предполагать, что их пищевые растения сильно отличались с течением времени. В юре это часто были низкорослые папоротники и голосеменные растения, такие как саговники и беннеттитовые[193]. В меловом периоде появляются цветковые растения, ставшие важной частью мировой флоры[194]. Анкилозавры могли перейти на данный вид растительности, в то же время продолжая поедать папоротники и низкорослые кустарники, которые были легко доступны в позднем меловом периоде[20]. Быстрый рост и сильный восстановительный потенциал цветущих растений, возможно объясняет большое разнообразие анкилозавров в этот период[194]. Предположение Кумбса о том, что эти животные могли копать землю в поисках пищи[77], опровергается широкими когтями, которые, вероятно, не подходят для такой деятельности[3]. Анкилозавры также не были способны жевать жёсткую кору деревьев, как например современные слоны[195]. Учитывая их сравнительно слабое жевание, время на поиск пищи могло занимать значительную часть дня[20].
В 2017 году Виктория Меган Арбор и Джордан Мэллон попытались подсчитать количество потребляемой пищи на примере рода анкилозавр. По их оценкам количество потребления сухих папоротников составило около 6 кг в день (около 2 тонн в год). Виктория Арбор посчитала эти цифры слишком малыми и предположила, что дополнительные потребности анкилозавров в питательных веществах могли быть получены за счёт потребления богатыми энергией плодовых тел. Маленькие беспозвоночные также могли иногда дополнять рацион[20]. В результате новых исследований, предполагающих наличие длинного мускулистого языка (в ротовой полости имеется сильно развитый подъязычный аппарат[196]), а также недавнее открытие скелета Liaoningosaurus с рыбой, сохранившейся в его грудной полости[197], Виктория Арбор делает заключение, что растительная пища могла занимать не весь рацион анкилозавров[20]. Впервые частично возможный насекомоядный рацион анкилозавров предположил Франц Нопча[198]. Такого же мнения придерживаются и ряд других исследователей[23][6]. Вероятное присутствие якобсонова органа[23][54] также говорит в пользу того, что рацион анкилозавров возможно не был ограничен только растениями, хотя последние, несомненно, составляли его значительную часть[6]. Рождественский предполагал, что анкилозавры могли питаться яйцами других динозавров. Эту точку зрения поддерживал и Лев Несов, считая что вне периода размножения других динозавров анкилозавры могли быть копрофагами или всеядными формами, однако относительно долгий период инкубации крупных яиц динозавров мог давать возможность анкилозаврам значительную часть годового цикла быть обеспеченными пищей[100].
На сегодняшний день панцирные динозавры сохранили самые большие свидетельства о содержимом желудка — кололит, чем любая другая группа растительноядных динозавров[199]. Найденный на территории Австралии род Kunbarrasaurus дал возможность увидеть, чем питались анкилозавры. Помимо скелета, в области желудка были найдены следы последнего приёма пищи животного: папоротника, плодов покрытосеменных, мелких семян и других растений с мягкими листьями[200][201]. Другой вид анкилозавра — Borealopelta, из Северной Америки, предпочитал папоротники группы лептоспорангиатные взамен других, и ел очень мало саговников и хвойных листьев[199]. Дэвис Фастовский в своей книге приводит подробный процесс потребления и переваривания пищи у анкилозавров. Клюв животного откусывал крупные части растения и они были разрезаны рядами маленьких листообразных зубов верхней челюсти на более мелкие кусочки, что согласуется со строением челюстей и ротовой полости анкилозавров. Мясистые щёки удерживали растительную массу на месте[202]. У анкилозавров также было большое вторичное нёбо, это означает, что они могли дышать во время жевания, в отличие от крокодилов[65]. Полноценный жевательный процесс был невозможен из-за ограниченной силы укуса[202], которая была в ниже, чем у цератопсид[203], в основном из-за короноидного отростка нижней челюсти, позволявший только умеренное усилие при закрытии челюсти. Таким образом, части растений были проглочены довольно большими кусками и, возможно, размолоты гастролитами в желудке — камни, которые были обнаружены у Panoplosaurus[62] и Borealopelta[199]. Широкий торс и видоизменённый таз — признаки наличия объёмной пищеварительной системы. Вполне возможно, что ферментация происходила с помощью симбиотических микроорганизмов в многокамерном желудке или в другом отделе пищеварительного тракта[200][202]. Предполагается также, что анкилозавры могли влиять на экостистемы, способствуя распространению папоротников из-за того, что их огромное количество фекалий представляли собой непереваренные спорангии, становясь таким образом богатым удобрением для гаметофитов, следующей фазы в жизненном цикле высших растений[201].

Вопрос, насколько эффективным был у анкилозавров жевательный аппарат, остаётся спорным. Долгое время считалось, что челюсти анкилозавров были в состоянии осуществлять лишь режущие движения в вертикальной плоскости с помощью ряда мелко зазубренных по краю зубов, на которых у большинства анкилозавров не обнаружено сколько-нибудь достоверных следов стирания. Животное срывало растения посредством острых, загнутых в виде тупого клюва, с помощью примитивной зубной системы мелко настригало мягкую растительность, не наносящую повреждений эмали зубов[6]. Георг Хаас в 1969 году провёл исследование мускул челюсти двух черепов анкилозавров и пришёл к выводу, что несмотря на большой размер черепов, связанная с ними мускулатура была относительно слабой. Он также заключил, что движения челюстей в основном ортогональны, и работали только в вертикальной плоскости. В результате Хаас сделал вывод, что анкилозавры, такие как Euoplocephalus, вероятно ели относительно мягкую, неабразивную растительность[189]. Однако более поздние исследования показали, что движение челюстей в горизонтальной плоскости были возможны, а также указали на их способность выдерживать значительные нагрузки. Микроскопическое исследование следов износа зубов у рода Euoplocephalus показало, что он был в состоянии делать более сложные движения. Это свидетельствовало о том, что по крайней мере у некоторых анкилозавров движения челюстей были приспособлены к более тщательному пережёвыванию пищи[204]. Количественный анализ черепов показал, что у базальных членов анкилозавров обработка пищи ограничивалась простым вертикальным движением челюстей, в раннем и позднем меловом периодах Северной Америки, Европы и Азии у некоторых видов нодозаврид и анкилозаврид эти движения дополнились горизонтальными движениями. Это может быть связано с различиями сред обитания и различиями в типах растительности близкородственных таксонов[205].
Передвижение
Окаменелости скелетов и палеоихнология (наука о ископаемых следах) показывают, что анкилозавры передвигались исключительно на четырёх конечностях[77][206]. Низкие остистые отростки крестцовых позвонков указывают на то, что анкилозавры не могли стоять на задних лапах. Принято считать, что анкилозавры были относительно медленными животными[207]. Тяжёлое телосложение, строение поясов и коротких конечностей говорит об отсутствии каких-либо приспособлений у анкилозавров к быстрому передвижению[24][65]. В своей работе Уолтер Кумбс перечислил характерные черты адаптации к бегу среди динозавров и оценил уровень передвижения анкилозавров от «низкого до среднего»[207].
Согласно расчётам Ричарда Энтони Тулборна в 1982 году, основанным на длине конечностей и предполагаемой массе тела, эти животные не могли развивать скорость более 6—8 км/ч[208]. Швейцарский палеонтолог Кристиан Майер определили максимальную скорость передвижения 11 км/ч при весе животного в 8 тонн. Такой точный расчёт стал возможен благодаря формуле британского учёного Александера: скорость динозавра вычисляют по высоте его бедра и удвоенной длине шага[174]. Тем не менее, основываясь на тропах окаменелых следов приписываемых анкилозаврам, исследователи вычислили, что средняя скорость передвижения равнялась 2,7—3,6 км/ч[209]. Поскольку конечности анкилозавров были короткими, чтобы достичь более высокой скорости, им приходилось на некоторое время отрываться от земли во время своего цикла передвижения. На это указывают согнутые конечности в области коленного сустава, скакательный сустав и пяточная кость, сросшаяся с большеберцовой костью, а также очень тяжёлая мускулатура. Учитывая то, что у большинства видов конечности располагались далеко друг от друга в продольном направлении, то такое движение было бы очень неэффективным и прирост скорости очень ограничен.
Однако тот факт, что анкилозавры не имели высокой скорости, не означает, что они были чрезвычайно медленными. Иногда применяя своего рода «модель черепахи» исследователи предполагают, что фактический вес панциря не был чрезмерным, поскольку остеодермы имели небольшую толщину, где лёгкая губчатая кость составляла большую их часть, особенно у нодозаврид[40]. Панцирь анкилозавров обычно составляет 10 % массы тела. Дополнительный вес с лихвой компенсировался огромной мускулатурой. Длинные подвздошные кости несли очень широкие мышцы бедра, а крайнее верхнее расширение плечевой кости на передних конечностях также указывает на наличие мощной мускулатуры. Это должно было позволить телу значительно ускориться в критических ситуациях.
Метаболизм
Структура костей анкилозавров указывает на умеренную температуру их тела, что в свою очередь говорит о довольно низком метаболизм. Это свидетельствует о не слишком активный образе жизни, при котором анкилозавры были спокойно передвигающимися по ландшафту формами. Снижение метаболизма не исключает непродолжительных, очень энергичных усилий. При низком метаболизме нет оснований думать о хладнокровности анкилозавров в том смысле, что животное могло зависеть от внешних источников тепла для поддержания температуры своего тела. Исследования показывают на способность анкилозавров сохранять постоянную температуру тела, независимо от температуры окружающей среды, с возможностью гетеротермии в зависимости от региона обитания[210]. Грегори С. Пол также указывает о наличии признаков гетеротермии у анкилозавров[211]. Мышечная активность и пищеварение в большой брюшной полости могли генерировать достаточно много энергии, которая позволяла хорошо удерживать большое тело. Так что хоть и была массовая эндотермия, но и метаболизм, генерируемой постоянной клеточной активность, не мог быть слишком низким. На это указывает открытие рода Minmi, обитавшего за Полярным кругом, которому в прохладную арктическую ночь в течение шести месяцев удавалось поддерживать приемлемую температуру тела. Исследователи не исключает возможности зимовки анкилозавров, обитавших в высоких широтах[211][212].
Мозг
Об интеллекте анкилозавров можно судить по структуре их мозговой коробки[69]. Форма и строение мозговой оболочки анкилозавров плохо изучены из-за достаточно сильного окостенения черепов[213]. Для изучения полости мозга применяются компьютерные томографии и слепки, называемые эндокастом. Эти слепки приблизительно соответствует размеру и форме мозга, так как мозг не заполняет полость полностью. Исследование эндокастов показали, что область переднего мозга (в основном головной мозг) была довольно маленькой, включая обонятельные доли, так что даже несмотря на наличие больших ноздрей у анкилозавров их обоняние не было очень острым. В области среднего мозга зрительные нервы не очень заметны, что позволяет предположить, что зрение тоже было не очень хорошим (примерно как у современных носорогов). Область гипофиза велика для размера эндокаста, но это характерно для большинства динозавров, независимо от размера. Задний мозг (мозжечок и продолговатый мозг) контролирует баланс, координацию движений и органы тела (сердце, лёгкие, печень, кишечник и т. д.). По сравнению с ныне живущими рептилиями, размер мозга у анкилозавров был примерно в два раза меньше[69]. Исследование Дж. А. Хопсона показало, что по сравнению с размерами тела, он был пропорционально меньше, чем у стегозавров[214]. Из всех динозавров только у завроподов был меньший мозг[193]. Маленький мозг анкилозавров соответствует присущему им менее активному образу жизни[215]. Некоторые более поздние исследования изучили форму мозга родов Panoplosaurus и Pinacosaurus. Оказалось, что мозг занимал примерно ту же часть черепа, что и у других птицетазовых динозавров. У анкилозавра длиной пять метров объём мозга составлял около 70 см³. Этот размер мозга сам по себе достаточен для сложных движений. Благодаря исследованию мозговой полости вида Bissektipelta archibaldi учёным удалось узнать, что масса мозга данного вида составляла около 26,5 грамма для трёхметровой особи, размер 8,5 см, а также, что по крайне мере у некоторых видов немалую часть мозга занимали обонятельные луковицы — около 60 % от размера больших полушарий. Также возможно, кровь мозга могла течь в разных направлениях и перераспределяться, сохраняя оптимальную температуру мозга животного. Устройство кровеносной системы в голове анкилозавров было больше похоже на сосуды современных ящериц, чем на более близких современных родственников динозавров — крокодилов или птиц[216].
Обоняние и слух
Некоторые виды анкилозавров (например Bissektipelta) обладали достаточно хорошим обонянием, которое, вероятно, помогало им искать пищу, сородичей противоположного пола и вовремя чувствовать приближение хищников. Нюх анкилозавров даже можно сравнить с нюхом известного хищника тираннозавра рекса — его обонятельные луковицы были всё же больше на 15 %, около 65-70 % от размеров больших полушарий. Также звуковой диапазон ранних анкилозавров составляет примерно от 300 до 3000 герц, — в таком же диапазоне слышат современные крокодилы. Это достаточно низкие частоты, которые соответствуют сравнительно крупным размерам анкилозавров. Чем крупнее современные животные, тем более низкочастотные звуки они издают и слышат. Палеонтологи предполагают, что в процессе эволюции анкилозавры увеличивались в размерах, поэтому более поздние формы воспринимали звуки ещё более низких частот[216]. Исследования также показали, что нодозавриды и анкилозавриды обладали разным слухом[213].
Кожные покровы
Почти все сохранившиеся до нашего времени ископаемые остатки анкилозавров не содержат кожных покровов, окаменелые оттиски эпидермиса мягких тканей встречаются редко. Тем не менее, хорошо сохранившиеся покровы существуют на нескольких окаменелостях анкилозавров, которые показывают, что остеодермы были покрыты эпидермальной чешуёй. Относительно многочисленные остеодермы также предоставляют очень важную информацию о структуре кожных покровов анкилозавров[217]. Предполагается, что при жизни эти костные структуры анкилозавров не подвергались воздействию внешней среды, так как несли на себе ороговевшие эпидермальные чешуйки[37][218]. В этом плане открытие вида Borealopelta markmitchelli, несущего почти полный набор передних остеодерм и связанных с ними ороговевших чешуек и оболочек, дало уникальную возможность задокументировать, как костные части остеодерм, так и уникальные эпидермальные чешуйки, покрывающих остеодермы. Исключительная сохранность эпидермальных структур значительно расширила данные об эпидермисе анкилозавров. Во-первых, этот образец даёт доказательства наличия утолщённых, ороговевших покровов почти на всех остеодермах, не оставляя никаких вопросов относительно повсеместного наличия этой ткани. В сочетании с другими проявлениями более изолированных сохранившихся ороговевших остеодермальных покровов у анкилозаврид, это даёт широкую поддержку мнению, что все остеодермы анкилозавров не были ни голыми, ни покрытыми тонкой тканью, а были покрыты утолщённым кератиновым слоем. Соотнося ископаемый материал рода Borealopelta с предыдущими находками, можно сделать вывод — это первое появление ороговевшего эпидермиса у анкилозавров[37].
На основе всё того же рода Borealopelta исследователи смогли идентифицировать меланин в органических остатках ископаемого с помощью спектроскопических анализов, а также обнаружили более светлую пигментацию больших шипов, что говорит о наличии окраса «противотени» по всему телу — переход от более тёмного верха к светлому низу, искажая зрительное восприятие объекта и делая его менее заметным. Сами покровы были окрашены меланином в красновато-коричневый цвет. Это первый пример возможной защитной окраски анкилозавров, как один из способов противодействия хищным динозаврам. Присутствие «противотени» у анкилозавров предполагает, что давление хищников было достаточно сильное и дало толчок в сторону приобретения защитной окраски у этих крупных растительноядных, несмотря на наличие массивной спинной и боковой костной «брони»[219].
Воспроизведение и развитие
Анкилозавры, как и большинство динозавров[220], вероятно, откладывали яйца, но в настоящее время нет ни одной находки яиц, которые можно было бы с точностью отнести к данной группе. Выдвигаются предположения, что ископаемые яйца Cairanoolithus, найденные в Европе, могут принадлежать нодозавриду рода Struthiosaurus. Несмотря на то, что представители данного рода были относительно небольшого размера, необычная форма их седалищной кости вполне позволяла им легко откладывать яйца большого размера (размер Cairanoolithus составляет 19 см). Однако данная интерпретация ископаемого яйца, как возможно принадлежащего нодозавридам, поднимает вопрос, почему подобные яйца не обнаружены в тех регионах, где нодозавриды были более многочисленны, таких как Северная Америка. Отсутствие такой записи может быть связано с тафономическими факторами, это может отражаться в преобладание у нодозаврид яиц с мягкой скорлупой или особом репродуктивном поведении у нодозаврид, как предполагается для большинства птицетазовых динозавров[221].
Также по сравнению с другими группами динозавров, известно очень мало окаменелостей молодых особей[222]. Полностью сохранившаяся окаменелость молодой особи, длиной 34 сантиметра, дошедшая до нас — Liaoningosaurus. Он значительно отличается от всех других анкилозавров рядом особенностей, включая наличие защиты в брюшной области, острых когтей и относительно больших зубов. Неясно, были ли это общие характеристики молодняка или родовые характеристики[26]. Известна и частично сохранившаяся в виде естественных слепков окаменелость детёныша нодозаврида — Propanoplosaurus marylandicus, настолько маленького, что можно интерпретировать его как недавно вылупившегося. Несмотря на довольно необычный тип сохранности, находку правильно называть окаменелостью тела, а не ихнитом (ископаемым отпечатком) как таковым, потому что она фиксирует не действие организма, а форму тела и структуру костей (включая частичное сочленение) динозавра. Находка является единственным прямым свидетельством вылупления и вероятного гнездования анкилозавров[223].
Очень мало известно и о стадиях роста у анкилозавров. Гистология их костей указывает на медленные темпы роста по сравнению с другими видами динозавров. Внутри группы более высокие темпы роста обнаружены у анкилозаврид, в то время как нодозавриды показали более медленные темпы роста, схожие с таковым у стегозавров. Относительно быстрый рост у анкилозавров происходил на стадии от раннего развития организма до достижения половой зрелости, затем отмечается замедление темпов роста. Относительно высокая скорость роста молоди может быть вызвана адаптацией к сокращению времени созревания костного панциря к моменту достижения половой зрелости особи. На поздней стадии развития быстрый рост попросту не имел решающего значения из-за сформировавшейся защиты и темпы роста могли снижаться. По-видимому анкилозавры, как и стегозавры, имели более высокие темпы роста, чем предполагаемый их предок — скутеллозавр. Тем не менее, по сравнению с другими видами динозавров (такими как орнитоподы, цератопсы, тероподы и зауроподоморфы), анкилозавры в частности, и тиреофоры в целом, росли медленнее[224].
Палеобиология
Среда обитания

Об образе жизни анкилозавров существует несколько точек зрения. С одной стороны, их рассматривают как животных сугубо наземных, связанных с водоёмами лишь потребностью в питье и сборе пищи в прибрежных заводях. С другой стороны, некоторые черты их строения, в первую очередь неуклюжесть, неповоротливость, свидетельствуют о более тесной связи их с водой. Большинство находок анкилозавров указывают на то, что эти животные предпочитали влажные среды обитания с богатой растительностью, такие как районы рек и озёр, прибрежные равнины[5][173]. В строении анкилозавров проявляются морфологические признаки, которые в комплексе могут указывать на их полуводный образ жизни. Это бочонковидная или сильно уплощённая форма туловища, мелкие зубы, способные измельчать только мягкую водную растительность; широкие плоские копытообразные широко расставленные концевые фаланги пальцев для передвижения по мягкому грунту. Систематичность находок массовых скоплений анкилозавров Азии в местах, представлявших собой обезвоживающуюся часть берега может быть объяснена, скорее всего, тесной связью жизни животных с водой. Подобная концентрация особей в одном местонахождении даёт основательный повод полагать, что анкилозавры вели полуводный образ жизни[5]. Эти предположения ещё сильнее подкрепились находкой вида Liaoningosaurus paradoxus. Наличие остатков рыбы в желудке, длинных конечностей и острых когтей предполагают вероятную адаптацию к полуводному образу жизни[26].
Многие находки нодозаврид в Северной Америке часто относятся к прибрежным морским отложениям, иногда окаменелости залегают в породах представлявших в древности открытую морскую среду, например Niobrarasaurus, Pawpawsaurus[107][152]. Высказываются предположения, что некоторые североамериканские формы могли обитать в прибрежных морских зона или по крайней мере недалеко от моря[152]. В то же время существуют мнения, что не всегда имеется связь между окаменелостью и средой обитания. Нахождение окаменелостей в морских отложениях может означать всего лишь то, что трупы анкилозавров были перенесены в море из их естественных прибрежно-равнинных мест обитания уже после смерти[152][225]. Также существуют предположения, что шансы превратиться в ископаемые и сохраниться до наших дней в таких местах намного выше. Редкие находки анкилозаврид в морских отложениях дают возможность предполагать предпочтение ими более внутренних или горных мест обитания[226], однако исследование 2016 года не обнаружило однозначной связи между местом находок и сред обитания[107].
То, что анкилозавры могли жить в засушливых и полузасушливых средах, очевидно из множества находок анкилозаврид в пустыне Гоби на территории современных Монголии и Китая, где в меловом периоде также была пустыня[56]. Очень сложная носовая полость этих животных (например у Euoplocephalus), рассматривается как приспособленность к климату пустынь. Основная гипотеза предполагает, что сложные полости носа и пазух могли использоваться для охлаждения или увлажнения воздуха[23][66]. Помимо этого следы присутствия анкилозавров обнаружены в высоких широтах (более 60˚) обоих полушарий, где биологическая продуктивность сдерживалась коротким вегетационным периодом, низкими температурами и ограниченной доступностью питательных веществ[103]. Возможно, именно их приспособляемость к множеству условий окружающей среды привела к долголетию данной группы в течение 100 миллионов лет[16].
Функции панциря и защита
Анкилозавры, как и другие растительноядные динозавры, служили потенциальной добычей для хищников. Все крупные плотоядные динозавры, которые могли преследовать и атаковать анкилозавров на суше, относятся к группе теропод. Учёные выяснили, что даже такой крупный и тяжёлый хищник, как тираннозавр, был гораздо быстрее анкилозавров и легко мог настигать их на открытом пространстве[227]. Классическая интерпретация функции «брони» состоит в том, что она служила им пассивной защитой[56][131]. Предполагается, что анкилозавры, у которых отсутствовала большая костная «булава» на конце хвоста, столкнувшись с крупным тероподом, прижимались к земле и втягивали конечности[228]. Плоский, защищённый панцирем верх, предлагал атакующему хищнику не так много точек атаки. Более уязвимые части туловища (например шея) были покрыты острыми шипами[27]. У ранних форм задняя часть тела часто прикрыта бедренным щитом[20]. Голова анкилозавров также могла быть атакована хищниками и обладала высокой степенью окостенения. Обороне способствовал тот факт, что череп и нижняя челюсть сами собой образовывали твёрдые костные структуры. У анкилозавров это было дополнительно усилено закрытием большинства отверстий черепа, слиянием различных элементов и в целом компактной структурой, особенно у анкилозаврид. Кроме того область глазниц и боковая часть нижней челюсти были защищены наросшими пластинами. У рода Euoplocephalus были обнаружены окостеневшие веки[61]. Ранее выдвигались предположения, что из-за их чрезвычайно коротких голов хвостовая «булава» анкилозаврид могла использоваться как отвлечение: тероподы по ошибке могли атаковать «булаву», а не голову[3][229]. Сегодня данная гипотеза уже не распространена[230]. Плоское строение туловища также уменьшало вероятность того, что крупный теропод перекатит анкилозавра на спину, с целью атаковать брюшную полость[231]. Пассивная защита могла заключаться также в поведении, предполагающее демонстрацию хищнику своих шипов (например у рода Sauropelta были достаточно длинные шипы в области предплечья), тем самым сигнализируя об угрозе со своей стороны. В то же время большинство анкилозавров не обладали наличием больших шипов, что позволяет предположить, демонстрация и угроза хищникам не имела большого значения для этих животных. Вместо этого, возможно, он издавали сильное шипение или другие звуки. Также возможно, что анкилозавры могли менять цвет своего покрова. Губчатые кости их остеодерм предполагают наличие множества сосудов, указывая на усиленное кровоснабжение, в результате чего кожа над остеодермами могла приобретать красноватый оттенок[232].
Хотя анкилозавры, несомненно, часто защищали себя таким образом, есть основания сомневаться в том, что они всегда были пассивными. Хвост нодозаврид был достаточно подвижным, покрытый остеодермальными шипами, и также мог служить эффективным оружием защиты[44]. У некоторых видов анкилозаврид хвостовой конец превратился в тяжёлую костную «булаву»[46], удар которой был достаточно мощным, чтобы сломать кости небольшому атакующему тероподу[42][233], или опрокинуть его на землю[234]. И хотя у анкилозаврид уже имеется пассивная защита в виде панциря, трудно пока найти другое назначение столь своеобразной конструкции на хвосте[27][144]. Более крупные нодозавриды должно быть могли таранить нападающих хищников. Большие тероподы могли запросто быть опрокинуты из-за своего двуногого способа передвижения, если динозавр весом в несколько тонн ударит по их конечностям[235]. Эффект от этого мог быть усилен наличием на плечах анкилозавров очень острых шипов[56].
При изучении возможностей защиты анкилозавров, часто пытались обнаружить их слабые места. Одна из гипотез недостатка защиты была основана на неуклюжести анкилозавров. Эту гипотезу впервые предложил Франц Нопча в 1928 году после изучения перевёрнутого образца сколозавра. Он предположил, что частое появление скелетов анкилозавров, лежащих на спине, возможно было вызвано смертью животных в результате случайного переворачивания при падения с холма, где они лежали не имея возможности встать[198]. Известно, что современных черепах иногда постигает та же участь[236]. Кеннет Карпентер отметил, что модель неуклюжести достаточно слаба, указывая на то, что прибрежные равнины Северной Америки, где перевёрнутые анкилозавры встречаются чаще всего, были относительно плоскими[206]. Другая гипотеза — вероятность опрокидывания анкилозавров хищниками. Эта модель также предложена Нопчей[198], и утверждает, что анкилозавры могли быть перевёрнуты большими плотоядными тероподами, чтобы подвергнуть атаке незащищённую брюшную полость[231]. У крупных теропод каждая задняя конечность имела силу в несколько тонн, способная легко перевернуть на спину небольшого анкилозавра. Согласно этой модели, на перевёрнутых скелетах предпочтительно должны быть следы зубов крупных теропод, таких как тираннозавриды, которые легко идентифицировать на ископаемых костях[237]. Анализ костей показал только один экземпляр с признаками хищничества и отсутствие связи между ориентацией туши и активностью плотоядных животных. Кроме того, более мелким тероподам, вероятно, было бы трудно перекатывать большого анкилозавра на спину[231]. Известно несколько случаев наличия защиты брюшной области у анкилозавров. У рода сайхания были обнаружены остеодермы на нижней части грудной клетки[23], также у молодых особей Liaoningosaurus были обнаружены пластины, покрывающие всю брюшную полость[197].

Анкилозавры на протяжении всего периода своего существования демонстрируют разностороннюю эволюцию защитных черт. Одна из основных и наиболее старых гипотез, которая подтверждается гистологическими данными гласит, что наличие своеобразной защиты было результатом отношений хищника и жертвы между тероподами и анкилозаврами. Такая гипотеза предсказывает, что хищническое давление со стороны теропод было движущей силой эволюции защиты анкилозавров. Исследования также показывают, что ранние анкилозавры были недостаточно защищены от хищников того же времени, в то время как позднемеловые анкилозавры были достаточно хорошо защищёными[101]. При попытке понять взаимосвязь защиты анкилозавров и хищников следует также учитывать, как тероподы убивали крупную добычу. В отличие от менее защищённых групп динозавров, таких как зауроподы, добыча в виде рогатых цератопсов и панцирных анкилозавров была совершенно иной. Крупные тероподы не боролись с ними, а пытались нанести жертве раны, рассчитывая на силу своего укуса. Вероятно своеобразная защита как цератопсов, так и анкилозавров, где остеодермы последних идеально подходили для противодействия хищникам, вызвали эволюцию среди динозавров-теропод[101][238]. В меловом периоде наблюдается эволюционирование разных групп теропод. В раннем мелу отмечается появление ютараптора, который представлял собой хищника средних размеров с большим серповидным когтем на втором пальце задних конечностей[239]. К позднему мелу такие тероподы, как тираннозавриды, приобрели огромные размеры[110]. У них был гораздо более сильный укус и их зубы могли напрямую проломить панцирь. Известен образец рода сайхания с отверстием над глазницей от укуса зуба хищника, вероятно тарбозавра. Однако рана зажила, давая возможность предполагать, что панцирь и крепкий череп обеспечивали относительную защиту анкилозаврам от хищных динозавров группы тираннозаврид[240]. Большие размеры, увеличенная сила укуса и хорошее зрение хищников мелового периода, возможно привели к эволюционной конкуренции «хищник—жертва», где у анкилозавров также отмечается увеличение размеров, комбинация костных пластин, защитного окраса и других стратегий визуальной защиты[219].
Остеодермальный панцирь анкилозавров, возникнув как защитное образование от хищников, возможно, расширил свои функции. Так, например, хотя кость и не является наилучшим проводником теплоты, панцирь вторично мог бы служить животному своего рода аппаратом для регулирования перенагревания и переохлаждения тела[19][40]. Считается, что большие анкилозавры испытывали трудности со сбросом потенциально избыточного тепла из-за их коренастого тела, и что остеодермы помогали им увеличить площадь поверхности тела для этой цели, а губчатые кости содержали каналы вен. У поздних анкилозаврид стенки шипов тонкие, более пористые, с большим количеством отверстий для кровеносных сосудов. Количество отверстий, пронизывающих их стенки, явно превышает необходимое для теплоснабжения рогового чехла. Обилие их, возможно, объяснялось усиленным кровоснабжением кожи над остеодермами[19]. Учитывая тот факт, что и шипы стегозавров, и шипы анкилозавров увеличивали поверхность туловища, ряд исследователей интерпретировали наличие шипов стегозавров как часть механизма отдачи или поглощения тепла[241]. Однако это также оспаривается некоторыми исследователями: у близкородственных стегозавров остеодермы явно покрыты рогом, а не толстой кожной тканью, и структура кости не кажется подходящей для быстрого кровообращения. Кроме того, панцирь вполне мог являться полезным щитом от неблагоприятно влияющих на организм изменений окружающей среды, таких, к примеру, как вредное воздействие радиации[242].
Социальное поведение

Размышления о социальном поведении анкилозавров, как и всех животных известных по ископаемым находкам, затруднительны[243]. Традиционно анкилозавров рассматривали как одиночные животные[226], основываясь на применении «модели современной черепахи». В качестве этой гипотезы высказывались несколько основных тезисов. Большая группа животных быстро поедают источник корма окружающей среды и вынуждены идти дальше в поисках новых мест пропитания. Анкилозаврам, будучи медленно движущимися формами, было бы затруднительно это делать[244] и время на удовлетворение потребностей в пище занимал бы значительную часть[20]. Кроме того, их низкий интеллект не способствовал сложному социальному поведению. Таким образом предполагалось, что у каждой отдельной особи анкилозавров была своя территория, которая также должна была быть довольно большой из-за низкой питательной ценности типичной для их мест обитания пищи. Молодняк не получая «родительского ухода» во время роста вынужден был рассыпаться по местности в поисках собственных мест обитания[243].
Однако реальные находки не согласуются с этой моделью. Некоторые места захоронений анкилозавров (Cedarpelta, Gastonia, Struthiosaurus austriacus, Talarurus, Hungarosaurus) содержат две и более особи[226][244]. Было также обнаружено захоронение большой группы молодых особей рода Pinacosaurus, которые предположительно представляли собой путешествующее стадо, погибших одновременно и погребённых песчаной бурей[245]. В общей сложности известно шесть скоплений массовой гибели анкилозавров, указывающих на стадный образ жизни[244]. Это было расценено как убедительное доказательство того, что стадное поведение у анкилозавров всё же присутствовало, или по крайней мере в группах молодых особей до их полного взросления[246]. Согласно современным представлениям, анкилозавры вполне могли ходить группами в поисках мест питания. Однако находки остатков нескольких животных в одном месте не обязательно указывают на групповое поведение. Предполагается, что у членов каждого известного массового захоронения могли быть свои особые факторы, заставляющие их формировать относительно небольшие стада, что указывает на более сложную социальную структуру у анкилозавров, чем считалось ранее[244]. Например остатки многочисленных неполовозрелых особей Pinacosaurus, могут указывать на то, что молодые особи формировали более крупные группы для защиты[246]. Тем не менее большинство находок дошли до наших дней в виде остатков одной особи[244], поэтому большинство исследователей предполагают, что эти животные жили поодиночке или максимум небольшими группами[20][247].
Некоторые особенности телосложения могут указывать на взаимодействие с сородичами. Большие, выступающие вверх шипы в области предплечья (например у рода Sauropelta), вероятно, служили в меньшей степени для защиты, и в большей для демонстрации[27]. Наблюдение полового диморфизма в длине шипа также вписывается в этот контекст[62]. Хвостовая часть анкилозаврид могла также использоваться в споре двух самцов — например за спаривание с самкой[3], либо в возможных кормовых спорах внутри вида[94].
Половой диморфизм
Надёжные признаки полового диморфизма у ископаемых организмов чрезвычайно трудно установить из-за недостаточности ископаемого материала и слабой методологии[248]. Как и в случае с многими другими группами динозавров, половой диморфизм анкилозавров предполагался у многих видов, но количественные данные часто отсутствуют. На сегодняшний день существует очень мало информации о возможном половом диморфизме у анкилозавров, хотя Кеннет Карпентер[62] предположил, что видимые различия как в длине шипов у Panoplosaurus, так и в расположении костных пластин у Edmontonia rugosidens могут представлять гендерные различия. Некоторые нодозавриды заметно выделяются удлинёнными шипами на плечах или около них, и было высказано предположение, что они могут сцепляться с шипами противника во время поединков за доминирование[249].
Роланд Ганглофф также отмечал возможную диморфность в пропорций черепа и рисунках костных пластин Edmontonia, но уточнил, что некоторые изменения могут быть вызваны такими факторами, как старение или даже дробление[168]. Паскаль Годфруа и Аттила Оши не исключали диморфные различия в черепном «орнаменте» у Pinacosaurus[250] и Hungarosaurus[251]. Пол Пенкальски отметил большое разнообразие черепов, относящихся к роду Euoplocephalus и определил это как признак полового диморфизма[29]. Однако, за редкими исключениями, ограниченное количество образцов или неполная сохранность материала черепа затрудняет идентификацию половых различий[251].
Крупнейшие экземпляры анкилозаврид Северной Америки из кампана имеют более мелкие верхние заглазничные шипы и аморфные шейные остеодермы. Половой диморфизм возможно частично объясняют некоторые наблюдаемые различия. Однако маловероятно, чтобы формы большинства остеодерм индивидуума были сексуально диморфными[252].
Наличие хвостовой «булавы» в качестве отличительных особенностей самцов были отвергнуты Уолтером Кумбсом[42]. Однако исследование вида Tianzhenosaurus youngi в 2020 году, в том числе и хвостовой «булавы», внесло существенный вклад в изучение полового диморфизма у анкилозавров. Изучение 4 скелетов данного вида дало возможность предполагать, что характерные особенности черепов и «булавы» на хвосте являются признаками полового диморфизма. У самцов выявлены большие и крепкие верхние заглазничные шипы, которые выступают вверх, а также большая и плоская хвостовая «булава» круглой формы. У самок более тонкие, меньшего размера верхние заглазничные шипы, которые выступают назад, а также небольшая и плоская хвостовая «булава» круглой формы с орнаментом напоминающим скорлупу грецкого ореха на её поверхности. Это исследование является первым полноценным изучением полового диморфизма у анкилозавров[253].
Межвидовое взаимодействие
Специалист по анкилозаврам Виктория Меган Арбор в своей публикации отметила несколько геологических формаций, содержащих большое видовое разнообразие панцирных динозавров. В качестве примера она приводит рода сайхания, тархия и Zaraapelta из формации Баруун-Гоёот и два неописанных рода из формации Нэмэгэт — обитавших в одно и то же время на территории современной Монголии. Арбор объяснила это видовое богатство тем, что анкилозавриды могли быть доминирующими крупными растительноядными животными данного региона, который в остальном состоял из небольших растительноядных или всеядных динозавров, а также мелкие птиц, ящериц и млекопитающих. Она делает вывод, что территория позволяла удовлетворять потребности в пищи по крайней мере для пяти популяций, хотя их соответствующие ниши были неясны. С другой стороны, по мнению палеонтолога, разнообразие анкилозаврид могло быть достигнуто и за счёт полового отбора. Различный черепной «орнамент» может дополнительно подтверждать данную теорию, при которой «украшения» в виде костных щитков на голове могли быть, по крайней мере частично, своего рода межвидовым различием[254]. Сосуществование высокого разнообразия видов как среди анкилозавров, так и среди других растительноядных динозавров, по сравнению с относительно небольшой территорией доступной им, имело место и на позднемеловом островном континенте Ларамидия (западная часть Северной Америки) — в формациях Сидар-Маунтин[англ.] и Дайносор-Парк[англ.][190][254]. Это также вызывает большой интерес палеонтологов. Для объяснения этих фактов выдвигаются различные гипотезы, среди которых наиболее часто упоминаются: разделение пищевых ниш[190] на основании различий в форме морды[76][191], механике челюстей и форме зубов[56], а также деление по высоте кормления[190].
«Полярные» анкилозавры
Летопись окаменелостей мелового периода показывает большое видовое разнообразие фаун полярных динозавров. Ископаемые остатки анкилозавров также известны из нескольких полярных местонахождений как в северном, так и южном полушариях[168][212]. Современные данные убедительно свидетельствуют о том, что многие полярные динозавры (включая и анкилозавров Новой Зеландии) зимовали, а не мигрировали. Считается, что анкилозавры были больше предрасположенные к перезимовке на основе их физической неспособности мигрировать (биомеханика и размеры). Низкое содержание питательных веществ в пище также предполагают более вероятную перезимовку, нежели энергозатратную миграцию[212]. Неуклюжая походка анкилозавров упоминается как основной и явный фактор, исключающий миграцию[255]. Данная модель даёт возможность палеонтологам предполагать, что анкилозавры, несмотря на их относительно большие размеры, должны были зимовать также в Австралии, Антарктиде и на Аляске, как они это делали в Новой Зеландии во времена кампана. Полярная фауна новозеландского микроконтинента конца мелового периода стоит особняком среди остальных полярных находок[212]. Фауна Маунгатанивхи (англ. Maungataniwha) представляет собой изолированную и уникальную фауну, основанную на больших размерах некоторых видов и сосуществование анкилозавров с зауроподами. Новая Зеландия во время кампана лежала в пределах антарктического круга, а движение морского дна 85 млн лет назад начало выталкивать её от антарктического материка[256]. Несмотря на большую широту, влияние относительно тёплых морских течений (средняя годовая температура примерно 14˚С) значительно согревали климат[212]. Молнар и Уиффен предполагают прохладный, влажный климат с относительными зимними холодами и снегом[147]. Палеогеографические реконструкции определяют Новую Зеландию как островную территорию во времена кампана. Если эта интерпретация верна, динозавры данной территории определённо зимовали, исключая возможность их миграции. Это показывает, что по крайней мере некоторые виды анкилозавров были способны зимовать в течение долгой австралийской зимы (примерно 3 месяца), а также могли выживать на низкокалорийном корме. Хотя условия на полюсах в меловом периоде были гораздо более стабильными, чем сегодня, долгая полярная ночь сыграла значительную роль в приспособляемости анкилозавров. Из всего этого следует, что анкилозавры высоких широт были холодоустойчивы[212].
В северном полушарии в конце мелового периода (68—65 миллионов лет назад) местонахождения анкилозавров, обнаруженные в Каканауте на северо-востоке России[187], а также на Аляске[168] показывают, что динозавры всё ещё были сильно разнообразны в арктических регионах незадолго до мел-палеогенового вымирания. Фрагменты анкилозавров, а также других видов динозавров мелового периода указывают на то, что они могли воспроизводиться в полярных регионах и, вероятно, жили в высоких широтах круглый год. Палеоботанические данные предполагают, что полярные анкилозавры Каканаута жили в умеренном климате, средняя годовая температура была около 10˚С[187]. На Аляске исследования указывают о колебания средних годовых температур в меловом периоде в пределах 3˚С—13˚С[103].
Гистология костей рода Antarctopelta указывает на то, что, как и в случае с другими полярными динозаврами Южного полушария, нет явных различий в степени роста по сравнению с его низкоширотными родственниками. Никакие очевидные физиологические изменения не связаны с распространением анкилозавров и других динозавров в высоких широтах (более 60˚ южной широты)[257].
Гипотезы
Ранние исследования североамериканских анкилозавров выявили многочисленные экземпляры, сохранившиеся в перевёрнутом виде[198][258]. После многолетнего коллекционирования динозавров в западной Канаде и США, Чарльз М. Штернберг отметил, что почти все собранные им образцы анкилозавров сохранились в перевёрнутом виде[259]. Наблюдения о частоте перевёрнутых анкилозавров получили широкое обсуждение. Согласно основной гипотезе, после смыва рекой или обрушения береговой части, туши анкилозавров оказываясь в воде, переворачивались вверх ногами, при этом тело держалось в таком положении из-за тяжёлых остеодерм. Вздутие туши из-за скопления газов в результате разложения также способствовали удержанию её в таком положении. Затем трупы либо опускались на дно, после дегазации, либо были отложены на берегу[231]. Кеннет Карпентер предположил, что броненосцы могут служить полезной аналогией для понимания того, почему анкилозавры почти всегда встречаются в перевёрнутом виде. Он утверждал, что после смети броненосцы раздуваются, и этого вздутия достаточно, чтобы тяжёлый панцирь перевернул животное на спину. Он предполагал, что аналогичный процесс происходил и у анкилозавров, у которых с броненосцами общее округлое тело и покрытая костным панцирем спина[206]. Исследование 2018 года показало, что 70 % скелетов анкилозавров из провинции Альберта были обнаружены в перевёрнутом виде. Компьютерное моделирование показало, что набухшие в воде туши имеют тенденцию переворачиваться быстрее всего у нодозаврид[231].

При изучении черепного материала анкилозавров было обнаружено, что у некоторых видов анкилозаврид имеются необычные, достаточно сложные дыхательные пути[13][23]. Эти носовые ходы анкилозаврид легко отличить от нодозаврид по целому ряду анатомических особенностей, в частности по уникальной форме. Как и у большинства динозавров, носовые ходы нодозаврид представляли собой парные трубки, идущие от ноздрей прямо к горлу. Напротив, носовые ходы анкилозаврид (например у эвоплоцефала) следовали по извилистому S-образному пути, и по обе стороны от носовых ходов черепа были пронизаны сложными пазухами. Кроме того, у монгольских анкилозаврид мелового периода (пинакозавр и сайхания) были свёрнутые структуры из тонкой кости в полости носа — турбиналии. Они присутствуют в носовых ходах многих млекопитающих, а также человека. Точное функции сложных носовых ходов, пазух и турбиналий анкилозаврид в настоящее время неизвестны[260][261]. Исследователи приписывают им несколько функций. По мнению некоторых исследователей, сложные полости носа и пазух могли использоваться для охлаждения или увлажнения воздуха у обитателей сухого климата[23][66]. Существуют предположения, что эти полости носа могли служить и для охлаждения мозга или для удаления конденсата при выдохе. Однако эти взгляды поддерживают не все учёные[65], так как эвоплоцефал и многие другие анкилозавриды, у которых были обнаружены такие же полости, обитали в более влажных средах обитания[65][66]. Было предложено множество других объяснений, таких как размещение солевых желез[262], улучшенное обоняние[66] или использование в качестве резонансной полости для генерации всех видов звуковых сигналов[262]. Носовые ходы и пазухи анкилозаврид напоминают те, что были в черепах гадрозавров, поэтому вполне вероятно, что анкилозавриды могли использовать их для создания характерных шумов[260]. Самая последняя идея заключается в том, что они косвенно уменьшают массу, а следовательно, и вес головы[263]. Возможно, сложенные носовые ходы и носовые пазухи выполняли все эти функции одновременно[260]. В настоящее время нет единой гипотезы, объясняющей сложные петлеобразные дыхательные пути[69].
Палеопатология
Большие язвенные ямки на внешней поверхности остеодерм анкилозавров известны достаточно давно. Они были приписаны без подтверждающих доказательств зубами хищника или болезни. Обнаружение многочисленных примеров остеодерм без ямок у моноспецифического анкилозавра (Gastonia) предоставила возможность учёным изучить эту патологию более подробно. Ямки обычно характеризуются углублениями с гладкими стенками, которые разрушают поверхность кости. Предполагается несколько причин этому: бактериальный или грибковый дерматоз; также выдвигается гипотеза о эрозии поверхности кости в результате образования гранулёмы в тонкой коже или на границе раздела кожа-остеодерма[264]. При возможном повреждении остеодерм эктопаразитами, известных в существующих и ископаемых панцирях черепах, это может указывать на вероятное существование паразитов у динозавров[265].
Тазовые и хвостовые элементы анкилозавров были исследованы на предмет признаков заживления ран, которые могли бы указывать на травмы при использовании хвостовой «булавы» в качестве оружия. Патологий позвонков задней части хвоста в местах окостеневших сухожилий не было обнаружено. Если хвостовые «булавы» анкилозаврид и использовались в качестве оружия, то они либо были хорошо приспособлены для сопротивления переломам, либо типичные травмы были слишком критичны и вряд ли зажили. Помимо этого несколько центральных хвостовых позвонков показывают признаки инфекции в нервных корешках и поперечных отростках. Часть окостеневших сухожилий хвоста имеет поражение, которое может представлять инфекцию, а ряд других имеют необычную бороздчатую текстуру указывающую на патологию, но её этиология неизвестна. Также обнаружены большие поражения тазовых костей у двух скелетов на стыке четвёртого спинно-крестцового поперечного отростка и подвздошной кости, их этиология также неизвестна[266].
Следы
Ископаемые следы анкилозавров распространены достаточно широко, включая Азию, Европу, Северную Америку и Южную Америку[65][173], Австралию[267]. Большинство следов сконцентрированы в прибрежных и пойменных отложениях, представлявших во времена динозавров влажные, c обильной растительностью среды обитания[173]. Несмотря на некоторый материал, собранный из засушливых (эоловых) отложений Азии, понятие анкилозавры, как обитателей влажных сред, поддерживается одновременным присутствием многочисленных водных видов позвоночных в местах находок ископаемых[65].
Анкилозаврам были приписаны несколько отдельных ихнотаксонов. Самым ранним ископаемым следом анкилозавров является — Metatetrapous valdensis, известен из берриасских слоёв Вальдена, Германия. Впервые был описан Францем Нопчей в 1923 году, однако последующая потеря исходного материала сделала это описание и статус ихнотаксона сомнительным для многих последующих авторов. Остались только очень краткие оригинальные описания, сопровождаемые одним рисунком[171][173]. В 1932 году К. Штернбергом был описан новый ихнотаксон — Tetrapodosaurus borealis из раннемеловой формации Гетинг, Британская Колумбия, Канада. Первоначально он приписал следы цератопсам[268], но позже они были с уверенностью отнесены к анкилозаврам семейства нодозаврид[173]. Сравнение морфологии стопы рода зауропелта с окаменелыми следами Tetrapodosaurus borealis позволяет предположить, что следы могут относятся к данному роду[206]. К этому же ихнотаксону относят следы из формации Zorrillo-Taberna Indiferenciadas в Мексике, более раннего среднеюрского (байосс) возраста, что делает эти следы одними из самых старых известных на сегодняшний день ихнотаксонов анкилозавров. Данные следы сохранились в виде выпуклых гипорельефов[86]. Следы Macropodosaurus gravis позднего мела (сеноман), найденные на территории Таджикистана, были описаны в 1964 году С. А. Захаровым и состояли из серии четырёхпалых следов около 50 см в длину и 30 см в ширину. Изначально Захаров отнёс следы Macropodosaurus к хищным динозаврам-тероподам, однако Р. Маккри отнёс их к анкилозаврам, основываясь на сходстве отпечатков задних лап в сравнении со следами Metatetrapous[86][173]. В 1984 году на основе следов из формации Эль-Молино в Боливии Джузеппе Леонарди описал ихнотаксон Ligabueichnium bolivianum[175]. Автор предположил возможную их связь с цератопсидом или анкилозавром, однако в последующей работе он упомянул, что «Ligabueichnium bolivianum может быть отнесён к необычно крупному представителю анкилозавров» и в настоящее время рядом авторов он рассматривается как принадлежащий анкилозавру[86]. Также известен ихнотаксон Deltapodus brodericki относящийся к юрским (аален—байосс) следам анкилозавра (или стегозавра) из Йоркшира, Англия[173]. Первоначально они были описаны как следы зауроподов, но позже, в 1994 году, Локли с коллегами отнесли их к возможным следам анкилозавров[149]. При невозможности отнесения ископаемых следов к какому либо инхнотаксону их определяют как принадлежащие неизвестным анкилозаврам — Ankylosauria indet. Найденные на территории Италии следы, относящиеся к нижней юре (синемюр), могут являться самыми ранними свидетельствами анкилозавров[84].
Систематика
Латинское название Ankylosauria было впервые использовано Генри Фэрфилдом Осборном в 1923 году для подотряда динозавров[12]. Эпонимом стал род анкилозавр (Ankylosaurus). Первое определение кладе анкилозавров было дано в 1997 году американским палеонтологом Кеннетом Карпентером как группа, состоящая из всех тиреофор, которые более тесно связаны с родом Ankylosaurus, чем родом Stegosaurus[269]. Этому определению по существу придерживается большинство палеонтологов сегодня. В 1998 году у другого американского палеонтолога Пола Серено было аналогичное определение: все представители клады Eurypoda, которые более тесно связаны с анкилозаврами, чем со стегозаврами[270]. В 2004 году Серено дал более осторожное определение анкилозавров. Оно звучало как группа, включающую все виды, более близкие к Ankylosaurus magniventris, чем к Stegosaurus stenops, Pachycephalosaurus wyomingensis и Parasaurolophus walkeri. Сложное определение отражает множество неопределённостей относительно точных взаимоотношений внутри птицетазовых динозавров. В том же году Джонатан Рихард Вагнер предложил ещё более сложное определение: все виды, более тесно связанные с Ankylosaurus magniventris, чем с Heterodontosaurus foxii, Pachycephalosaurus wyomingensis, Stegosaurus armatus, Iguanodon bernissartensis и Ceratops montanus[271]. Это сделано для того, чтобы граница соответствовала общепринятому представлению об анкилозаврах: массивные, приземистые животные, с множеством костных пластин на теле.
Проблемы систематики
Несмотря на долгую историю исследований, взаимоотношения анкилозавров внутри группы остаются плохо изученными, а существующие кладистические анализы страдают от ограниченности ископаемого материала[105]. В 1978 году американский палеонтолог и специалист по анкилозаврам Уолтер Престон Кумбс заявил, что всех анкилозавров можно строго разделить на две основные группы: нодозавриды и анкилозавриды[13]. Это представление было подвержено сомнению ещё в конце 1990-х годов рядом кладистических исследований, которые показали, что некоторые ранние формы не могут быть отнесены ни к одной из этих групп[105]. Таким образом, в основании генеалогического древа за пределами групп нодозаврид и анкилозаврид оказались более ранние, примитивные анкилозавры. Для этой группы Джеймс Киркланд в своём филогенетическом анализе использовал старый таксон Polacanthidae, впервые предложенный в далёком 1911 году[272]. В 2001 году Дун Чжимин использовал другой таксон — сцелидозавриды (Scelidosauridae), первоначально предложенный Томасом Генри Гексли в 1869 году, куда помимо вновь описанного им рода Bienosaurus включил ряд других примитивных анкилозавров, ссылаясь на их близкое родство[273]. В этом же году Кеннет Карпентер предложил новую кладу Ankylosauromorpha, объединяющую сцелидозавра, анкилозаврид, нодозаврид, полакантид, а также род Minmi, считающимся примитивным анкилозавром[35]. Однако другие систематики исключают данную группу, как и проведённым им филогенетический анализ. Полученное Карпентером филогенетическое дерево не рассматривается, поскольку таксоны были отнесены к кладам до анализа матрицы признаков[274]. Последующий более обширный филогенетический анализ с использованием только черепных признаков не смог проверить достоверность клады Polacanthidae/Polacanthinae из-за отсутствия черепного материала ранних анкилозавров[3].
С тех пор было проведено много кладистических анализов, часто с очень разными результатами. Существует ряд различных подходов в систематике анкилозавров. Уильям Блоу придерживался того же мнения, что и Уолтер Кумбс, считая полакантидид примитивными нодозавридами[275]. Этой же точки зрения придерживается и Питер Гальтон в своей публикации, посвящённой ранним анкилозаврам[276]. В других исследованиях, например у Джеймса Киркланда, было обнаружено, что полакантиды являются базальными членами клады анкилозаврид[272]. Третий подход состоит в том, что полакантиды образуют раннюю боковую ветвь анкилозавров[35]. Эта гипотеза была очень популярна среди некоторых исследователей, но её недостатком является то, что для её сестринских групп, нодозаврид и анкилозаврид, всё ещё существует длинная скрытая линия эволюционного развития. Ещё одна версия, которая не имеет этого недостатка, заключается в том, что полакантиды являются парафилетической группой[141]. Один из последних пересмотренных комплексных анализов анкилозавров указывает на то, что некоторые представители группы полакантид являются базальными как в группе нодозаврид, так в группе анкилозаврид[105]. Это подтверждается самым последним и всеобъемлющим на данный момент кладистическим анализом, опубликованным Викторией Меган Арбор в 2016 году[60]. Наиболее сложная ситуация в систематике анкилозавров заключается в распределении ранних форм и их базальном положений в двух основных семействах. Отсутствие согласия по этому вопросу отчасти связано с необычной морфологией черепа анкилозавров: например черепной «орнамент» закрывает все костные швы взрослых особей[57], что делает практически невозможным точное определение границ черепных костей, тем самым ограничивая межвидовые сравнения. В то же время посткраниальный скелет (часть скелета, за исключением черепа) у анкилозавров консервативен и одинаков почти для всех представителей группы[105].
В 2021 году Дэвид Б. Норман предложили пересмотреть определение анкилозавров, ограничив его двумя семействами — нодозавриды и анкилозавриды, и определив кладу Ankylosauromorpha, предложенную Кеннет Карпентер в далёком 2001 году, для всех таксонов, более близких к анкилозавру, чем к стегозавру[277]. В том же году Даниэль Мадзия с коллегами сочли пересмотр определения анкилозавров нежелательным, предпочтя отказаться от Ankylosauromorpha как младшего синонима Ankylosauria с тем же определением. Неофициальная клада Euankylosauria, предложенная Норманом в качестве альтернативы переопределению Ankylosauria, содержащая семейства нодозаврид и анкилозаврид, обсуждалась ими как потенциально полезная в будущем[278], и была определена другой группой учёных под руководством Серхио Сото-Акунья как таковая в результате их новых филогенетических результатов, которые сгруппировали таксоны Antarctopelta, Kunbarrasaurus (Minmi sp. из более ранних анализов) и Stegouros, как уникальную группу анкилозавров, не относящихся к кладе Euankylosauria, и определили их в новую кладу Parankylosauria[279].
Внешняя систематика
В XXI веке ранжированная классификация в значительной степени потеряла популярность среди палеонтологов динозавров. Вместо привычных отрядов и подотрядов всё чаще стал использоваться термин клада (группа). Анкилозавры являются монофилетической кладой четвероногих растительноядных динозавров, особенностью которых являются костные образования на туловище, а также покрытие черепа остеодермальными щитками[3]. Сестринской группой анкилозаврам являются стегозавры, у которых также на спине были костные пластины, но расположенные вертикально в два ряда. Анкилозавры и стегозавры образуют кладу Eurypoda. Вместе с некоторыми примитивными представителями из нижней юры (например рода Scutellosaurus, Emausaurus и Scelidosaurus) группа Eurypoda образуют кладу тиреофоры (буквально «щитоносцы»)[55][105]. Тиреофоры относятся к группе птицетазовых динозавров (Ornithischia)[105].
Ниже представлена упрощённая кладограмма группы тиреофор, построенная в результате филогенетического анализа Ричардом Томпсоном и его коллегами в 2011 году[105]:
| Тиреофоры |
| ||||||||||||||||||||||||||||||
Внутренняя систематика
Внутренняя систематика анкилозавров противоречива. Традиционно анкилозавры включают в себя два семейства: нодозавриды и анкилозавриды[13][105], но также предложено и третье семейство — Polacanthidae[272][35].
Анкилозавриды характеризуются черепом, резко расширяющимся в области орбит и за ними, ширина его равна или превышает длину; наличием роговых образований на заднебоковых частях черепа; верх черепа покрыт большим количеством мелких остеодермальных щитков[280]; у некоторых более поздних анкилозаврид имеется наличие хвостовой «булавы»[46]. Анкилозаврид можно поделить на две группы — Shamosaurinae и Ankylosaurinae. К первой будут отнесены два рода анкилозаврид — шамозавр, как один из самых древних представителей семейства, и сайхания, обладающая более примитивными чертами строения. Вторая группа включает всех остальных азиатских анкилозавров, древние представители которых, возможно, были исходными для североамериканских эвоплоцефалюса и анкилозавра[97].
Нодозавриды имеют более узкий и вытянутый череп, длина которого значительно превышает ширину; крупные шипы в плечевой области; верх черепа покрыт немногочисленными крупными остеодермальными щитками; отсутствуют роговые образования за орбитами и костный нарост на конце хвоста[280]. Обладали достаточно массивным плечевым поясом. Клювы были более узкие, чем у анкилозаврид — различия, связанные с их избирательностью в отношении растительного корма[69]. Большинство находок нодозаврид происходит из Северной Америки. Среди представителей семейства Уолтер Кумбс наметил несколько ветвей: одна из них была представлена примитивными рода — Hylaeosaurus и Struthiosaurus, характеризующихся небольшими размерами, другая ветвь представлена североамериканскими нодозавридами[13], их ранние представители — Mymoorapelta и Gargoyleosaurus.
Семейство Polacanthidae было введено американским палеонтологом Джорджем Ребером Виландом в 1911 году для обозначения группы анкилозавров, которые казались ему промежуточными между анкилозавридами и нодозавридами[281]. Однако это предложение не было принято во внимание, и было возрождено только в 1998 году Джеймсом Киркландом[272]. Группа объединила черты обеих семейств, и поэтому иногда рассматривается как подсемейство анкило-, а иногда и нодозаврид[105], другие исследования рассматривают её как отдельную группу[40][60] — обычно её определяют как всех анкилозавров, более близких к Gastonia, чем к Edmontonia или Euoplocephalus[35]. Большинство филогенетических анализов не выделяют эту группу в отдельную кладу[3][46][105].
Кладограмма на основе исследований Серхио Сото-Акунья с соавторами, 2021 год[279].
| Thyreophora |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Классификация
По данным сайта Fossilworks, на декабрь 2020 года в инфраотряд включают следующие таксоны[282]:
- Семейство Ankylosauridae Brown, 1908 — Анкилозавриды
- Подсемейство Ankylosaurinae Nopcsa, 1918
- Семейство Nodosauridae Marsh, 1890 — Нодозавриды
- Подсемейство Struthiosaurinae Nopcsa, 1923
- Семейство Polacanthidae Wieland, 1911
- Род Gastonia Kirkland, 1998
- Роды incertae sedis
- Acanthopholis Huxley, 1867
- Aletopelta Ford & Kirkland, 2001
- Amtosaurus Kurzanov & Tumanova, 1978
- Bissektipelta Parish & Barrett, 2004
- Dracopelta Galton, 1980
- Kunbarrasaurus Leahey et al., 2015
- Metatetrapous Ballerstedt, 1922
- Niobrarasaurus Carpenter et al., 1995
- Palaeoscincus Leidy, 1856
- Qijiangpus Xing et al., 2007
- Sarcolestes Lydekker, 1893
- Stegopelta Williston, 1905
- Tianchiasaurus Dong, 1993
Ещё больше десятка родов включают в инфраотряд в статусе nomen dubium: Brachypodosaurus Chakravarti, 1934, Cryptodraco Lydekker, 1889, Cryptosaurus Seeley, 1875, Eucercosaurus Seeley, 1879, Hierosaurus Wieland, 1909, Maleevus Tumanova, 1987, Peishansaurus Bohlin, 1953, Polacanthoides Nopcsa, 1928, Priconodon Marsh, 1888, Priodontognathus Seeley, 1875, Rhadinosaurus Seeley, 1881, Rhodanosaurus Nopcsa, 1929, Sauroplites Bohlin, 1953, Stegosaurides Bohlin, 1953, Syngonosaurus Seeley, 1879[282].
| Хронологическое распределение анкилозавров | |||||||
|---|---|---|---|---|---|---|---|
| Polacanthidae | Ankylosauridae | Nodosauridae | incertae sedis | ||||
| Период | Отдел | Ярус | Возраст | ||||
| Мел | Верхний мел | Маастрихт | 72,1—66,0 | Ankylosaurus Nodocephalosaurus Tarchia | Edmontonia Struthiosaurus | ||
| Кампан | 83,6—72,1 | Euoplocephalus Pinacosaurus Saichania | Aletopelta Antarctopelta* Edmontonia Panoplosaurus Struthiosaurus | Palaeoscincus | |||
| Сантон | 86,3—83,6 | Talarurus Tsagantegia | Hungarosaurus Niobrarasaurus | Niobrarasaurus | |||
| Коньяк | 89,8—86,3 | Bissektipelta | Niobrarasaurus | Amtosaurus | |||
| Турон | 93,9—89,8 | ||||||
| Сеноман | 100,5—93,9 | Zhongyuanosaurus Crichtonsaurus | Animantarx Nodosaurus Silvisaurus Stegopelta | Acanthopholis Amtosaurus Bissektipelta Stegopelta | |||
| Нижний мел | Альб | 113,0—100,5 | Cedarpelta Gobisaurus Shamosaurus | Anoplosaurus Pawpawsaurus Peloroplites Prioconodon Sauropelta Texasetes | Kunbarrasaurus Qijiangpus Stegopelta | ||
| Апт | 125,0—113,0 | Minmi | Sauropelta | Qijiangpus | |||
| Баррем | 129,4—125,0 | Gastonia | Hoplitosaurus Liaoningosaurus Polacanthus | Hylaeosaurus | Qijiangpus | ||
| Готерив | 132,9—129,4 | Gastonia | Hylaeosaurus | ||||
| Валанжин | 139,8—132,9 | Hylaeosaurus | |||||
| Берриас | 145,0—139,8 | Metatetrapous | |||||
| Юра | Верхняя юра | Титон | 152,1—145,0 | Dracopelta Gargoyleosaurus Mymoorapelta | |||
| Киммеридж | 157,3—152,1 | Gargoyleosaurus | |||||
| Оксфорд | 163,5—157,3 | ||||||
| Средняя юра | Келловей | 166,1—163,5 | Sarcolestes Tianchiasaurus | ||||
| Бат | 168,3—166,1 | ||||||
| Байос | 170,3—168,3 | ||||||
| Аален | 174,1—170,3 | ||||||
| Нижняя юра | Тоар | 182,7—174,1 | |||||
| Плинсбах | 190,8—182,7 | ||||||
| Синемюр | 199,3—190,8 | ||||||
| Геттанг | 201,3—199,3 | ||||||
| Деление дано в соответствии с IUGS по состоянию на декабрь 2016 | |||||||
В культуре
В 2020 году Австрия выпустила €3 монету с изображением Ankylosaurus magniventris. На лицевой стороне 3 евро представлен цветной анкилозавр в родной доисторической среде обитания. На заднем плане изобразили хищника, который представлял угрозу для растительноядного существа[283].
Примечания
- Комментарии
- ↑ Передняя часть хвоста образца ROM 75860 рода Zuul в настоящее время закрыта мягкими тканями, поэтому общая длина хвоста может немного превышать 300 см.
- Использованные источники
- ↑ 1 2 Малеев Е. А. 1964. Подотряд Ankylosauria. Анкилозавры. Основы палеонтологии. М.: Наука, Т. 12. С. 574—577.
- ↑ Анкилозавры / А. К. Рождественский // Ангола — Барзас. — М. : Советская энциклопедия, 1970. — (Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров ; 1969—1978, т. 2).
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 Vickaryous, M. K., Maryańska, T., Weishampel, D. B. 2004. Ankylosauria. Pp. 363—392. In D. B. Weishampel, P. Dodson, and H. Osmólska (eds) The Dinosauria, 2nd edition. University of California Press, Berkeley.
- ↑ Paul, 2016
- ↑ 1 2 3 Туманова, 1987, с. 59
- ↑ 1 2 3 4 5 Туманова, 1987, с. 62.
- ↑ Туманова, 1987, с. 71.
- ↑ 1 2 3 Zhiming, D. 1993. An ankylosaur (ornithischian dinosaur) from the Middle Jurassic of the Junggar Basin, China. Vertebrata PalAsiatica, 31: 257—266 [1] Архивная копия от 15 февраля 2021 на Wayback Machine
- ↑ Weishampel et al., 2004, p. 619
- ↑ 1 2 Fossilworks: Ankylosauria. fossilworks.org. Дата обращения: 14 января 2021. Архивировано 9 июня 2020 года.
- ↑ 1 2 3 4 Carpenter, K. and B. Breithaupt. 1986. Latest Cretaceous occurrence of nodosaurid ankylosaurs (Dinosauria, Ornithischia) in western North America and the gradual extinction of the dinosaurs. Journal of Vertebrate Paleontology 6: 251—257.
- ↑ 1 2 3 Obsborn, H. F. 1923. Two Lower Cretaceous dinosaurs of Mongolia. American Museum Novitates, 95: 1—10
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 Coombs, W. P. 1978. The families of the ornithischian dinosaur order Ankylosauria. Journal of Paleontology 21: 143—170.
- ↑ ankylosaurs - Encyclopedia of Life. eol.org. Дата обращения: 14 января 2021. Архивировано 20 ноября 2020 года.
- ↑ Currie and Padian, 1997, p. 162
- ↑ 1 2 3 Carpenter, 2004, p. 961
- ↑ 1 2 Ben Creisler Dinosauria Translation and Pronunciation Guide.
- ↑ 1 2 3 Brown, B. 1908. The Ankylosauridae, a new family of armored dinosaurs from the Upper Cretaceous. American Museum of Natural History Bulletin 24: 187—201.
- ↑ 1 2 3 4 Туманова, 1987, с. 60
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Victoria M. Arbour, Jordan C. Mallon. Unusual cranial and postcranial anatomy in the archetypal ankylosaur Ankylosaurus magniventris (англ.) // FACETS 2. — 2017. — P. 764–794. — doi:10.1139/facets-2017-0063.
- ↑ 1 2 Paul, 2016, p. 258.
- ↑ Paul, 2016, р. 265
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 Maryanska, T. 1977. Ankylosauridae (Dinosauria) from Mongolia. Palaeontologia Polonica, 37: 85—151. [2] Архивная копия от 31 августа 2021 на Wayback Machine
- ↑ 1 2 3 4 Туманова, 1987, с. 61
- ↑ 1 2 3 4 Carpenter, 2012, рp. 513—514
- ↑ 1 2 3 Xu X., Wang X.-L., and You H.-L. 2001. A juvenile ankylosaur from China. Naturwissenschaften, 88 (7): 297—300
- ↑ 1 2 3 4 Carpenter, 2012, р. 521
- ↑ 1 2 3 Carpenter, 2012, р. 505.
- ↑ 1 2 Penkalski, P. 2001. Variation in specimens referred to Euoplocephalus tutus. Pp. 261—298. In: K. Carpenter (ed.). The armored dinosaurs. Bloomington: Indiana University Press.
- ↑ Ankylosaurus- The Bullet-Proof Dinosaur ¦ Newsgrape.com. web.archive.org (10 мая 2012). Дата обращения: 7 января 2021. Архивировано из оригинала 10 мая 2012 года.
- ↑ Малеев, Е. А. 1956. Панцирные динозавры верхнего мела Монголии (Семейство Анкилозавриды). Труды Палеонтологического Института Академии Наук СССР, 62: 51—91.
- ↑ Weishampel et al., 2004, p. 517—606.
- ↑ 1 2 3 4 5 6 Туманова, 1987, с. 49—50
- ↑ 1 2 3 Carpenter, 2012, pр. 515—516
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 Carpenter, K. 2001. Phylogenetic analysis of the Ankylosauria. Pр. 455—482. In Carpenter, Kenneth (eds). The Armored Dinosaurs. Indiana University Press.
- ↑ Gaston, R. W., Schellenbach, J. and Kirkland, J. I. 2001. Mounted Skeleton of the Polacanthine Ankylosaur Gastonia burgei. Pp. 386—398. In K. Carpenter (ed.). The Armored Dinosaurs. Indiana University Press, Bloomington.
- ↑ 1 2 3 Brown, C.M., et al. 2017. An exceptionally preserved armored dinosaur reveals the morphology andallometry of osteoderms and their horny epidermal coverings. PeerJ, 5 (3)
- ↑ Carpenter, 2012, р. 515
- ↑ Scheyer, T. M. and Sander, P. M. 2004. Histology of ankylosaur osteoderms: implications for systematics and function. Journal of Vertebrate Paleontology 24: 874—894.
- ↑ 1 2 3 4 Hayashi, S., Carpenter, K., Scheyer, T. M., Watabe, M. and Suzuki. D. 2010. Function and evolution of ankylosaur dermal armor. Acta Palaeontologica Polonica, 55 (2): 213—228.
- ↑ Parks, W.A. 1924. Dyoplosaurus acutosquameus: a new genus and species of armoured dinosaur with notes on a skeleton of Prosaurolophus maximus. Univ. of Toronto Studies (Geological Series), 18: 1-35
- ↑ 1 2 3 4 5 Coombs, W. P. 1995. Ankylosaurian tail clubs of middle Campanian to early Maastrichtian age from western North America, with description of a tiny club from Alberta and discussion of tail orientation and tail club function. Canadian Journal of Earth Sciences 32: 902—912.
- ↑ 1 2 Carpenter, 2012, р. 516
- ↑ 1 2 Arbour, V. M., Zanno, L. E. 2018. The evolution of tail weaponization in amniotes. Proceedings of the Royal Society B: Biological Sciences 285
- ↑ 1 2 Arbour, V. M. and Currie, P. J. 2013. Euoplocephalus tutus and the diversity of ankylosaurid dinosaurs in the Late Cretaceous of Alberta, Canada, and Montana, USA. PLoS ONE, 8: e62421
- ↑ 1 2 3 4 5 6 7 8 9 Han, F., Zheng, W., Hu, D., Xu, X., Barrett, P. M. 2014. A new basal ankylosaurid (Dinosauria: Ornithischia) from the Lower Cretaceous Jiufotang Formation of Liaoning Province, China. PLoS ONE 9 (8)
- ↑ 1 2 3 Zheng, W., Jin, X., Azuma, Y., Wang, Q., Miyata, K. & Xu, X. 2018. The most basal ankylosaurine dinosaur from the Albian-Cenomanian of China, with implications for the evolution of the tail club. Scientific Reports 8 (1): 3711
- ↑ 1 2 3 4 5 Blows, W. T. 2013. Notice of nodosaur (Dinosauria, Ankylosauria) remains from the mid-Cretaceous of Cambridge, England, with comments on cervical half-ring armour. Proceedings of the Geologists Association 125 (1): 56—62.
- ↑ Ostrom, J. H. 1970. Stratigraphy and paleontology of the Cloverly Formation (Lower Cretaceous) of the Bighorn Basin and Wyoming and Montana. Bull. Peabody Mus. Natur. Hist. Jale Univ., 35: 1—152.
- ↑ 1 2 3 Туманова, 1987, с. 25
- ↑ Weishampel et al., 2004, pp. 388—390
- ↑ Weishampel et al., 2004, p. 390
- ↑ Eaton, T. 1960. A new armoured dinosaur from the Creataceous of Kansas. Univ. Kans. Paleontol. Contrib., 8: 1—24.
- ↑ 1 2 Maryańska, T. 1971. New data on the skull of Pinacosaurus grangeri (Ankylosauria). Palaeontologia Polonica 25: 45—53.
- ↑ 1 2 3 4 Sereno, P. C. 1986. Phylogeny of the bird-hipped dinosaurs (Order Ornithischia). National Geographic Research, 2: 234—256
- ↑ 1 2 3 4 5 Carpenter, K. 1997. Ankylosaurs. Pp. 308—316. In J. Farlow and M. Brett-Surman (eds.). The Complete Dinosaur. Indiana University Press, Bloomington.
- ↑ 1 2 3 Vickaryous, M. K., Russell, A. P. and Currie, P. J. 2001. Cranial ornamentation of ankylosaurs (Ornithischia: Thyreophora): Reappraisal of developmental hypotheses. Pp. 318—340. In: K. Carpenter (ed.). The Armored Dinosaurs. Indiana University Press, Bloomington.
- ↑ 1 2 Carpenter, K., Kirkland, J. I., Burge, D. and Bird, J. 2001. Disarticulated skull of a new primitive ankylosaurid from the Lower Cretaceous of Eastern Utah. Pp. 211—328. In K. Carpenter (ed.). The Armored Dinosaurs. Indiana University Press, Bloomington.
- ↑ 1 2 Carpenter, 2012, р. 507
- ↑ 1 2 3 4 5 6 7 8 9 10 11 Arbour, V. M., Currie, P. J. 2016. Systematics, phylogeny and palaeobiogeography of the ankylosaurid dinosaurs. Journal of Systematic Palaeontology 14 (5): 385—444.
- ↑ 1 2 Coombs, W. P. 1972. The bony eyelid of Euoplocephalus (Reptilia: Ornithischia). Journal of Paleontology 46: 637—650.
- ↑ 1 2 3 4 Carpenter, K. 1990. Ankylosaur systematics: example using Panoplosaurus and Edmontonia (Ankylosauria, Nodosauridae). Pp. 282—298. In K. Carpenter and P. Currie (eds.). Dinosaur Systematics: Perspectives and Approaches. Cambridge University Press, New York.
- ↑ 1 2 Туманова, 1987
- ↑ Туманова, 1987, с. 26
- ↑ 1 2 3 4 5 6 7 Weishampel et al., 2004, p. 392
- ↑ 1 2 3 4 5 Carpenter, 2012, р. 508
- ↑ Туманова, 1987, с. 31
- ↑ 1 2 3 4 Туманова, 1987, с. 12
- ↑ 1 2 3 4 5 6 Carpenter, 2012, р. 509
- ↑ 1 2 Туманова, 1987, с. 53
- ↑ Carpenter, 2012, р. 510
- ↑ Paul, 2016, р. 26
- ↑ Туманова, 1987, с. 43
- ↑ Carpenter, 2012, рp. 511—512
- ↑ 1 2 3 4 5 Ankylosauria. tolweb.org. Дата обращения: 5 января 2021. Архивировано 27 февраля 2021 года.
- ↑ 1 2 3 Carpenter, K. 1982. Skeletal and dermal armor reconstruction of Euoplocephalus tutus (Ornithischia: Ankylosauridae) from the Late Cretaceous Oldman Formation of Alberta. Canadian Journal of Earth Sciences, 19: 689—697.
- ↑ 1 2 3 Coombs, W. P. 1978. Forelimb muscles of the Ankylosauria (Reptilia: Ornithischia). Journal of Paleontology 52: 642—658.
- ↑ Norman, D. B., Witmer, L. M. and Weishampel, D. B. 2004. Basal Thyreophora. Pр. 335—342. In D. B. Weishampel, P. Dodson, and H. Osmolska (eds.), The Dinosauria. Berkeley: University of California Press.
- ↑ 1 2 Туманова, 1987, с. 52
- ↑ PBDB. paleobiodb.org. Дата обращения: 7 января 2021. Архивировано 9 января 2021 года.
- ↑ Clark, N.D.L. 2001. A thyreophoran dinosaur from the early Bajocian (Middle Jurassic) of the Isle of Skye, Scotland. Scottish Journal of Geology, 37: 19—26.
- ↑ 1 2 Туманова, 1987, с. 55
- ↑ Sereno, P. C. 1999. The evolution of dinosaurs. Science, 284: 2137—2146
- ↑ 1 2 Avanzini, M., Leonardi, G., Tomasoni R. and Campolongo M. 2001. Enigmatic dinosaur trackways from the Lower Jurassic (Pliensbachian) of the Sarca Valley, northeast Italy. Ichnos 8: 235—242
- ↑ Carpenter, K., DiCroсe, T., Gilpin, D., Kinneer, B., Sanders, F., TidWell, V. & Shaw, A. 2002. Origins of the Early and «Middle» Cretaceous dinosaurs of North America: implications for plate tectonics. Proceedings of the International Symposium on New Concepts in Global Tectonics: 289—308.
- ↑ 1 2 3 4 Rosa, R. A., Velasco, P., Lozano-Carmona, D.-E., Arellano, J. 2018. Middle Jurassic ankylosaur tracks from Mexico. Boletín de la Sociedad Geológica Mexicana 70 (2): 379—395. [3]
- ↑ Carpenter, 2012, р. 518
- ↑ 1 2 3 Galton, P. M. 1980. Armored dinosaurs (Ornithischia: Ankylosauria) from the Middle and Upper Jurassic of England. Géobios, 13: 825—837.
- ↑ Carpenter, K., Miles, C. and Cloward, K. 1998. Skull of a Jurassic ankylosaur (Dinosauria). Nature 393: 782—783
- ↑ 1 2 Pereda-Suberbiola, X., Diaz-Martinez, I., Salgado, L. and de Valais, S. 2015. Sintesis del registro fosil de dinosaurios tireoforos en Gondwana. Ameghiniana, 15 (1): 90—107.
- ↑ Weishampel et al., 2004, p. 626
- ↑ Paul, 2016, р. 19—21
- ↑ 1 2 Wyenberg-Henzler, T. 2022. Ecomorphospace occupation of large herbivorous dinosaurs from Late Jurassic through to Late Cretaceous time in North America. PeerJ 10:e13174
- ↑ 1 2 Paul, 2016, р. 22
- ↑ 1 2 Carpenter, K. and Kirkland, J. I. 1998. Review of Lower and Middle Cretaceous ankylosaurs from North America. Pp. 249—270. In S.G. Lucas, J.I. Kirkland and J.W. Estep (eds.). Lower and Middle Cretaceous Terrestrial Ecosystems. New Mexico Museum of Natural History and Science Bulletin No. 14.
- ↑ Pereda-Suberbiola, X. 1992. A revised census of European Late Cretaceous nodosaurids (Ornithischia: Ankylosauria): last occurrence and possible extinction scenarios. Terra Nova 4: 641—648
- ↑ 1 2 Туманова, 1987, с. 54
- ↑ Carpenter, 2012, р. 519
- ↑ Britt, B. B., Eberth, D. A., Scheetz, R. D., Greenhalgh, B. W., and Stadtman, K. L. 2009. Taphonomy of debris-flow hosted dinosaur bonebeds at Dalton Wells, Utah (Lower Cretaceous, Cedar Mountain Formation, USA), Palaeogeogr. Palaeocl., 280: 1—22.
- ↑ 1 2 3 Несов, 1995, с. 21
- ↑ 1 2 3 Freer, C. 2017. Dinosaur Warfare: Ankylosaur and Theropod Coevolution. Biology. [4] Архивная копия от 29 марта 2020 на Wayback Machine
- ↑ Fossilworks: Liaoningosaurus. fossilworks.org. Дата обращения: 14 января 2021. Архивировано 4 июля 2020 года.
- ↑ 1 2 3 Fiorillo, A. R. 2006. Review of the dinosaur record of Alaska with comments regarding Korean dinosaurs as comparable high-latitude fossil faunas. Journal of Paleontological Society of Korea 22 (1): 15—27.
- ↑ Arbour, V. M. & Currie, P. J. 2015. Ankylosaurid dinosaur tail clubs evolved through stepwise acquisition of key features. Journal of Anatomy 227 (4): 514—523
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 Thompson, R. S., Parrish, J. C., Maidment, S., Barrett,P. M. 2011. Phylogeny of the ankylosaurian dinosaurs (Ornithischia: Thyreophora). Journal of Systematic Palaeontology 10 (2): 301—312
- ↑ McDonald, A.T. and Wolfe, D. G. 2018. A new nodosaurid ankylosaur (Dinosauria:Thyreophora) from the Upper Cretaceous Menefee Formation of New Mexico. PeerJ, 6 (2)
- ↑ 1 2 3 Arbour V. M., Zanno L. E., Gates T. 2016. Ankylosaurian dinosaur palaeoenvironmental associations were influenced by extirpation, sea-level fluctuation, and geodispersal. Palaeogeography, Palaeoclimatology, Palaeoecology 449: 289—299
- ↑ 1 2 Gasparini, Z., Pereda-Suberbiola, X. and Molnar, R. 1996. New data on the ankylosaurian dinosaurs from the Late Cretaceous of the Antarctic Peninsula. Memoirs of the Queensland Museum 39: 583—594.
- ↑ Coria, R. A. and Salgado, L. 2001. South American Ankylosaurs. Pp. 159—168. In K. Carpenter (ed.). The Armored Dinosaurs. Indiana University Press, Bloomington.
- ↑ 1 2 Paul, 2016, р. 23
- ↑ Smith et al., 1994
- ↑ 1 2 Несов, 1995, с. 75
- ↑ Hildebrand, A. R. et al. 1991. Chicxulub Crater: a possible Cretaceous/Tertiary boundary impact crater on the Yucatan Peninsula, Mexico. Geology 19: 867—871.
- ↑ Ankylosauridae. tolweb.org. Дата обращения: 13 января 2021. Архивировано 10 апреля 2021 года.
- ↑ Mantell, G. 1833. The Geology of the South-East of England. London: Longman, Rees, Orme, Brown, Green, & Longman, pp. 289—313
- ↑ Owen, R. 1842. Report on British fossil reptiles. Part II. Reports of the British Association for the Advancement of Science, 11: 60—204.
- ↑ Anonymous (1865) A new Wealden dragon. Order, Sauria; Family, Dinosaurian; genus, Polacanthus; Species, foxi . The Illustrated London News 47: 270
- ↑ Fox, W. 1866. On a new Wealden saurian named Polacanthus. Report of the British Association for the Advancement of Science, Birmingham 1865: 56
- ↑ Huxley, T. H. 1867. On Acanthopholis horridus, a new reptile from the Chalk-marl. Geological Magazine 4: 65—67.
- ↑ Owen, R. 1861. A monograph of a fossil dinosaur (Scelidosaurus harrisonii, Owen) of the Lower Lias, part I. Monographs on the British fossil Reptilia from the Oolitic Formations 1: 1—14.
- ↑ Madzia, D., Boyd, C. A. and Mazuch, M. 2018. A basal ornithopod dinosaur from the Cenomanian of the Czech Republic. Journal of Systematic Palaeontology 16 (11): 967—979.
- ↑ Holmes, Thom. 1998. Fossil Feud: The Rivalry of the First American Dinosaur Hunters. Parsippany, NJ: Julian Messner.
- ↑ Bunzel, E. 1871. Die Reptilfauna der Gosaformation in der Neuen Welt bei Wiener-Neustadt. Abhandlungen der Kaiserlich-Königlichen Geologischen Reichsanstalt. 5: 1—18.
- ↑ Seeley, H. G. 1879. On the Dinosauria of the Cambridge Greensand. Quarterly Journal of the Geological Society of London 35: 591—635.
- ↑ Seeley, H. G. 1875. On the maxillary bone of a new dinosaur (Priodontognathus phillipsii), contained in the Woodwardian Museum of the University of Cambridge. Quarterly Journal of the Geological Society of London 31: 439—443.
- ↑ Seeley, H. G. 1875. On the femur of Cryptosaurus eumerus, Seeley, a dinosaur from the Oxford Clay of Great Gransden. Quarterly Journal of the Geological Society of London 319: 149—151.
- ↑ Lydekker, R. 1893. On the jaw of a new carnivorous dinosaur from the Oxford Clay of Peterborough. Quarterly Journal of the Geological Society of London 49: 284—287.
- ↑ Marsh, O. C. 1890. Additional characters of the Ceratopsidae, with notice of new Cretaceous dinosaurs. American Journal of Science 39: 418—426
- ↑ 1 2 Lambe, L. M. 1902. New genera and species from the Belly River series. Contribs Canad. Paleontol., 3 (2): 25—81.
- ↑ Marsh, O. C. 1895. On the affinities and classification of the dinosaurian reptiles. American Journal of Science, 50 (300): 483—498
- ↑ 1 2 Marsh, O. C. 1896. The dinosaurs of North America. In: United States Geological Survey 16th annual report 1894-95, 55: 133—244
- ↑ Nopcsa, F. 1923. Notes on British dinosaurs. Part VI. Acanthopholis. Geol. Mag., 60: 193—199.
- ↑ Romer, A. S. 1927. The pelvic musculature of the ornithischian dinosaurs. Acta zool., 8 (2/3): 225—275.
- ↑ Nopcsa, F. 1929. Dinosaurierreste aus Siebenbttrgen. Geol. hung. Ser. paleontol., 1 (4): 1—76.
- ↑ Gilmore, C. W. 1930. On dinosaurian reptiles from the Two Medicine formation of Montana. Proceedings of the United States National Museum 77 (16): 1—39.
- ↑ Romer, A. S. 1956. Osteology of the reptiles. Chicago: Univ. Chicago press, p. 1—772.
- ↑ Maryanska, T. and Osmólska, H. 1985. On ornithischian phylogeny. Acta Palaeontologica Polonica, 30 (3—4): 137—149.
- ↑ Coombs, W. P. 1971, The Ankylosauridae. Ph.D. thesis, Columbia University, New York, NY, p. 487.
- ↑ 1 2 Coombs, W. P., Maryanska, T. 1990. Ankylosauria. Pp. 456—483. In D. B. Weishampel. P. Dodson & H. Osmólska (eds). The Dinosauria. 1st Edition. University of California Press, Berkeley.
- ↑ Vickaryous, M. K., Russell, A. P., Currie, P. J., Zhao, X.-J. 2001. A new ankylosaurid (Dinosauria: Ankylosauria) from the Lower Cretaceous of China, with comments on ankylosaurian relationships. Canadian Journal of Earth Sciences, 38: 1767—1780.
- ↑ 1 2 3 4 Hill, R. V., Witmer, L. M., Norell, M. A. 2003. A new specimen of Pinacosaurus grangeri (Dinosauria: Ornithischia) from the Late Cretaceous of Mongolia: ontogeny and phylogeny of ankylosaurs. American Museum Novitates, 3395: 1—29.
- ↑ Berkey, C., Morris, F. K. 1927. Geology of Mongolia. Natural History of Central Asia 2: 1—475.
- ↑ 1 2 Gilmore, C.W. 1933. Two new dinosaurian reptiles from Mongolia with notes on some fragmentary specimens. American Museum Novitates 679: 1—20.
- ↑ 1 2 3 4 5 Туманова, 1987, с. 63
- ↑ Soto-Acuña S., Vargas A., Kaluza J., et al. 2021. Bizarre tail weaponry in a transitional ankylosaur from subantarctic Chile. Nature 600: 259—263.
- ↑ Maidment, S. C. R., Strachan, S. J., Ouarhache, D., Scheyer, T. M., Brown, E. E., Fernandez, V.; Johanson, Z., Raven, T. J., Barrett, P. M. 2021. Bizarre dermal armour suggests the first African ankylosaur. Nature Ecology & Evolution: 1-6.
- ↑ 1 2 3 Molnar, R. E. and Wiffen, J. 1994. A Late Cretaceous polar dinosaur fauna from New Zealand. Cretaceous Research 15: 689—706.
- ↑ Russell, D. A., Russell, D. E., Taquet, P. & Thomas, H. 1976. Nouvelles récoltes de Vertébrés dans les terrains continentaux du Crétacé supérieur de la région de Majunga (Madagascar). Compte Rendu Sommaire de la Société Géologique de France, 5: 205—208
- ↑ 1 2 3 4 5 6 7 Osi, A. 2015. The European ankylosaur record: a review. Hantkeniana, 10: 89—106.
- ↑ Galton, P. M. 1983. Armored dinosaurs (Ornithischia: Ankylosauria) from the Middle and Upper Jurassic of Europe. Palaeontographica, Abteilung A 182: 1—25.
- ↑ Pereda-Suberbiola, X. and Barret, P. M. 1999. A systematic review of ankylosaurian dinosaur remains from the Albian-Cenomanian of England. Special Papers in Palaeontology, 60: 177—208
- ↑ 1 2 3 4 Fastovsky et al., 2005, p. 134
- ↑ Averianov, A. O. 2002. An ankylosaurid (Ornithischia: Ankylosauria) braincase from the Upper Cretaceous Bissekty formation of Uzbekistan. Bull. l’Institut R. des Sci. Nat. Belqique, Sci. la Terre. 72: 97—110.
- ↑ 1 2 Galton, P. M. 2019. Earliest record of an ankylosaurian dinosaur (Ornithischia Thyreophora): dermal armour from Lower Kota Formation (Lower Jurassic) of India. Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen 291 (2): 205—219.
- ↑ Hayakawa, H. & Carpenter, K. 1996. First occurrence of nodosauridankylosaurs in Asia. Palaeontological Society of Japan 71: 98
- ↑ Augustin, F. J., Matzke, A., Pfretzschner, H.-U. 2020. First evidence of an ankylosaur (Dinosauria, Ornithischia) from the Jurassic Qigu Formation (Junggar Basin, NW China) and the early fossil record of Ankylosauria. Geobios 61: 1—10
- ↑ Шувалов В. Ф. 1974. О геологическом строении и возрасте местонахождений Хобур и Хурэи-Дух // Фауна и биостратиграфия мезозоя Монголии. М.: Наука, с. 296—304.
- ↑ 1 2 Туманова, 1987, с. 67
- ↑ Малеев Е. А. 1954. Панцирные динозавры верхнего мела Монголии // Тр. ПИН. Т. 48. С. 142—170.
- ↑ Туманова, 1987, с. 10
- ↑ Туманова, 1987, с. 68
- ↑ Туманова, 1987, с. 69—70
- ↑ Coombs, W. P. 1979. Osteology and myology of the hindlimb in the Ankylosauria (Reptilia, Ornithischia). Journal of Paleontology 53: 666—684.
- ↑ Kirkland, J. and Carpenter, K. 1994. North America’s first pre-Cretaceous ankylosaur (Dinosauria) from the Upper Jurassic Morrison Formation of Western Colorado. Brigham Young University Geology Studies, 40: 25—42.
- ↑ Penkalski, P. 2018. Revised systematics of the armoured dinosaur Euoplocephalus and its allies. Neues Jahrbuch für Geologie und Paläontologie — Abhandlungen 287 (3): 261—306.
- ↑ 1 2 Victoria M. Arbour, David C. Evans. A new ankylosaurine dinosaur from the Judith River Formation of Montana, USA, based on an exceptional skeleton with soft tissue preservation // Royal Society Open Science. — Т. 4, вып. 5. — С. 161086. — doi:10.1098/rsos.161086. Архивировано 11 ноября 2020 года.
- ↑ 1 2 Wiersma, J. P. & Irmis, R. B. 2018. A new southern Laramidian ankylosaurid, Akainacephalus johnsoni gen. et sp. nov., from the upper Campanian Kaiparowits Formation of southern Utah, USA. PeerJ 6: e5016.
- ↑ 1 2 3 4 Gangloff, R. A. 1995. Edmontonia sp. the first record of an ankylosaur from Alaska. Journal of Vertebrate Paleontology 15 (1): 195—200.
- ↑ 1 2 Туманова, Т. А., Алифанов В. Р., Болотский Ю. Л. 2003. В России впервые обнаружены остатки анкилозавров. Природа (3): 69—70
- ↑ Victoria M. Arbour, Michael E. Burns, Robert M. Sullivan, Spencer G. Lucas, Amanda K. Cantrell. A New Ankylosaurid Dinosaur from the Upper Cretaceous (Kirtlandian) of New Mexico with Implications for Ankylosaurid Diversity in the Upper Cretaceous of Western North America (англ.) // PLOS ONE. — 2014-09-24. — Vol. 9, iss. 9. — P. e108804. — ISSN 1932-6203. — doi:10.1371/journal.pone.0108804. Архивировано 5 марта 2021 года.
- ↑ 1 2 Francischini, H., Sales, M., Schultz,C. L. 2017. The Presence of Ankylosaur Tracks in the Guará Formation (Brazil) and Remarks on the Spatial and Temporal Distribution of Late Jurassic Dinosaurs. Ichnos 25 (3).
- ↑ Murray, A., Riguetti, F., Rozadilla, S. 2019. New ankylosaur (Thyreophora, ornithischia) remains from the Upper Cretaceous of Patagonia. Journal of South American Earth Sciences 96.
- ↑ 1 2 3 4 5 6 7 8 McCrea, R. T., Lockley, M. G. & Meyer, C. A. 2001. Global distribution of purported ankylosaur track occurrences. In K. Carpenter (Ed.). The armored dinosaurs. Pp. 413—454. Bloomington: Indiana University Press.
- ↑ 1 2 Знание-сила, 2004 № 10 (928)
- ↑ 1 2 Leonardi, G. 1984. Le impronte fossili di Dinosauri. Sulle Orme dei Dinosauri, 164—186.
- ↑ Meyer, Ch. A. et al. 2020. The Late Cretaceous dinosaur track record of Bolivia — Review and perspective. Journal of South American Earth Sciences, 106
- ↑ Salgado, L., Coria, R. A. 1996. First evidence of an ankylosaur (Dinosauria, Ornithischia) in South America. Ameghiniana 33 (4): 367—371.
- ↑ Gasparini et al. 2015. Late Cretaceous reptilian biota of the La Colonia Formation, central Patagonia, Argentina: occurrences, preservation and paleoenvironments. Cretaceous Research 54: 154—168.
- ↑ Leonardi, G. and Carvalho, I. d. S. 2002. Icnofósseis da Bacia do Rio do Peixe, PB: o mais marcante registro de pegadas de dinossauros do Brasil. C. Schobbenhaus, D. d. A. Campos, E. T. Queiroz, M. Winge, & M. Berbert-Born (eds.), Sítios Geológicos e Paleontológicos do Brasil. SIGEP, Brasilia 101—111.
- ↑ Leahey, L. G., Salisbury, S. W. 2013. First evidence of ankylosaurian dinosaurs (Ornithischia: Thyreophora) from the mid-Cretaceous (late Albian-Cenomanian) Winton Formation of Queensland, Australia. An Australasian Journal of Palaeontology 37: 249—257
- ↑ Long, J. A. et al. 2002. Dinosaurs of Australia and New Zealand and Other Animals of the Mesozoic Era. New South Wales University Press, Sydney; 188 pp.
- ↑ Wiffen, J. 1996. Dinosaurian palaeobiology: a New Zealand perspective. Memoirs of the Queensland Museum 39: 725—731.
- ↑ 1 2 Lamanna, M., Case, J., Roberts, E., Arbour, V. M. 2019. Late Cretaceous non-avian dinosaurs from the James Ross Basin, Antarctica: description of new material, updated synthesis, biostratigraphy, and paleobiogeography. Advances in Polar Science 30 (3): 228—250.
- ↑ Gasparini, Z., Olivero, E., Scasso, R. & Rinaldi, C. 1987. Un ankilosaurio (Reptilia, Ornithischia) Campaniano en el continente antártico. Anais do X Congresso Brasileiro de Paleontologia, Río de Janeiro: 131—141.
- ↑ Архангельский, М. С., Иванов, А. В., Нелихов, А. Е. 2012. Когда Волга была морем. — Саратов: Сарат. гос. техн. ун-т. — 56 с. http://herba.msu.ru/shipunov/school/books/arkhangelsky2012_kogda_volga_byla_morem.pdf Архивная копия от 27 августа 2018 на Wayback Machine
- ↑ Рождественский А. К. 1957. О месторождении верхнемеловых динозавров на р. Амур. Vertebrata Palasiatica 1 (4): 285—291
- ↑ 1 2 3 Godefroit, P., Golovneva, L., Shchepetov, S., Garcia, G. and Alekseev, P. 2009. The last polar dinosaurs: high diversity of latest Cretaceous arctic dinosaurs in Russia. Naturwissenschaften 96: 495—501.
- ↑ Matthews, W. 1922. A super-dreadnaught of theanimal world: the armored dinosaur Palaeoscincus. Natural History, 22: 333—342.
- ↑ 1 2 Haas, G. 1969. On the jaw musculature of Ankylosaurus. American Museum Novitates, 2399: 1—11.
- ↑ 1 2 3 4 Mallon, J. C., Evans, D. C., Ryan, M. J., Anderson, J. S. 2013. Feeding height stratification among the herbivorous dinosaurs from the Dinosaur Park Formation (upper Campanian) of Alberta, Canada. BMC Ecology 13: 14
- ↑ 1 2 Carpenter, 2004, p. 985
- ↑ Fastvosky et al., 2005, p. 135
- ↑ 1 2 Fastovsky et al., 2005, p. 137
- ↑ 1 2 Paul, 2016, р. 21
- ↑ Mallon, J. C. and Anderson, J. S. 2014. The functional and palaeoecological implications of tooth morphology and wear for the megaherbivorous dinosaurs from the Dinosaur Park Formation (upper Campanian) of Alberta, Canada. PLoS ONE, 9: e98605.
- ↑ Hill, R. V., D’Emic, M. D., Bever, G. S., Norell, M. A. 2015. A complex hyobranchial apparatus in a Cretaceous dinosaur and the antiquity of avian paraglossalia. Zoological Journal of the Linnean Society 175 (4): 892—909
- ↑ 1 2 Ji, Q., Wu, X., Cheng, Y., Ten, F., Wang, X. and Ji, Y. 2016. Fish hunting ankylosaurs (Dinosauria, Ornithischia) from the Cretaceous of China. Journal of Geology 40: 183—190.
- ↑ 1 2 3 4 Nopcsa, F. 1928. Palaeontological notes on reptiles. Geologica Hungerica, Seria Palaeontologica, 1: 1—84.
- ↑ 1 2 3 Brown, C. M., et al. 2020. Dietary palaeoecology of an Early Cretaceous armoured dinosaur (Ornithischia; Nodosauridae) based on floral analysis of stomach contents. Royal Society Open Science 7 (6): 200305.
- ↑ 1 2 Molnar, R. E. and Clifford, H. T. 2000. Gut contents of a small ankylosaur. Journal of Vertebrate Paleontology 20 (1): 194—196.
- ↑ 1 2 Molnar, R. E. and Clifford, T. H. 2001. An ankylosaurian cololite from Queensland, Australia. Pp. 399—412. In Kenneth Carpenter (ed.). The Armored Dinosaurs. Indiana University Press.
- ↑ 1 2 3 Fastovsky et al., 2005, pp. 135—136
- ↑ Mallon J. C. 2019. Competition structured a Late Cretaceous megaherbivorous dinosaur assemblage. Scientific Reports 9: 1—18.
- ↑ Rybczynski, N. and Vickaryous, M. K. 2001. Evidence of Complex Jaw Movement in the Late Cretaceous Ankylosaurid, Euoplocephalus tutus (Dinosauria: Thyreophora). In K. Carpenter (ed.). The Armored Dinosaurs. Indiana University Press, Bloomington.
- ↑ Ősi, A., Prondvai, E., Mallon, J. & Bodor, E. R. 2016. Diversity and convergences in the evolution of feeding adaptations in ankylosaurs (Dinosauria: Ornithischia). Historical Biology 29 (4): 539—570.
- ↑ 1 2 3 4 Carpenter, K. 1984. Skeletal reconstruction and life restoration of Sauropelta (Ankylosauria: Nodosauridae) from the Cretaceous of North America. Canadian Journal of Earth Sciences, 21, 1491—1498.
- ↑ 1 2 Coombs, W. P. 1978. Theoretical aspects of cursorial adaptations in dinosaurs. Quarterly Review of Biology, 53: 393—418.
- ↑ Thulborn, R. A. 1982. Speeds and gaits of dinosaurs. Palaeogeography, Palaeoclimatology, Palaeoecology, 38: 227—256.
- ↑ McCrea, R. T. 2000. Vertebrate Palaeoichnology of the Lower Cretaceous (Lower Albian) Gates Formation of Alberta. Saskatoon, University of Saskatchewan, 133 p.
- ↑ Fastovsky and Weishampel, 2009, p. 265—266
- ↑ 1 2 Paul, 2016, р. 49
- ↑ 1 2 3 4 5 6 Bell, P. R. and Snively, E. 2008. Polar dinosaurs on parade: a review of dinosaur migration. Alcheringa An Australasian Journal of Palaeontology 32 (3): 271—284.
- ↑ 1 2 Paulina-Carabajal, A. et al. 2017. Neuroanatomy of the ankylosaurid dinosaurs Tarchia teresae and Talarurus plicatospineus from the Upper Cretaceous of Mongolia, with comments on endocranial variability among ankylosaurs. Palaeogeography, Palaeoclimatology, Palaeoecology 494: 135—146.
- ↑ Hopson, J. A. 1980. Relative brain size in dinosaurs -implications for dinosaurian endothermy. Pp. 287—370. In Thomas, R. D. K. and Olson, E. C. (eds.). A CoId Look qt the Warm-Blooded Dinosaurs. Westview Press, Boulder, CO.
- ↑ Buchholtz, E. 2012. Dinosaur Paleoneurology. Pp. 191—208. In Brett-Surman, M.К., Farlow, J. O. and Holtz T. R. (eds.). The Complete Dinosaur. Edition: 2nd. Indiana University Press, Bloomington.
- ↑ 1 2 Ivan Kuzmin, Ivan Petrov, Alexander Averianov, Elizaveta Boitsova, Pavel Skutschas, Hans-Dieter Sues. The braincase of Bissektipelta archibaldi — new insights into endocranial osteology, vasculature, and paleoneurobiology of ankylosaurian dinosaurs (англ.). Biological Communications (5 июня 2020). Дата обращения: 7 января 2021. Архивировано 31 января 2019 года.
- ↑ Arbour, V. M., Burns, M. E., Bell, P. R., and Currie, P. J. 2014. Epidermal and dermal integumentary structures of ankylosaurian dinosaurs. Journal of Morphology, 275(1): 39—50.
- ↑ Gilmore, C. W. 1914. Osteology of the armored Dinosauria in the United States National museum: with special reference to the genus Stegosaurus. United States National Museum Bulletin 89: 1—143
- ↑ 1 2 Brown, C. M. et al. 2017. An exceptionally preserved three-dimensional armored dinosaur reveals insights into coloration and Cretaceous predator-prey dynamics. Current Biology 27: 2514—2521
- ↑ Currie and Padian, 1997, p. 206
- ↑ Sellés, A. G., Galobart, À. 2015. Reassessing the endemic European Upper Cretaceous dinosaur egg Cairanoolithus. Historical Biology, 28 (5): 583—596
- ↑ Tanke, D., Brett-Surman, M. 2001. Evidence of hatchling and nesting-size hadrosaur (Reptilia: Ornithischia) from Dinosaur Provincial Park (Dinosaur Park Formation: Campanian), Alberta. In: Tanke, D. H. & Carpenter, K. (ed.). Mesozoic Vertebrate Life. Indiana University Press, pp. 206—218
- ↑ Stanford, R., Weishampel, D., DeLeon V. 2011. The First Hatchling Dinosaur Reported from the Eastern United States: Propanoplosaurus marylandicus (Dinosauria: Ankylosauria) from the Early Cretaceous of Maryland, U.S.A. Journal of Paleontology 85 (5): 916—924
- ↑ Stein, M., Hayashi, S., Sander, P. M. 2013. Long Bone Histology and Growth Patterns in Ankylosaurs: Implications for Life History and Evolution. PLOS ONE 8 (7)
- ↑ This Is the Best Dinosaur Fossil of Its Kind Ever Found (англ.). Magazine (12 мая 2017). Дата обращения: 9 января 2021. Архивировано 3 января 2021 года.
- ↑ 1 2 3 Weishampel et al., 2004, p. 391
- ↑ Hutchinson, J. R., Garcia, M. 2002. Tyrannosaurus was not a fast runner. Nature 415 (6875): 1018—1021
- ↑ Fastovsky et al., 2005, р. 139.
- ↑ Thulborn, T. 1993. Mimicry in ankylosaurid dinosaurs. Record ofthe South Australian Museum 27: 151—158
- ↑ Carpenter, 2004, p. 965
- ↑ 1 2 3 4 5 Mallon, J. C., Henderson, D. M., McDonough, C. M. and Loughry, W. J. 2018. A «bloat-and-float» taphonomic model best explains the upside-down preservation of ankylosaurs". Palaeogeography, Palaeoclimatology, Palaeoecology, 497: 117—127.
- ↑ Carpenter, 2012, р. 522
- ↑ Нейш и Барретт, 2018
- ↑ Баррет и Сане, 2005, с. 68.
- ↑ Palmer, 1999
- ↑ Delmas, V., Baudry, E., Girondot, M., Prevot-Julliard, A.-C. 2007. The righting response as a fitness index in freshwater turtles. Biological Journal of the Linnean Society 91 (1): 99—109
- ↑ Jacobsen, A. R. 1998. Feeding behaviour of carnivorous dinosaurs as determined by tooth marks on dinosaur bones. Historical Biology 13: 17—26
- ↑ Hurum, J. H., and Sabath, K. 2003. Giant theropod dinosaurs from Asia and North America: Skulls of Tarbosaurus bataar and Tyrannosaurus rex compared. Acta Palaeontologica Polonica 48 (2): 161—190.
- ↑ Kirkland, J. I., Burge, D., Gaston, R. 1993. A large dromaeosaurid (Theropoda) from the Lower Cretaceous of Eastern Utah. Hunteria 2 (10): 1—16
- ↑ Gallagher, W. B., Tumanova, T. A., Dodson, P., Axel, L. 1998. CT scanning Asian ankylosaurs: paleopathology in a Tarchia skull. Journal of Vertebrate Paleontology 18: 44A—45A
- ↑ Farlow, J., Thompson, C., Rosner, D. 1976. Plates of the dinosaur Stegosaurus: forced convection heat loss fins? Science 192: 1123—1125.
- ↑ Туманова, 1987, с. 51
- ↑ 1 2 Isles, T. 2009. The socio-sexual behaviour of extant archosaurs: Implications for understanding dinosaur behaviour. Historical Biology 21 (Nos. 3—4): 139—214
- ↑ 1 2 3 4 5 Botfalvai, G., Prondvai, E. and Osi, A. 2020. Living alone or moving in herds? A holistic approach highlights complexity in the social lifestyle of Cretaceous ankylosaurs. Cretaceous Research 118
- ↑ Currie, P. J. 1989. Long distance dinosaurs. Natural History 6: 60—65.
- ↑ 1 2 Burns, M. E., Currie P. J., Sissons R. L. and Arbour V. M. 2011. Juvenile specimens of Pinacosaurus grangeri Gilmore, 1933 (Ornithischia: Ankylosauria) from the Late Cretaceous of China, with comments on the specific taxonomy of Pinacosaurus. Cretaceous Research 32: 174—186
- ↑ Fastovsky et al., 2005, p. 135.
- ↑ Mallon, J. C. 2017. Recognizing sexual dimorphism in the fossil record: lessons from nonavian dinosaurs. Paleobiology 43 (3): 1—13.
- ↑ Coombs, W. P. 1990. Behaviour patterns in dinosaurs. Pp. 32—42. In: Wieshampel, D. B., Dodson, P., Osmolska H. (eds.). The dinosauria. Berkeley (CA): University of California Press.
- ↑ Godefroit, P., Pereda Suberbiola, X., Li, H., Dong, Z-M. 1999. A new species of the ankylosaurid dinosaur Pinacosaurus from the Late Cretaceous of Inner Mongolia (P.R. China). Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre 69: 17—36.
- ↑ 1 2 Åsi, A., Magyar, J., Rosta, K. & Vickaryous, M. 2021. Cranial ornamentation in the Late Cretaceous nodosaurid ankylosaur Hungarosaurus. PeerJ 9:e11010
- ↑ Penkalski, P. & Blows, W. T. 2013. Scolosaurus cutleri (Ornithischia: Ankylosauria) from the Upper Cretaceous Dinosaur Park Formation of Alberta, Canada. Canadian Journal of Earth Sciences, 50 (2):171—182.
- ↑ Duiping, M. 2019. Preliminary study on the sexual dimorphism of Tianzhenosaurus youngi. Journal of Geology 43 (4): 589—594
- ↑ 1 2 Arbour, V. M., Currie, P. J., Badamgarav, D. 2014. The ankylosaurid dinosaurs of the Upper Cretaceous Baruungoyot and Nemegt formations of Mongolia. Zoological Journal of the Linnean Society. 172: 631—652.
- ↑ Paul, G. S. 1997. Dinosaur models: the good, the bad, and using them to estimate the mass of dinosaurs. Pp. 129—142. In D. L. Wolberg, E. Stump & G. Rosenberg (eds.). Dinofest International: Proceedings of a symposium sponsored by Arizona State University. Philadelphia: Academy of Natural Sciences.
- ↑ Davy, B. 2006. Bollons Seamount and early New Zealand-Antarctic seafloor spreading. Geochemistry, Geophysics, Geosystems 7 (6): 1—18.
- ↑ Cerda, I. A. et al. 2019. Paleobiological inferences for the Antarctic dinosaur Antarctopelta oliveroi (Ornithischia: Ankylosauria) based on bone histology of the holotype. Cretaceous Research 103: 104—171.
- ↑ Sternberg, C. M. 1933. Relationships and habitat of Troödon and the nodosaurs. Annals and Magazine of Natural History 11 (62): 231—235
- ↑ Sternberg, C. M. 1970. Comments on dinosaurian preservation in the Cretaceous of Alberta and Wyoming. National Museums of Canada Publications in Palaeontology 4: 1—9
- ↑ 1 2 3 Lucas, 2016, р. 142
- ↑ Bourke, J. M., Porter, W. R., Witmer, L. M. 2018. Convoluted nasal passages function as efficient heat exchangers in ankylosaurs (Dinosauria: Ornithischia: Thyreophora). PLoS ONE 13 (12): e0207381
- ↑ 1 2 Maryanska, 1977, p. 117
- ↑ Witmer, L. M. and Ridgely, R. C. 2008. The paranasal air sinuses of predatory and armored dinosaurs (Archosauria: Theropoda and Ankylosauria) and their contribution to cephalic structure. Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology 291: 1362—1388
- ↑ Matthias, A., McWhinney, L., Carpenter, K. 2016. Pathological pitting in ankylosaur (Dinosauria) osteoderms. International Journal of Paleopathology 13: 82—90
- ↑ Currie and Padian, 1997, p. 528
- ↑ Arbour, V. M., Currie, P. J., 2011. Tail and pelvis pathologies of ankylosaurian dinosaurs. Historical Biology 23, 375—390.
- ↑ Salisbury, S., Romilio, A., Herne, M. C., Tucker, R. T. & Nair, J. P. 2017. The dinosaurian ichnofauna of the Lower Certaceous (Valanginian-Barremian) Broome Sandstone of the Walmadany Area (James Price point), Dampier Peninsula, Western Australia. Journal of Vertebrate Paleontology 36 (1): 1—152.
- ↑ Sternberg, C. M. 1932. Dinosaur tracks from Peace River, British Columbia. National Museum of Canada, Annual Report, 1930: 59—85.
- ↑ Carpenter, K. 1997. Ankylosauria. Pp. 16—20. In P. Currie and K. Padian (eds.). The Encyclopedia of Dinosaurs. Academic Press, San Diego.
- ↑ Sereno, P. C. 1998. A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria. Neues Jahrbuch für Geologie und Paläontologie — Abhandlungen 210 (1): 41—83
- ↑ Wagner, J. 2004. The phylogenetic nomenclature of ornithischian dinosaurs (Vertebrata: Reptilia) and a consideration of the difficulties in converting the names of extinct clades. First International Phylogenetic Nomenclature Meeting At: Paris, France
- ↑ 1 2 3 4 Kirkland, J. I. 1998. A polacanthine ankylosaur (Ornithischia: Dinosauria) from the Early Cretaceous (Barremian) of eastern Utah. New Mexico Museum of Natural History and Science, Bulletin, 14: 271—281.
- ↑ Zhiming, D. 2001. Primitive Armored Dinosaur from the Lufeng Basin, China. In: Tanke, D. H. & Carpenter, K. (ed.). Mesozoic Vertebrate Life. Indiana University Press, pp. 237—243
- ↑ Arbour, V. M. 2014. Systematics, evolution, and biogeography of the ankylosaurid dinosaurs. University of Alberta, 1—418.
- ↑ Blows, W.T. 1996. A new species of Polacanthus (Ornthischia; Ankylosauria) from the Lower Cretaceous of Sussex, England. Geological Magazine, 133: 671—681.
- ↑ Galton, P. M. 2019. Earliest record of an ankylosaurian dinosaur (Ornithischia: Thyreophora): Dermal armor from Lower Kota Formation (Lower Jurassic) of India. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 291 (2): 205—219.
- ↑ Norman, D.B. 2021. Scelidosaurus harrisonii (Dinosauria: Ornithischia) from the Early Jurassic of Dorset, England: biology and phylogenetic relationships. Zoological Journal of the Linnean Society. 191 (1): 1–86.
- ↑ Madzia, D., Arbour, V.M., Boyd, C.A., Farke, A.A., Cruzado-Caballero, P., Evans, D.C. 2021. The phylogenetic nomenclature of ornithischian dinosaurs. PeerJ. 9: e12362.
- ↑ 1 2 Soto-Acuña, Sergio et al. 2021. Bizarre tail weaponry in a transitional ankylosaur from subantarctic Chile. Nature. 600 (7888): 259–263.
- ↑ 1 2 Туманова, 1987, с. 14
- ↑ Wieland, G. R. 1911. Notes on the armored Dinosauria. The American Journal of Science 4 (31): 112—124
- ↑ 1 2 †Ankylosauria (англ.) информация на сайте Fossilworks. (Дата обращения: 15 октября 2017).
- ↑ Ankylosaurus magniventris 2020 3 Euro coin (англ.). www.muenzeoesterreich.at. Дата обращения: 7 января 2021. Архивировано 9 января 2021 года.
Литература
- Carpenter K. Redescription of Ankylosaurus magniventris Brown 1908 (Ankylosauridae) from the Upper Cretaceous of the Western Interior of North America (англ.) // Canadian Journal of Earth Sciences. — 2004. — Iss. 41. — P. 961—986.
- Carpenter K. Ankylosaurs (англ.) // J. Farlow and M. Brett-Surman. The Complete Dinosaur. Edition: 2nd. — Bloomington: Indiana University Press, 2012. — P. 505—525.
- Currie P. J. and Padian K. Encyclopedia of Dinosaurs (англ.). — San Diego: Academic Press, 1997. — 869 p.
- Fastovsky D. E. and Weishampel D. B. The Evolution and Extinction of the Dinosaurs (англ.). — New York: Cambridge University Press, 2005. — 485 p.
- Fastovsky D. E. and Weishampel D. B. Dinosaurs: A Concise Natural History (англ.). — New York: Cambridge University Press, 2009. — 395 p.
- Lucas S. G. Dinosaurs. The Textbook. (англ.). — Columbia University Press, 2016. — 392 p.
- Maryanska T. Ankylosauridae (Dinosauria) from Mongolia (англ.) // Palaeontologia Polonica. — 1977. — Iss. 37. — P. 85—151.
- Palmer D. The Marshall Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals (англ.). — London: Marshall Editions, 1999. — 159 p.
- Paul G. S. The Princeton Field Guide to Dinosaurs (2 ed.) (англ.). — Princeton University Press, 2016. — 360 p.
- Smith A. G., Smith D. G. and Funnell B. M. Atlas of Mesozoic and Cenozoic coastlines (англ.). — Cambridge: Cambridge University Press, 1994. — 99 p.
- Weishampel D. B., Dodson P., Osmólska H., eds. The Dinosauria[англ.] (англ.). — 2nd ed. — Berkeley: University of California Press, 2004. — 880 p.
- Баррет П., Сане Х. Л. Большая энциклопедия динозавров/Dinozaurs - A Natural History. — Москва: Издательский дом «ОНИКС 21 век», 2005. — 192 с.
- Нейш Д., Барретт П. Динозавры. 150 000 000 лет господства на Земле/Dinosaurs: How They Lived and Evolved. — Москва: Альпина нон-фикшн, 2018. — 223 с.
- Несов, Лев Александрович. Динозавры Северной Евразии: новые данные о составе комплексов, экологии и палеогеографии. — Санкт-Петербург: Научно-исследовательский институт земной коры, 1995. — 156 с.
- Туманова Т. А. Панцирные динозавры Монголии. Совместная Советско-Монгольская палеонтологическая экспедиция // Наука. — 1987. — Вып. 37. — С. 1—85.
| Таксономия |
|---|