Археи
| Археи | |
|---|---|
 Halobacteria, штамм NRC-1, каждая клетка длиной около 5 мкм | |
| Научная классификация | |
Домен: Археи | |
| Международное научное название | |
| Archaea Woese, Kandler[англ.] and Wheelis[англ.] 1990 | |
| Синонимы | |
| Типы[2] | |
| |
Архе́и[3] (в единственном числе — архе́я, от лат. Archaea, от др.-греч. ἀρχαῖος «извечный, древний, первозданный, старый») — домен живых организмов (по трёхдоменной системе Карла Вёзе наряду с бактериями и эукариотами). Археи представляют собой одноклеточные микроорганизмы, не имеющие ядра, а также каких-либо мембранных органелл.
Ранее археи объединяли с бактериями в общую группу, называемую прокариоты (или царство Дробянки (лат. Monera)), и они назывались архебактерии, однако сейчас такая классификация считается устаревшей[4]: установлено, что археи имеют свою независимую эволюционную историю и характеризуются многими биохимическими особенностями, отличающими их от других форм жизни.
Сейчас археи подразделяют на более чем 7 типов. Из них наиболее изучены кренархеоты (Crenarchaeota) и эвриархеоты (Euryarchaeota). Классифицировать археи по-прежнему сложно, так как подавляющее большинство из них никогда не выращивались в лабораторных условиях и идентифицировались только по анализу нуклеиновых кислот из проб, полученных из мест их обитания.
Археи и бактерии очень похожи по размеру и форме клеток, хотя некоторые археи имеют довольно необычную форму, например, клетки Haloquadratum walsbyi плоские и квадратные. Несмотря на внешнее сходство с бактериями, некоторые гены и метаболические пути архей сближают их с эукариотами (в частности ферменты, катализирующие процессы транскрипции и трансляции). Другие аспекты биохимии архей уникальны, к примеру, присутствие в клеточных мембранах липидов, содержащих простую эфирную связь. Большая часть архей — хемоавтотрофы. Они используют значительно больше источников энергии, чем эукариоты: начиная от обыкновенных органических соединений, таких как сахара, и заканчивая аммиаком, ионами металлов и даже водородом. Солеустойчивые археи — галоархеи (Haloarchaea) — используют в качестве источника энергии солнечный свет, другие виды архей фиксируют углерод, однако, в отличие от растений и цианобактерий (синезелёных водорослей), ни один вид архей не делает и то, и другое одновременно. Размножение у архей бесполое: бинарное деление, фрагментация и почкование. В отличие от бактерий и эукариот, ни один известный вид архей не формирует спор.
Изначально архей считали экстремофилами, живущими в суровых условиях — горячих источниках, солёных озёрах, однако потом их нашли и в более привычных местах, включая почву, океаны, болота и толстую кишку человека. Архей особенно много в океанах, и, возможно, планктонные археи — самая многочисленная группа ныне живущих организмов. Археи признаны важной составляющей жизни на Земле. Они играют роль в круговоротах углерода и азота. Ни один из известных представителей архей не является ни паразитом (за исключением наноархеот, являющихся паразитами других архей), ни патогенным организмом, однако они часто бывают мутуалистами и комменсалами. Некоторые представители являются метаногенами и обитают в пищеварительном тракте человека и жвачных, где они помогают осуществлять пищеварение. Метаногены используются в производстве биогаза и при очистке канализационных сточных вод, а ферменты экстремофильных микроорганизмов, сохраняющие активность при высоких температурах и в контакте с органическими растворителями, находят своё применение в биотехнологии.
История открытия

Первые представители группы обнаружены в различных экстремальных средах обитания[5], например, геотермальных источниках.
На протяжении большей части XX века прокариоты считались единой группой и классифицировались по биохимическим, морфологическим и метаболическим особенностям. К примеру, микробиологи пытались классифицировать микроорганизмы в зависимости от формы клеток, деталей строения клеточной стенки и потребляемых микроорганизмами веществ[6]. В 1965 году было предложено устанавливать степень родства разных прокариот на основании сходства строения их генов[7]. Этот подход, филогенетика, в наши дни является основным.
Впервые археи были выделены в качестве отдельной группы прокариот на филогенетическом древе в 1977 году Карлом Вёзе и Джорджем Эдвардом Фоксом при сравнительном анализе 16S рРНК. Изначально эти две группы обозначались как архебактерии (лат. Archaebacteria) и эубактерии (лат. Eubacteria) и рассматривались как царства или подцарства, которые Вёзе и Фокс называли термином Urkingdoms. Вёзе настаивал, что эта группа прокариот есть фундаментально отличный тип жизни. Чтобы подчеркнуть это отличие, впоследствии две группы прокариот были названы археями и бактериями[8]. В трёхдоменной системе Карла Вёзе обе эти группы и эукариоты были возведены в ранг домена. Этот термин был предложен Вёзе в 1990 году[9] для обозначения самого верхнего ранга в классификации организмов, включающей одно или несколько царств.
В первое время к новому домену причисляли только метаногенные микроорганизмы. Считалось, что археи населяют только места с экстремальными условиями: горячие источники, солёные озёра. Однако к концу XX века микробиологи пришли к выводу, что археи — большая и разнообразная группа организмов, широко распространённая в природе. Многие виды архей населяют вполне обычные среды обитания, например, почвы или воды океана[10]. Такая переоценка была вызвана применением метода полимеразной цепной реакции для идентификации прокариот в образцах воды и почвы по их нуклеиновым кислотам. Данный метод позволяет выявлять и идентифицировать организмы, которые по тем или иным причинам не культивируются в лабораторных условиях[11][12].
Происхождение и эволюция
Хотя возможные окаменелости прокариотических клеток датированы возрастом в 3,5 млрд лет, большинство прокариот не имеет характерных морфологических особенностей, и поэтому окаменелые формы нельзя определённо идентифицировать именно как останки архей[13]. В то же время химические остатки уникальных для архей липидов более информативны, так как эти соединения у других организмов не встречаются[14]. В некоторых публикациях указывается, что останки липидов архей или эукариот присутствуют в породах возрастом 2,7 млрд лет[15], однако достоверность этих данных остаётся под сомнением[16]. Эти липиды обнаружены в докембрийских формациях. Древнейшие из подобных остатков найдены в Исуанском зеленокаменном поясе на западе Гренландии, где находятся самые старые на Земле осадочные породы, сформировавшиеся 3,8 млрд лет назад[17]. Археи могут быть древнейшими живыми существами, населяющими Землю[18].
Вёзе утверждал, что археи, бактерии и эукариоты представляют собой три раздельные линии, рано отделившиеся от общей предковой группы организмов[19][20]. Возможно, это произошло ещё до клеточной эволюции, когда отсутствие типичной клеточной мембраны давало возможности к неограниченному горизонтальному переносу генов, и предки трёх доменов различались между собой по фиксируемым комплектам генов[20][21]. Не исключено, что последний общий предок архей и бактерий был термофилом, это даёт основания предположить, что низкие температуры были «экстремальной средой» для архей, и организмы, приспособившиеся к ним, появились только позже[22]. Сейчас археи и бактерии связаны между собой не больше, чем с эукариотами, и термин «прокариоты» обозначает лишь «не эукариоты», что ограничивает его применимость[23].
Сравнительная характеристика архей и других доменов
В приведённой таблице показаны некоторые черты архей, свойственные и не свойственные другим доменам[24]. Многие из этих свойств также обсуждаются ниже.
| Свойственно археям и бактериям | Свойственно археям и эукариотам | Свойственно только археям |
|---|---|---|
| Нет оформленного ядра и мембранных органелл | Нет пептидогликана (муреина) | Структура клеточной стенки (к примеру, клеточные стенки некоторых архей содержат псевдомуреин) |
| Кольцевая хромосома | ДНК связана с гистонами[25][26] | В клеточной мембране присутствуют липиды, содержащие простую эфирную связь |
| Гены объединены в опероны | Трансляция начинается с метионина[27] | Структура флагеллинов[28] |
| Схожие РНК-полимераза, промоторы и другие компоненты транскрипционного комплекса, есть интроны и процессинг РНК[28][29][30] | Структура рибосом (некоторые признаки сближают с бактериями, некоторые — с эукариотами) | |
| Полицистронная мРНК | Схожие репликация и репарация ДНК[31] | Структура и метаболизм тРНК[28][32] |
| Размер клеток на несколько порядков меньше, чем у эукариот | Схожая АТФаза (тип V) |
Родство с другими прокариотами
Установление степени родства между тремя доменами имеет ключевое значение для понимания возникновения жизни. Большинство метаболических путей, в которых задействована большая часть генов организма, схожи у бактерий и архей, в то время как гены, отвечающие за экспрессию других генов, очень похожи у архей и эукариот[33]. По строению клеток археи наиболее близки к грамположительным бактериям: клетка покрыта единственной плазматической мембраной, дополнительная внешняя мембрана, характерная для грамотрицательных бактерий, отсутствует[34], клеточные стенки различного химического состава, как правило, толстые[35]. В филогенетическом древе, основанном на сравнительном анализе структур гомологичных генов/белков прокариот, гомологи архей наиболее близки к таковым грамположительных бактерий[34]. В некоторых важнейших белках архей и грамположительных бактерий, таких как Hsp70 и глутаминовая синтетаза I, обнаруживаются одинаковые эволюционно консервативные вставки и делеции[34][36][37].
Гупта (англ. Gupta) предположил, что археи отделились от грамположительных бактерий в результате отбора по признаку устойчивости к действию антибиотиков[34][36][38]. Это основывается на наблюдении, что археи устойчивы к огромному числу антибиотиков, в основном производимых грамположительными бактериями[34][36], и что эти антибиотики действуют главным образом на гены, которые отличают бактерий от архей. Согласно гипотезе Гупты, давление отбора в направлении формирования устойчивости к антибиотикам грамположительных бактерий в конце концов привело к существенным изменениям в структуре генов-мишеней антибиотиков у некоторых микроорганизмов, которые стали общими предками современных архей[38]. Предполагаемая эволюция архей под действием антибиотиков и других неблагоприятных факторов также может объяснить их адаптацию к экстремальным условиям, таким как повышенные температура и кислотность, как результат поиска ниш, свободных от продуцирующих антибиотики организмов[38][39]. Т. Кавалир-Смит выдвинул схожее предположение[40]. Версия Гупты также подтверждается другими работами, изучающими родственные черты в структурах белков[41], и исследованиями, показавшими, что грамположительные бактерии могли быть первой ветвью, отделившейся от общего древа прокариот[42].
Родство с эукариотами
Эволюционное родство между археями и эукариотами остаётся неясным. Помимо сходства в структуре и функциях клеток, между ними существует сходство на генетическом уровне. Установлено, что группа архей кренархеоты стоят ближе к эукариотам, чем к другому типу архей — эвриархеотам[43]. Кроме того, у некоторых бактерий, как Thermotoga maritima обнаружены архееподобные гены, переданные путём горизонтального переноса[44]. Наиболее распространена гипотеза, согласно которой предок эукариот рано отделился от архей[45][46], а эукариоты возникли в результате слияния археи и эубактерии, ставших цитоплазмой и ядром новой клетки. Эта гипотеза объясняет различные генетические сходства, но сталкивается с трудностями в объяснении клеточной структуры[47].
Строение
Форма клеток и колоний
Отдельные клетки архей достигают от 0,1 до 15 мкм в диаметре и могут иметь различную форму: шара, палочки, спирали или диска[48]. Некоторые кренархеоты имеют другую форму, например, Sulfolobus — неправильной дольчатой формы; Thermofilum — тонкой нитевидной формы и меньше 1 мкм в диаметре, а Thermoproteus и Pyrobaculum почти идеально прямоугольные[49]. Haloquadratum walsbyi — плоские квадратные археи, живущие в сверхсолёных водоёмах[50]. Такие необычные формы клеток, вероятно, обеспечиваются клеточной стенкой и прокариотическим цитоскелетом. У архей обнаружены белки, родственные компонентам цитоскелета других организмов[51], а также показано присутствие филаментов в их клетках[52], однако у архей, в отличие от других организмов, эти структуры плохо изучены[53]. У Thermoplasma и Ferroplasma клеточная стенка отсутствует, поэтому их клетки имеют неправильную форму и похожи на амёб[54].
Клетки некоторых видов архей могут объединяться в агрегаты и филаменты длиной до 200 мкм. Эти организмы могут формировать биоплёнки[55]. В культурах Thermococcus coalescens клетки сливаются друг с другом, формируя одну крупную клетку[56]. Археи рода Pyrodictium образуют сложные многоклеточные колонии, в которых клетки объединены с помощью длинных, тонких, полых трубок, называемых cannulae, которые выступают над поверхностями клеток и собирают их в густое кустовидное скопление[57]. Функции этих трубок не ясны, но, возможно, они осуществляют коммуникацию и обмен питательными веществами между соседними клетками[58]. Существуют и многовидовые колонии, как, например, «нить жемчуга», обнаруженная в 2001 году в болоте в Германии. Круглые беловатые колонии некоторых необычных эвриархеот перемежаются тонкими нитями, которые могут достигать до 15 см в длину и состоят из особых видов бактерий[59].
Археи и бактерии имеют очень похожую структуру клеток, однако их состав и организация отделяют архей от бактерий. Как у бактерий, у них отсутствуют внутренние мембраны и органеллы[23], клеточные мембраны, как правило, ограничены клеточной стенкой, а плавание осуществляется за счёт одного или более жгутиков[60]. Структурно археи наиболее схожи с грамположительными бактериями. Большинство имеет одну плазматическую мембрану и клеточную стенку, периплазматическое пространство отсутствует. Исключением из этого главного правила является Ignicoccus, у которого крупное периплазматическое пространство, ограниченное наружной мембраной, содержит окружённые мембраной везикулы[61].
Мембраны
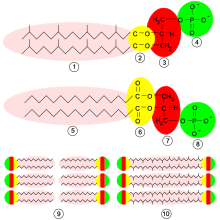
Молекулы, из которых построены мембраны архей, сильно отличаются от тех, которые используются в мембранах других организмов. Это указывает на то, что археи состоят лишь в отдалённом родстве с бактериями и эукариотами[62]. У всех живых организмов клеточные мембраны построены из фосфолипидов. Молекулы фосфолипидов состоят из двух частей: гидрофильной полярной, состоящей из фосфатов, и гидрофобной неполярной, состоящей из липидов. Эти компоненты объединены через остаток глицерина. В воде молекулы фосфолипидов кластеризуются, при этом фосфатные «головки» оказываются обращёнными к воде, а липидные «хвосты» — обращёнными от неё и спрятанными внутрь кластера. Главная составляющая мембраны — два слоя таких фосфолипидов, называемые липидным бислоем.
Эти фосфолипиды у архей обладают четырьмя необычными чертами:
- У бактерий и эукариот мембраны состоят главным образом из глицерин-сложноэфирных липидов, тогда как у архей они сложены из глицерин-эфирных липидов[63]. Различается тип связи между остатками липидов и глицерина. Связи двух типов обозначены жёлтым на схеме справа. В сложноэфирных липидах связь сложноэфирная, а в эфирных — эфирная. Эфирные связи химически более стойкие, чем сложноэфирные. Эта стабильность помогает археям выживать при высоких температурах, а также в сильнокислых и сильнощелочных средах[64]. Бактерии и эукариоты содержат некоторое количество эфирных липидов, но по сравнению с археями они не являются главной составляющей мембран.
- Имеется отличие в стереохимии — у архей асимметрический центр глицериновой составляющей имеет L-конфигурацию, а не D-, как у других организмов. Поэтому для синтеза фосфолипидов археи используют совершенно другие ферменты, чем бактерии и эукариоты. Такие ферменты появились очень рано в истории жизни, что указывает на то, что археи рано отделились от двух других доменов[62].
- Липидные «хвосты» архей химически отличны от таковых у других организмов. Основу липидов архей составляет изопреноидная боковая цепь, и их липиды представляют собой длинные цепи с множеством побочных ветвей, иногда даже с циклопропановыми и циклогексановыми кольцами[65]. Хотя изопреноиды играют важную роль в биохимии многих организмов, только археи используют их для создания фосфолипидов. Предполагают, что эти разветвлённые цепи, как и эфирные связи, служат для приспособления к обитанию при высоких температурах. Установлено, что изопреноидные мембраны сохраняют в широком диапазоне температур (0—100 °C) жидкокристаллическое состояние, что необходимо для их нормального биологического функционирования. Проницаемость таких мембран для ионов и низкомолекулярных органических веществ также мало изменяется с повышением температуры, в отличие от мембран из «обычных» липидов, у которых она резко возрастает[66].
- У некоторых архей липидный бислой заменяется монослоем. Фактически при этом липидные «хвосты» двух разных фосфолипидных молекул сливаются с образованием одной молекулы с двумя полярными головками. Эти слияния делают мембрану более стойкой и лучше приспособленной для суровых условий[67]. К примеру, ферроплазма имеет липиды этого типа, и они помогают ей выживать в сильнокислых условиях[68].
Клеточная стенка
Большинство архей (но не Thermoplasma и Ferroplasma) обладают клеточной стенкой[54]. У большей части из них она сформирована молекулами поверхностных белков, образующих наружный S-слой[69]. S-слой представляет собой жёсткую сетку из белковых молекул, покрывающих клетку снаружи, подобно кольчуге[70]. Этот слой защищает клетку от физических и химических воздействий, а также предотвращает контакт макромолекул с клеточной мембраной[71]. В отличие от бактерий, клеточная стенка архей не содержит пептидогликан[72]. Метанобактерии (лат. Methanobacteriales) имеют клеточные стенки, содержащие псевдопептидогликан, который напоминает пептидогликан эубактерий по морфологии, функции и физической структуре, но отличен по химической: в нём нет остатков D-аминокислот и N-ацетилмурамовой кислоты[71].
Жгутики
Жгутик архей иногда называют археллум[73]. Жгутики архей работают так же, как и у бактерий: их длинные нити приводятся в движение вращательным механизмом в основании жгутика. Этот механизм работает за счёт трансмембранного протонного градиента. Тем не менее жгутики архей значительно отличаются от бактериальных по строению и способу сборки[60]. Два типа жгутиков развились из разных предковых структур. Бактериальный жгутик и система секреции III типа имели общую предковую структуру[74][75], а архейный жгутик произошёл от бактериальных пилей IV типа[76]. Жгутик бактерий полый и собирается из субъединиц, которые проходят вверх по центральной поре к концу жгутика. Жгутики же архей строятся путём добавления субъединиц в их основание[77]. Кроме того, в отличие от бактериальных жгутиков, в жгутики архей входит несколько видов флагеллинов.
Метаболизм
Археи демонстрируют огромное разнообразие химических реакций, протекающих в их клетках в процессе метаболизма, а также источников энергии. Эти реакции классифицируются по группам питания в зависимости от источников энергии и углерода. Некоторые археи получают энергию из неорганических соединений, таких как сера или аммиак (они являются литотрофами). К ним относятся нитрифицирующие археи, метаногены и анаэробные метаноокислители[78]. В окислительно-восстановительных реакциях одно соединение отдаёт электроны другому, а выделяющаяся при этом энергия служит топливом для осуществления различных клеточных процессов. Соединение, отдающее электроны, называется донором, а принимающее — акцептором. Выделяющаяся энергия идёт на образование АТФ путём хемиосмоса. В сущности, это основной процесс, протекающий в митохондриях эукариотических клеток[79].
Другие группы архей используют в качестве источника энергии солнечный свет (их называют фототрофами). Однако ни один из этих организмов не образует кислород в процессе фотосинтеза[79]. Многие базовые метаболические процессы являются общими для всех форм жизни, например, археи используют модифицированный вариант гликолиза (путь Энтнера-Дудорова), а также полный или частичный цикл Кребса (трикарбоновых кислот)[28]. Это, вероятно, отражает раннее возникновение этих путей в истории жизни и их высокую эффективность[80].
| Тип питания | Источник энергии | Источник углерода | Примеры |
|---|---|---|---|
| Фототрофы | Солнечный свет | Органические соединения | Halobacteria |
| Литотрофы | Неорганические соединения | Органические соединения или фиксация углерода | Ferroglobus, Methanobacteria, Pyrolobus |
| Органотрофы | Органические соединения | Органические соединения или фиксация углерода | Pyrococcus, Sulfolobus, Methanosarcinales |
Некоторые эвриархеоты являются метаногенами и обитают в анаэробных средах, таких как болота. Такой тип метаболизма появился рано, и возможно даже, что первый свободноживущий организм был метаногеном[81]. Обычная для этих организмов биохимическая реакция представляет собой окисление водорода с использованием углекислого газа в качестве акцептора электронов. Для осуществления метаногенеза необходимо множество различных коферментов, уникальных для этих архей, таких как кофермент М и метанофуран[82]. Некоторые органические соединения, такие как спирты, уксусная и муравьиная кислоты, могут использоваться метаногенами в качестве альтернативных акцепторов электронов. Подобные реакции протекают у архей, живущих в пищеварительном тракте. У ацидотрофных архей уксусная кислота распадается непосредственно на метан и углекислый газ. Такие ацидотрофные археи относятся к отряду Methanosarcinales. Они являются важной составляющей сообществ микроорганизмов, продуцирующих биогаз[83].
Другие археи используют атмосферный углекислый газ как источник углерода благодаря процессу фиксации углерода (то есть являются автотрофами). Этот процесс включает в себя либо сильно изменённый цикл Кальвина[84], либо метаболический путь, известный как 3-гидроксилпропионат/4-гидроксибутиратный цикл[85]. Кренархеоты также используют обратный цикл Кребса, а эвриархеоты — восстановительный ацетил-СоА процесс[86]. Фиксация углерода осуществляется за счёт энергии, получаемой из неорганических соединений. Ни один известный вид архей не фотосинтезирует[87]. Источники энергии, которые используют археи, чрезвычайно разнообразны, начиная от окисления аммиака Nitrosopumilales[88][89] до окисления сероводорода или элементарной серы, проводимого Sulfolobus, при этом в качестве акцепторов электронов могут использоваться кислород или ионы металлов[79].

Фототрофные археи используют солнечный свет для получения химической энергии в виде АТФ. У Halobacteria активируемые светом ионные насосы как бактериородопсин и галородопсин создают ионный градиент путём выкачивания ионов из клетки через плазматическую мембрану. Запасённая в этом электрохимическом градиенте энергия преобразуется в АТФ с помощью АТФ-синтазы[48]. Этот процесс представляет собой форму фотофосфорилирования. Способность этих насосов переносить ионы через мембраны при освещении обусловлена изменениями, которые происходят в структуре ретинолового кофактора, скрытого в центре белка, под действием света[91].
Генетика
Как правило, археи имеют одиночную кольцевую хромосому, размер которой может достигать 5 751 492 пар нуклеотидов у Methanosarcina acetivorans[92], обладающей самым большим известным геномом среди архей. Одну десятую размера этого генома составляет геном с 490 885 парами нуклеотидов у Nanoarchaeum equitans, имеющего самый маленький известный геном среди архей; он содержит лишь 537 генов, кодирующих белки[93]. Также у архей обнаружены более мелкие независимые молекулы ДНК, так называемые плазмиды. Возможно, плазмиды могут передаваться между клетками при физическом контакте, в ходе процесса сходного с конъюгацией бактерий[94][95].
Археи могут поражаться вирусами, содержащими двухцепочечную ДНК. Вирусы архей часто неродственны другим группам вирусов и имеют различные необычные формы, включая бутылки, крючки и капли[97]. Эти вирусы были тщательно изучены на термофилах, в основном отрядов Sulfolobales и Thermoproteales[98]. В 2009 году был открыт вирус, содержащий одноцепочечную ДНК и поражающий галофильные археи[99]. Защитные реакции архей против вирусов могут включать механизм, близкий к РНК-интерференции эукариот[100][101].
Археи генетически отличны от эукариот и бактерий, причём до 15 % белков, кодируемых одним геномом археи, уникальны для этого домена, хотя функции большинства этих белков неизвестны[102]. Большая часть уникальных белков, функция которых известна, принадлежит эвриархеотам и задействована в метаногенезе. Белки, общие для архей, бактерий и эукариот, участвуют в основных клеточных функциях и касаются в основном транскрипции, трансляции и метаболизма нуклеотидов[103]. К другим особенностям архей можно отнести организацию генов, выполняющих связанные функции (к примеру, гены, ответственные за разные этапы одного и того же метаболического процесса), в опероны и большие отличия в строении генов тРНК и их аминоацил-тРНК синтетаз[103].
Транскрипция и трансляция архей больше напоминают эти процессы в клетках эукариот, чем бактерий, причём РНК-полимераза и рибосомы архей очень близки к аналогичным структурам у эукариот[104]. Хотя у архей есть лишь один тип РНК-полимеразы, по строению и функции в транскрипции она близка к РНК-полимеразе II эукариот, при этом схожие группы белков (главные факторы транскрипции) обеспечивают связывание РНК-полимеразы с промотором гена[105]. В то же время другие факторы транскрипции архей более близки к таковым у бактерий[106]. Процессинг РНК у архей проще, чем у эукариот, так как большинство генов архей не содержит интронов, хотя в генах их тРНК и рРНК их достаточно много[107], также они присутствуют в небольшом количестве генов, кодирующих белки[108][109].
Размножение
Археи размножаются бесполым путём: бинарным или множественным делением, фрагментацией или почкованием. Мейоза не происходит, поэтому даже если представители конкретного вида архей существуют более чем в одной форме, все они имеют одинаковый генетический материал[48]. Клеточное деление определяется клеточным циклом: после того, как хромосома реплицировалась и две дочерние хромосомы разошлись, клетка делится[110]. Детали изучены лишь у рода Sulfolobus, но особенности его цикла очень схожи с таковыми и у эукариот, и у бактерий. Репликация хромосом начинается с множественных точек начала репликации с помощью ДНК-полимеразы, похожей на аналогичные ферменты эукариот[111]. Однако белки, управляющие клеточным делением, такие как FtsZ, которые формируют сжимающее кольцо вокруг клетки, и компоненты септы, проходящей через центр клетки, схожи с их бактериальными эквивалентами[110].
Археи не образуют споры[112]. Некоторые виды Haloarchaea могут претерпевать смену фенотипа и существовать как клетки нескольких различных типов, включая толстостенные клетки, устойчивые к осмотическому шоку и позволяющие археям выживать в воде с низкой концентрацией соли. Однако эти структуры не служат для размножения, а скорее помогают археям осваивать новые среды обитания[113].
Экология

Среды обитания
Археи живут в широком диапазоне сред обитания и являются важной частью глобальной экосистемы, могут составлять до 20 % общей биомассы[114]. Первые открытые археи были экстремофилами[78]. Действительно, многие археи выживают при высоких температурах, часто свыше 100 °C, и обнаружены в гейзерах, чёрных курильщиках и маслосборниках. Другие приспособились к жизни в очень холодных условиях, в сильносолёных, сильнокислых и сильнощелочных средах, а также при высоком давлении — до 700 атмосфер (барофилы). Однако среди архей есть и мезофилы, обитающие в мягких условиях, в болотистых местностях, сточных водах, океанах и почве[10].
Экстремофильные археи относятся к четырём главным физиологическим группам: галофилам, термофилам, ацидофилам (кислотоустойчивые) и алкалифилам (щелочеустойчивые)[115]. Эти группы нельзя рассматривать в ранге типа или как другие самостоятельные таксоны. Они не взаимоисключают друг друга, и некоторые археи относятся одновременно к нескольким группам. Тем не менее, они являются удобной стартовой точкой для классификации.
Галофилы, включая род Halobacterium, живут в экстремально солёных средах, таких как солёные озёра, и при минерализации больше 20—25 % превосходят по численности своих соседей-бактерий[78]. Термофилы лучше всего растут на температурах свыше 45 °C в таких местах, как горячие источники; для гипертермофилов оптимальная температура — 80 °C и выше[116]. Methanopyrus kandleri (штамм 116) растёт при 122 °C, рекордно высокой температуре для всех организмов[117].
Другие археи обитают в очень кислых или щелочных средах[115]. Например, наиболее устойчивый ацидофил Picrophilus torridus растёт при pH = 0, что эквивалентно 1,2 молярной серной кислоте[118].
Устойчивость к экстремальным условиям внешней среды сделала архей центральной темой в обсуждениях возможных свойств жизни на других планетах[119]. Некоторые среды, в которых обитают экстремофилы, не сильно отличаются от таковых на Марсе[120], что наводит на мысль о возможном переносе таких устойчивых микроорганизмов между планетами на метеоритах[121].
Недавно несколько работ показали, что археи обитают не только в термофильных и мезофильных условиях, но также встречаются, иногда в большом количестве, и в местах с низкими температурами. Например, археи встречаются в холодных водах, таких как полярные моря[122]. Ещё более важно, что огромное количество архей обнаружено повсеместно в океанах в неэкстремальных условиях в составе планктона (как часть пикопланктона)[123]. Хотя эти археи могут присутствовать в поистине колоссальном количестве (до 40 % от общей биомассы микробов), почти ни один из этих видов не был изолирован, выращен и изучен в чистой культуре[124]. Поэтому наше понимание роли архей в экологии океана, их влияния на глобальный биогеохимический круговорот остаётся в значительной мере неполным[125]. Некоторые морские кренархеоты способны к нитрификации, поэтому вероятно, что они оказывают влияние на океанический круговорот азота[126], хотя эти океанические кренархеоты могут использовать и другие источники энергии[127]. Большое число архей также обнаружено в осадке, покрывающем океаническое дно, причём они составляют большинство живых клеток на глубине больше 1 м от уровня океанического дна[128][129].
Роль в круговороте веществ
Археи вторично используют такие элементы, как углерод, азот и серу в своих различных средах обитания. Хотя такие превращения необходимы для нормального функционирования экосистемы, археи могут также содействовать вредным изменениям, вызванным деятельностью человека, и даже вызвать загрязнение.
Археи осуществляют многие этапы круговорота азота. Это включает в себя как реакции, удаляющие азот из экосистемы, к примеру, азотное дыхание и денитрификация, так и процессы, в ходе которых поглощается азот, такие как усвоение нитратов и фиксация азота[130][131]. Недавно была открыта причастность архей к окислению аммиака. Эти реакции особенно важны в океанах[89][132]. Археи также играют важную роль в почвенном окислении аммиака. Они образуют нитриты, которые затем окисляются другими микробами в нитраты. Последние потребляются растениями и другими организмами[133].
В круговороте серы археи, живущие за счёт окисления соединений серы, получают их из каменистых пород и делают их доступными для других организмов. Однако виды, осуществляющие это, такие как Sulfolobus, образуют серную кислоту как побочный продукт, и существование таких организмов в заброшенных шахтах может, совместно с кислотными шахтными водами, причинить вред окружающей среде[134].
В круговороте углерода метаногены удаляют водород и играют важную роль в разложении органической материи популяциями микроорганизмов, выступающих как разлагатели в анаэробных экосистемах, таких как илы, болота и водоочистные сооружения[135]. Однако метан — один из самых распространённых газов в земной атмосфере, вызывающих парниковый эффект, достигая 18 % от общего объёма парниковых газов[136]. Он в 25 раз более эффективен по способности вызывать парниковый эффект, чем углекислый газ[137]. Метаногены (главный источник атмосферного метана) выделяют большую часть ежегодного выброса метана[138]. Поэтому эти археи причастны к созданию парникового эффекта на Земле и глобальному потеплению.
Взаимодействие с другими организмами

Хорошо изученные отношения между археями и другими организмами — мутуализм и комменсализм. Пока не существует чётких доказательств существования патогенных или паразитических видов архей[139][140]. Однако была предположена связь между некоторыми видами метаногенов и инфекциями полости рта[141][142]. Кроме того, вид Nanoarchaeum equitans, возможно, является паразитом другого вида архей, поскольку он выживает и размножается только на клетках кренархеота Ignicoccus hospitalis[143] и не приносит никакой очевидной выгоды своему хозяину[144]. С другой стороны, архееподобные ацидофильные наноорганизмы Ричмондских рудников (ARMAN)[145] иногда прикрепляются к клеткам других архей в биоплёнках кислых сточных вод рудников[146]. Природа этого взаимодействия не ясна, но, в отличие от случая Nanorchaeaum—Ignicoccus, сверхмелкие клетки ARMAN всегда остаются независимыми от клеток Thermoplasmatales.
Мутуализм
Один из хорошо понятных примеров мутуализма — взаимодействие простейших и метанообразующих архей, обитающих в пищеварительном тракте животных, способных переваривать целлюлозу, таких как жвачные и термиты[147]. В этих анаэробных условиях простейшие разлагают целлюлозу для получения энергии. В этом процессе в качестве побочного продукта освобождается водород, однако высокий его уровень сокращает получение энергии. Метаногены превращают водород в метан, и простейшие могут дальше нормально получать энергию[148].
В случае анаэробных простейших вроде Plagiopyla frontata археи живут внутри клетки простейшего и потребляют водород, образуемый в его гидрогеносомах[149][150]. Археи также взаимодействуют и с более крупными организмами. Например, морская архея Cenarchaeum symbiosum живёт внутри (как эндосимбионт) губки Axinella mexicana[151].
Комменсализм
Археи могут быть комменсалами, то есть существовать совместно с другим организмом, не принося ему ни пользы, ни вреда, но с выгодой для себя. К примеру, метаноген Methanobrevibacter smithii — наиболее типичный представитель архей в микрофлоре человека. Каждый десятый прокариот в человеческом пищеварительном тракте принадлежит к этому виду[152]. В пищеварительном тракте термитов и человека эти метаногены в действительности могут быть мутуалистами, взаимодействующими с другими микробами пищеварительного тракта и способствующими пищеварению[153]. Археи также взаимодействуют с другими организмами, к примеру, живут на внешней поверхности кораллов[154] и в части почвы, прилегающей к корням растений (ризосфере)[155][156].
Классификация
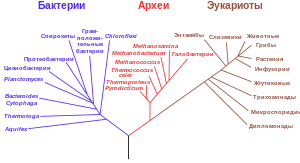
Классификация архей, как и прокариот в целом, быстро меняется и во многом остаётся спорной. Современные системы классификации стремятся объединить археи в группы организмов со схожими структурными свойствами и общими предками[157]. Эти классификации основаны на анализе структуры генов рРНК для установления родственных отношений между организмами (молекулярная филогенетика)[158]. Большую часть архей, выращиваемых в лабораториях и хорошо изученных, относят к двум главным типам, кренархеоты (Crenarchaeota) и эвриархеоты (Euryarchaeota). Другие группы были выделены в порядке рабочей гипотезы. Например, довольно необычный вид Nanoarchaeum equitans, открытый в 2003 году, был выделен в самостоятельный тип Nanoarchaeota[159]. Был также предложен новый тип Korarchaeota. Он объединяет небольшую группу термофильных видов, обладающих особенностями обоих основных типов, но более родственно близких к кренархеотам[160][161]. Другие недавно открытые виды имеют лишь дальнее родство с вышеперечисленными группами, например, архейные ацидофильные наноорганизмы Ричмондских рудников (ARMAN)[145], открытые в 2006 году[162] и являющиеся одними из самых мелких известных на сегодняшний момент организмов[163].

Разделение архей на виды также спорно. В биологии вид определяется как группа близкородственных организмов. Обычный критерий, которым пользуются в подобных ситуациях — организмы одного вида могут скрещиваться друг с другом, но не с особями других видов, — в данном случае не работает, поскольку археи размножаются только бесполым путём[164].
Археи демонстрируют высокий уровень горизонтального переноса генов между линиями. Некоторые исследователи предполагают, что особи можно объединять в популяции, похожие на виды, при условии высокой степени схожести их геномов и редко случающегося переноса генов между организмами с менее схожими геномами, как в случае рода ферроплазма (Ferroplasma)[165]. С другой стороны, изучение рода Halorubrum показало существование значимой передачи генов между дальнеродственными популяциями, что ограничивает применимость этого критерия[166]. Вторая проблема состоит в том, какое практическое значение может иметь подобное разделение на виды[167].
Современные данные о генетическом разнообразии архей фрагментарны, и общее число их видов не может быть оценено с какой-либо точностью[158]. Сравнительный анализ структур 16S рРНК архей позволил предположить существование 18—23 филогенетических групп уровня типов, причём представители лишь восьми групп выращены непосредственно в лаборатории и изучены (с учётом вероятной полифилии некоторых выделяемых в данный момент типов). Многие из этих гипотетических групп известны лишь по одной последовательности рРНК, что говорит о том, что пределы разнообразия этих организмов остаются неясными[168]. Многие бактерии также никогда не культивировались в лаборатории, что приводит к схожим проблемам при их характеристике[169].
По устоявшейся классификации на апрель 2021 года выделяют не менее 12 типов архей[2][170]:
- Crenarchaeota Garrity and Holt 2001 — Кренархеоты — термофилы, термоацидофилы, серные анаэробные бактерии;
- Euryarchaeota Garrity and Holt 2001 — Эвриархеоты — метаногенные и галофильные археи;
- Thaumarchaeota Brochier-Armanet et al. 2008 — в основном окислители аммония, как например, морской аммоний-окислитель Nitrosopumilus maritimus и аммоний-окислитель преимущественно почвенного происхождения Nitrososphaera gargensis. Недавние филогенетические исследования, основанные на сравнительном анализе структур рибосомальных белков и других важнейших генов, подтвердили существование этого типа[171];
- Candidatus Aigarchaeota Nunoura et al. 2011
- Candidatus Diapherotrites Rinke et al. 2013
- Candidatus Korarchaeota Barns et al. 1996 — Корархеоты — ДНК обнаружена в геотермальных источниках США, Исландии, на рисовых полях Японии, культивируемые виды пока неизвестны;
- Candidatus Lokiarchaeota Spang et al. 2015 — наиболее известный представитель, Lokiarchaeum, выделен на основании генома, собранного при метагеномном анализе образцов, полученных рядом с гидротермальными источниками в Атлантическом океане на глубине 2,35 км[172];
- Candidatus Nanoarchaeota Huber et al. 2002 — Наноархеоты — единственные известные представители Nanoarchaeum equitans и Nanobsidianus stetteri;
- Candidatus Nanohaloarchaeota Rinke et al. 2013
- Candidatus Parvarchaeota Rinke et al. 2013
- Candidatus Undinarchaeota Dombrowski et al. 2020
- Candidatus Verstraetearchaeota Vanwonterghem et al. 2016
Выделяют ещё несколько типов на основании данных метагеномики, в том числе:

Филогенетический анализ показал, что Lokiarchaeota и эукариоты образуют монофилетическую кладу — в их геномах обнаружены близкие гены, например, гены, кодирующие белки, отвечающие за изменение формы клеточной мембраны, определение формы клетки и динамический цитоскелет. Результаты этого исследования служат подтверждением так называемой двухдоменной, или эоцитной гипотезы, согласно которой эукариоты появились как особая группа внутри архей, близкая к Lokiarchaeota и приобретшая митохондрии в результате эндосимбиоза[172].
В январе 2016 года были опубликованы результаты метагеномных исследований по реконструкции геномов архей из морских осадочных отложений, которые свидетельствуют об обнаружении нового типа архей — Thorarchaeota. Организмы этой группы способны к образованию ацетата при деградации белков. Они также имеют гены, необходимые для восстановления элементарной серы и тиосульфата, поэтому эти организмы участвуют в круговороте серы[173].
Значение в технологии и промышленности
Экстремофильные археи, особенно устойчивые к высоким температурам или повышенной кислотности/щёлочности среды, являются источником ферментов, работающих в этих суровых условиях[174][175]. Эти ферменты находят множество применений. Например, термостабильные ДНК-полимеразы, такие как Pfu ДНК-полимераза вида Pyrococcus furiosus, полностью изменили молекулярную биологию, дав возможность использовать полимеразную цепную реакцию для простого и быстрого клонирования ДНК. В промышленности амилазы, галактозидазы и пуллуланазы других видов Pyrococcus, функционирующие при температуре свыше 100 °C, применяются при производстве продуктов питания при высоких температурах, к примеру, при производстве молока и сыворотки с низким содержанием лактозы[176]. Ферменты этих термофильных архей остаются очень стабильными в органических растворителях, что позволяет использовать их в безопасных для окружающей среды процессах в зелёной химии для синтеза органических соединений[175]. Их стабильность делает эти ферменты удобными для использования в структурной биологии, поэтому аналоги ферментов бактерий и эукариот, получаемые из экстремофильных архей, часто применяются в структурных исследованиях[177].
По сравнению с применением ферментов архей, использование самих организмов в биотехнологии развито слабо. Метанообразующие археи — важная часть водоочистных сооружений, так как они входят в сообщество микроорганизмов, осуществляющих анаэробное разложение и образование биогаза[178]. В обогащении полезных ископаемых ацидофильные археи могут использоваться для получения металлов из руд, в том числе золота, кобальта и меди[179].
Археи могут дать потенциально полезные антибиотики. Пока описано мало археоцинов, но предполагается, что их существует сотни, особенно много их может быть получено из родов Haloarchaea и Sulfolobus[180]. Эти антибиотики по структуре отличны от бактериальных, поэтому они могут иметь другие механизмы действия. Кроме того, они могут позволить создать селектируемые маркеры для использования в молекулярной биологии архей[181].
Примечания
- ↑ Archaea (англ.) на сайте Национального центра биотехнологической информации (NCBI). (Дата обращения: 3 сентября 2020).
- ↑ 1 2 Domain Archaea : [англ.] // LPSN[англ.]. — Leibniz Institute DSMZ[нем.]. (Дата обращения: 16 мая 2021).
- ↑ Археи : [арх. 3 января 2023] / Бонч-Осмоловская Е. А. // Анкилоз — Банка. — М. : Большая российская энциклопедия, 2005. — С. 311—312. — (Большая российская энциклопедия : [в 35 т.] / гл. ред. Ю. С. Осипов ; 2004—2017, т. 2). — ISBN 5-85270-330-3.
- ↑ Pace N. R. Time for a change (англ.) // Nature. — 2006. — May (vol. 441, no. 7091). — P. 289. — doi:10.1038/441289a. — . — PMID 16710401.
- ↑ Archaea: The Third Domain of Life. Дата обращения: 25 июля 2012. Архивировано 30 августа 2014 года.
- ↑ Staley J. T. The bacterial species dilemma and the genomic-phylogenetic species concept (англ.) // Philos. Trans. R. Soc. Lond., B, Biol. Sci. : journal. — 2006. — Vol. 361, no. 1475. — P. 1899—1909. — doi:10.1098/rstb.2006.1914. — PMID 17062409. — PMC 1857736.
- ↑ Zuckerkandl E., Pauling L. Molecules as documents of evolutionary history (англ.) // J. Theor. Biol.[англ.] : journal. — 1965. — Vol. 8, no. 2. — P. 357—366. — doi:10.1016/0022-5193(65)90083-4. — PMID 5876245.
- ↑ Woese C. R., Kandler O., Wheelis M. L. Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1990. — Vol. 87, no. 12. — P. 4576—4579. — doi:10.1073/pnas.87.12.4576. — . — PMID 2112744. — PMC 54159. Архивировано 16 октября 2019 года.
- ↑ Woese C.R., Kandler O., Wheelis M.L. Towards a Natural System of Organisms: Proposal for the Domains Archaea, Bacteria, and Eucarya // Proc. Natl. Acad. Sci. USA. — 1990. — Т. 87. — С. 4576—4579. Архивировано 9 июля 2008 года.
- ↑ 1 2 DeLong E. F. Everything in moderation: archaea as 'non-extremophiles' (англ.) // Curr. Opin. Genet. Dev. : journal. — 1998. — Vol. 8, no. 6. — P. 649—654. — doi:10.1016/S0959-437X(98)80032-4. — PMID 9914204.
- ↑ Theron J., Cloete T. E. Molecular techniques for determining microbial diversity and community structure in natural environments (англ.) // Crit. Rev. Microbiol.[англ.] : journal. — 2000. — Vol. 26, no. 1. — P. 37—57. — doi:10.1080/10408410091154174. — PMID 10782339.
- ↑ Schmidt T. M. The maturing of microbial ecology (англ.) // Int. Microbiol.[англ.] : journal. — 2006. — Vol. 9, no. 3. — P. 217—223. — PMID 17061212. Архивировано 11 сентября 2008 года.
- ↑ Schopf J. Fossil evidence of Archaean life (англ.) // Philos Trans R Soc Lond B Biol Sci : journal. — 2006. — Vol. 361, no. 1470. — P. 869—885. — doi:10.1098/rstb.2006.1834. — PMID 16754604. — PMC 1578735. Архивировано 23 февраля 2011 года.
- ↑ Chappe B., Albrecht P., Michaelis W. Polar Lipids of Archaebacteria in Sediments and Petroleums (англ.) // Science : journal. — 1982. — July (vol. 217, no. 4554). — P. 65—66. — doi:10.1126/science.217.4554.65. — . — PMID 17739984.
- ↑ Brocks J. J., Logan G. A., Buick R., Summons R. E. Archean molecular fossils and the early rise of eukaryotes (англ.) // Science : journal. — 1999. — Vol. 285, no. 5430. — P. 1033—1036. — doi:10.1126/science.285.5430.1033. — PMID 10446042.
- ↑ Rasmussen B., Fletcher I. R., Brocks J. J., Kilburn M. R. Reassessing the first appearance of eukaryotes and cyanobacteria (англ.) // Nature : journal. — 2008. — October (vol. 455, no. 7216). — P. 1101—1104. — doi:10.1038/nature07381. — . — PMID 18948954.
- ↑ Hahn, Jürgen; Pat Haug. Traces of Archaebacteria in ancient sediments (англ.) // System Applied Microbiology. — 1986. — Vol. 7, no. Archaebacteria '85 Proceedings. — P. 178—183.
- ↑ Wang M., Yafremava L. S., Caetano-Anollés D., Mittenthal J. E., Caetano-Anollés G. Reductive evolution of architectural repertoires in proteomes and the birth of the tripartite world (англ.) // Genome Res. : journal. — 2007. — Vol. 17, no. 11. — P. 1572—1585. — doi:10.1101/gr.6454307. — PMID 17908824. — PMC 2045140.
- ↑ Woese C. R., Gupta R. Are archaebacteria merely derived 'prokaryotes'? (англ.) // Nature. — 1981. — Vol. 289, no. 5793. — P. 95—6. — doi:10.1038/289095a0. — . — PMID 6161309.
- ↑ 1 2 Woese C. R. The universal ancestor (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1998. — Vol. 95, no. 12. — P. 6854—6859. — doi:10.1073/pnas.95.12.6854. — . — PMID 9618502. — PMC 22660. Архивировано 18 сентября 2019 года.
- ↑ Kandler O. The early diversification of life and the origin of the three domains: A proposal. In: Wiegel J., Adams W.W., editors. Thermophiles: The keys to molecular evolution and the origin of life? Athens: Taylor and Francis, 1998: 19-31.
- ↑ Gribaldo S., Brochier-Armanet C. The origin and evolution of Archaea: a state of the art (англ.) // Philos. Trans. R. Soc. Lond., B, Biol. Sci. : journal. — 2006. — Vol. 361, no. 1470. — P. 1007—1022. — doi:10.1098/rstb.2006.1841. — PMID 16754611. — PMC 1578729. Архивировано 4 июня 2012 года. Архивированная копия. Дата обращения: 21 июля 2012. Архивировано из оригинала 4 июня 2012 года.
- ↑ 1 2 Woese C. R. There must be a prokaryote somewhere: microbiology's search for itself (англ.) // Microbiology and Molecular Biology Reviews[англ.] : journal. — American Society for Microbiology[англ.], 1994. — 1 March (vol. 58, no. 1). — P. 1—9. — PMID 8177167. — PMC 372949. Архивировано 18 сентября 2019 года.
- ↑ Information is from Willey J.M., Sherwood L.M., Woolverton C.J. Microbiology 7th ed. (2008), Ch. 19 pp. 474—475, except where noted.
- ↑ Talbert P. B., Henikoff S. Histone variants – ancient wrap artists of the epigenome (англ.) // Nature Reviews Molecular Cell Biology : journal. — 2010. — Vol. 11. — P. 264—275. — doi:10.1038/nrm2861.
- ↑ Sandman K., Reeve J. N. Archaeal histones and the origin of the histone fold (англ.) // Curr. Opin. Microbiol : journal. — 2006. — Vol. 9. — P. 520—525. — doi:10.1016/j.mib.2006.08.003.
- ↑ у бактерий трансляция начинается с формилметионина
- ↑ 1 2 3 4 Zillig W. Comparative biochemistry of Archaea and Bacteria (англ.) // Curr. Opin. Gen. Dev.. — 1991. — December (vol. 1, no. 4). — P. 544—551. — doi:10.1016/S0959-437X(05)80206-0. — PMID 1822288.
- ↑ Bell S. D., Jackson S. P. Mechanism and regulation of transcription in archaea (англ.) // Curr. Opin. Microbiol. : journal. — 2001. — April (vol. 4, no. 2). — P. 208—213. — doi:10.1016/S1369-5274(00)00190-9. — PMID 11282478.
- ↑ Reeve J. N. Archaeal chromatin and transcription (англ.) // Microbiology[англ.] : journal. — Microbiology Society[англ.], 2003. — May (vol. 48, no. 3). — P. 587—598. — PMID 12694606.
- ↑ Kelman L. M., Kelman Z. Archaea: an archetype for replication initiation studies? (англ.) // Microbiology[англ.] : journal. — Microbiology Society[англ.], 2003. — May (vol. 48, no. 3). — P. 605—615. — PMID 12694608.
- ↑ Phillips G., Chikwana V. M., Maxwell A., et al. Discovery and characterization of an amidinotransferase involved in the modification of archaeal tRNA (англ.) // J. Biol. Chem. : journal. — 2010. — April (vol. 285, no. 17). — P. 12706—12713. — doi:10.1074/jbc.M110.102236. — PMID 20129918. — PMC 2857094.
- ↑ Koonin E.V., Mushegian A.R., Galperin M.Y., Walker D.R. Comparison of archaeal and bacterial genomes: computer analysis of protein sequences predicts novel functions and suggests a chimeric origin for the archaea. Mol Microbiol 1997; 25:619-637.
- ↑ 1 2 3 4 5 Gupta R. S. Protein phylogenies and signature sequences: A reappraisal of evolutionary relationships among archaebacteria, eubacteria, and eukaryotes (англ.) // Microbiology and Molecular Biology Reviews[англ.] : journal. — American Society for Microbiology[англ.], 1998. — Vol. 62. — P. 1435—1491. Архивировано 15 марта 2021 года.
- ↑ Koch A.L. Were Gram-positive rods the first bacteria? Trends Microbiol 2003; 11(4):166-170.
- ↑ 1 2 3 Gupta R. S. What are archaebacteria: life's third domain or monoderm prokaryotes related to gram-positive bacteria? A new proposal for the classification of prokaryotic organisms (англ.) // Mol. Microbiol : journal. — 1998. — Vol. 29. — P. 695—708. Архивировано 6 июля 2017 года.
- ↑ Brown J.R., Masuchi Y., Robb F.T., Doolittle W.F. Evolutionary relationships of bacterial and archaeal glutamine synthetase genes. J Mol Evol 1994; 38(6):566-576.
- ↑ 1 2 3 Gupta, R.S.(2000) The natural evolutionary relationships among prokaryotes. Crit. Rev. Microbiol. 26: 111—131.
- ↑ Gupta R.S. Molecular Sequences and the Early History of Life. In: Sapp J., editor. Microbial Phylogeny and Evolution: Concepts and Controversies. New York: Oxford University Press, 2005: 160—183.
- ↑ Cavalier-Smith T. The neomuran origin of archaebacteria, the negibacterial root of the universal tree and bacterial megaclassification. Int J Syst Evol Microbiol 2002; 52(Pt 1):7-76.
- ↑ Valas R.E., Bourne P.E.: 2011 The origin of a derived superkingdom: how a Gram-positive bacterium crossed the desert to become an archaeon. Biol Direct 6: 16.
- ↑ Skophammer R.G., Herbold C.W., Rivera M.C., Servin J.A., Lake J.A. Evidence that the root of the tree of life is not within the Archaea. Mol Biol Evol 2006; 23(9):1648-1651.
- ↑ Lake J. A. Origin of the eukaryotic nucleus determined by rate-invariant analysis of rRNA sequences (англ.) // Nature : journal. — 1988. — January (vol. 331, no. 6152). — P. 184—186. — doi:10.1038/331184a0. — . — PMID 3340165.
- ↑ Nelson K. E., Clayton R. A., Gill S. R., ; Utterback, T.R.; Malek, J.A.; Linher, K.D.; Garrett, M.M.; Stewart, A.M.; Cotton, M.D.; Pratt, M.S.; Phillips, C.A.; Richardson, D.; Heidelberg, J.; Sutton, G.G.; Fleischmann, R.D.; Eisen, J.A.; White, O.; Salzberg, S.L.; Smith, H.O.; Venter, J.C.; Fraser, C.M. et al. Evidence for lateral gene transfer between Archaea and bacteria from genome sequence of Thermotoga maritima (англ.) // Nature : journal. — 1999. — Vol. 399, no. 6734. — P. 323—329. — doi:10.1038/20601. — . — PMID 10360571.
- ↑ Gouy M., Li W. H. Phylogenetic analysis based on rRNA sequences supports the archaebacterial rather than the eocyte tree (англ.) // Nature : journal. — 1989. — May (vol. 339, no. 6220). — P. 145—147. — doi:10.1038/339145a0. — . — PMID 2497353.
- ↑ Yutin N., Makarova K. S., Mekhedov S. L., Wolf Y.I., Koonin E.V. The deep archaeal roots of eukaryotes (англ.) // Molecular Biology and Evolution[англ.] : journal. — Oxford University Press, 2008. — May (vol. 25, no. 8). — P. 1619—1630. — doi:10.1093/molbev/msn108. — PMID 18463089. — PMC 2464739. Архивировано 3 мая 2009 года.
- ↑ Lake J. A. Origin of the eukaryotic nucleus determined by rate-invariant analysis of rRNA sequences (англ.) // Nature : journal. — 1988. — Vol. 331, no. 6152. — P. 184—186. — doi:10.1038/331184a0. — . — PMID 3340165.
- ↑ 1 2 3 Krieg, Noel. Bergey's Manual of Systematic Bacteriology (англ.). — US: Springer, 2005. — P. 21—6. — ISBN 978-0-387-24143-2.
- ↑ Barns, Sue and Burggraf, Siegfried. (1997) Crenarchaeota Архивная копия от 2 мая 2012 на Wayback Machine. Version 01 January 1997. in The Tree of Life Web Project
- ↑ Walsby, A. E. A square bacterium (англ.) // Nature. — 1980. — Vol. 283, no. 5742. — P. 69—71. — doi:10.1038/283069a0. — .
- ↑ Hara F., Yamashiro K., Nemoto N., et al. An actin homolog of the archaeon Thermoplasma acidophilum that retains the ancient characteristics of eukaryotic actin (англ.) // American Society for Microbiology[англ.] : journal. — 2007. — Vol. 189, no. 5. — P. 2039—2045. — doi:10.1128/JB.01454-06. — PMID 17189356. — PMC 1855749. Архивировано 27 мая 2020 года.
- ↑ Trent J. D., Kagawa H. K., Yaoi T., Olle E., Zaluzec N. J. Chaperonin filaments: the archaeal cytoskeleton? (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1997. — Vol. 94, no. 10. — P. 5383—5388. — doi:10.1073/pnas.94.10.5383. — . — PMID 9144246. — PMC 24687. Архивировано 27 мая 2020 года.
- ↑ Hixon W. G., Searcy D. G. Cytoskeleton in the archaebacterium Thermoplasma acidophilum? Viscosity increase in soluble extracts (англ.) // BioSystems : journal. — 1993. — Vol. 29, no. 2—3. — P. 151—160. — doi:10.1016/0303-2647(93)90091-P. — PMID 8374067.
- ↑ 1 2 Golyshina O. V., Pivovarova T. A., Karavaiko G. I., et al. Ferroplasma acidiphilum gen. nov., sp. nov., an acidophilic, autotrophic, ferrous-iron-oxidizing, cell-wall-lacking, mesophilic member of the Ferroplasmaceae fam. nov., comprising a distinct lineage of the Archaea (англ.) // Int. J. Syst. Evol. Microbiol.[англ.] : journal. — 2000. — 1 May (vol. 50, no. 3). — P. 997—1006. — PMID 10843038. Архивировано 13 ноября 2023 года.
- ↑ Hall-Stoodley L., Costerton J. W., Stoodley P. Bacterial biofilms: from the natural environment to infectious diseases (англ.) // Nat. Rev. Microbiol. : journal. — 2004. — Vol. 2, no. 2. — P. 95—108. — doi:10.1038/nrmicro821. — PMID 15040259.
- ↑ Kuwabara T., Minaba M., Iwayama Y., ; Kamekura, M. et al. Thermococcus coalescens sp. nov., a cell-fusing hyperthermophilic archaeon from Suiyo Seamount (англ.) // Int. J. Syst. Evol. Microbiol.[англ.] : journal. — 2005. — November (vol. 55, no. Pt 6). — P. 2507—2514. — doi:10.1099/ijs.0.63432-0. — PMID 16280518. (недоступная ссылка)
- ↑ Nickell S., Hegerl R., Baumeister W., Rachel R. Pyrodictium cannulae enter the periplasmic space but do not enter the cytoplasm, as revealed by cryo-electron tomography (англ.) // J. Struct. Biol.[англ.] : journal. — 2003. — Vol. 141, no. 1. — P. 34—42. — doi:10.1016/S1047-8477(02)00581-6. — PMID 12576018. Архивировано 27 июня 2018 года.
- ↑ Horn C., Paulmann B., Kerlen G., Junker N., Huber H. In vivo observation of cell division of anaerobic hyperthermophiles by using a high-intensity dark-field microscope (англ.) // American Society for Microbiology[англ.] : journal. — 1999. — 15 August (vol. 181, no. 16). — P. 5114—5118. — PMID 10438790. — PMC 94007. Архивировано 16 октября 2019 года.
- ↑ Rudolph C., Wanner G., Huber R. Natural communities of novel archaea and bacteria growing in cold sulfurous springs with a string-of-pearls-like morphology (англ.) // Appl. Environ. Microbiol.[англ.] : journal. — 2001. — May (vol. 67, no. 5). — P. 2336—2344. — doi:10.1128/AEM.67.5.2336-2344.2001. — PMID 11319120. — PMC 92875.
- ↑ 1 2 Thomas N. A., Bardy S. L., Jarrell K. F. The archaeal flagellum: a different kind of prokaryotic motility structure (англ.) // Microbiology and Molecular Biology Reviews[англ.] : journal. — American Society for Microbiology[англ.], 2001. — Vol. 25, no. 2. — P. 147—174. — doi:10.1111/j.1574-6976.2001.tb00575.x. — PMID 11250034.
- ↑ Rachel R., Wyschkony I., Riehl S., Huber H. The ultrastructure of Ignicoccus: evidence for a novel outer membrane and for intracellular vesicle budding in an archaeon (англ.) // Archaea : journal. — 2002. — March (vol. 1, no. 1). — P. 9—18. — doi:10.1155/2002/307480. — PMID 15803654. — PMC 2685547. Архивировано 24 февраля 2009 года. Архивированная копия. Дата обращения: 21 июля 2012. Архивировано из оригинала 24 февраля 2009 года.
- ↑ 1 2 Koga Y., Morii H. Biosynthesis of ether-type polar lipids in archaea and evolutionary considerations (англ.) // Microbiology and Molecular Biology Reviews[англ.] : journal. — American Society for Microbiology[англ.], 2007. — Vol. 71, no. 1. — P. 97—120. — doi:10.1128/MMBR.00033-06. — PMID 17347520. — PMC 1847378. Архивировано 27 мая 2020 года.
- ↑ De Rosa M., Gambacorta A., Gliozzi A. Structure, biosynthesis, and physicochemical properties of archaebacterial lipids (англ.) // Microbiology and Molecular Biology Reviews[англ.] : journal. — American Society for Microbiology[англ.], 1986. — 1 March (vol. 50, no. 1). — P. 70—80. — PMID 3083222. — PMC 373054. Архивировано 16 октября 2019 года.
- ↑ Albers S. V., van de Vossenberg J. L., Driessen A. J., Konings W. N. Adaptations of the archaeal cell membrane to heat stress (англ.) // Frontiers in Bioscience[англ.] : journal. — Frontiers in Bioscience[англ.], 2000. — September (vol. 5). — P. D813—20. — doi:10.2741/albers. — PMID 10966867. Архивировано 27 октября 2012 года.
- ↑ Damsté J. S., Schouten S., Hopmans E. C., van Duin A. C., Geenevasen J. A. Crenarchaeol: the characteristic core glycerol dibiphytanyl glycerol tetraether membrane lipid of cosmopolitan pelagic crenarchaeota (англ.) // J. Lipid Res.[англ.] : journal. — 2002. — October (vol. 43, no. 10). — P. 1641—1651. — doi:10.1194/jlr.M200148-JLR200. — PMID 12364548. Архивировано 27 мая 2020 года.
- ↑ Koga Y., Morii H. Recent advances in structural research on ether lipids from archaea including comparative and physiological aspects (англ.) // Biosci. Biotechnol. Biochem.[англ.] : journal. — 2005. — November (vol. 69, no. 11). — P. 2019—2034. — doi:10.1271/bbb.69.2019. — PMID 16306681. Архивировано 31 декабря 2008 года. Архивированная копия. Дата обращения: 21 июля 2012. Архивировано 31 декабря 2008 года.
- ↑ Hanford M. J., Peeples T. L. Archaeal tetraether lipids: unique structures and applications (англ.) // Appl. Biochem. Biotechnol.. — 2002. — January (vol. 97, no. 1). — P. 45—62. — doi:10.1385/ABAB:97:1:45. — PMID 11900115.
- ↑ Macalady J. L., Vestling M. M., Baumler D., Boekelheide N., Kaspar C. W., Banfield J. F. Tetraether-linked membrane monolayers in Ferroplasma spp: a key to survival in acid (англ.) // Extremophiles : journal. — 2004. — October (vol. 8, no. 5). — P. 411—419. — doi:10.1007/s00792-004-0404-5. — PMID 15258835.
- ↑ Sára M., Sleytr U. B. S-Layer proteins (англ.) // American Society for Microbiology[англ.] : journal. — 2000. — Vol. 182, no. 4. — P. 859—868. — doi:10.1128/JB.182.4.859-868.2000. — PMID 10648507. — PMC 94357. Архивировано 27 мая 2020 года.
- ↑ Engelhardt H., Peters J. Structural research on surface layers: a focus on stability, surface layer homology domains, and surface layer-cell wall interactions (англ.) // J Struct Biol[англ.] : journal. — 1998. — Vol. 124, no. 2—3. — P. 276—302. — doi:10.1006/jsbi.1998.4070. — PMID 10049812.
- ↑ 1 2 Kandler, O.; König, H. Cell wall polymers in Archaea (Archaebacteria) (англ.) // Cellular and Molecular Life Sciences (CMLS). — 1998. — Vol. 54, no. 4. — P. 305—308. — doi:10.1007/s000180050156. (недоступная ссылка)
- ↑ Howland, John L. The Surprising Archaea: Discovering Another Domain of Life (англ.). — Oxford: Oxford University Press, 2000. — P. 32. — ISBN 0-19-511183-4.
- ↑ Albers Sonja-Verena, Jarrell Ken F. The archaellum: how archaea swim (англ.) // Frontiers in Microbiology. — 2015. — 27 January (vol. 6). — ISSN 1664-302X. — doi:10.3389/fmicb.2015.00023.
- ↑ Gophna U., Ron E. Z., Graur D. Bacterial type III secretion systems are ancient and evolved by multiple horizontal-transfer events (англ.) // Gene[англ.] : journal. — Elsevier, 2003. — July (vol. 312). — P. 151—163. — doi:10.1016/S0378-1119(03)00612-7. — PMID 12909351. Архивировано 22 ноября 2017 года.
- ↑ Nguyen L., Paulsen I. T., Tchieu J., Hueck C. J., Saier M. H. Phylogenetic analyses of the constituents of Type III protein secretion systems (англ.) // J. Mol. Microbiol. Biotechnol. : journal. — 2000. — April (vol. 2, no. 2). — P. 125—144. — PMID 10939240.
- ↑ Ng S. Y., Chaban B., Jarrell K. F. Archaeal flagella, bacterial flagella and type IV pili: a comparison of genes and posttranslational modifications (англ.) // J. Mol. Microbiol. Biotechnol. : journal. — 2006. — Vol. 11, no. 3—5. — P. 167—191. — doi:10.1159/000094053. — PMID 16983194.
- ↑ Bardy S. L., Ng S. Y., Jarrell K. F. Prokaryotic motility structures (англ.) // Microbiology[англ.] : journal. — Microbiology Society[англ.], 2003. — February (vol. 149, no. Pt 2). — P. 295—304. — doi:10.1099/mic.0.25948-0. — PMID 12624192.
- ↑ 1 2 3 Valentine D. L. Adaptations to energy stress dictate the ecology and evolution of the Archaea (англ.) // Nat. Rev. Microbiol. : journal. — 2007. — Vol. 5, no. 4. — P. 316—323. — doi:10.1038/nrmicro1619. — PMID 17334387.
- ↑ 1 2 3 Schäfer G., Engelhard M., Müller V. Bioenergetics of the Archaea (англ.) // Microbiology and Molecular Biology Reviews[англ.] : journal. — American Society for Microbiology[англ.], 1999. — 1 September (vol. 63, no. 3). — P. 570—620. — PMID 10477309. — PMC 103747. Архивировано 16 октября 2019 года.
- ↑ Romano A., Conway T. Evolution of carbohydrate metabolic pathways (англ.) // Res Microbiol. — 1996. — Vol. 147, no. 6—7. — P. 448—455. — doi:10.1016/0923-2508(96)83998-2. — PMID 9084754.
- ↑ Koch A. How did bacteria come to be? (англ.) // Advances in Microbial Physiology[англ.] : journal. — Academic Press, 1998. — Vol. 40. — P. 353—399. — doi:10.1016/S0065-2911(08)60135-6. — PMID 9889982.
- ↑ DiMarco A. A., Bobik T. A., Wolfe R. S. Unusual coenzymes of methanogenesis (англ.) // Annu. Rev. Biochem.[англ.] : journal. — 1990. — Vol. 59. — P. 355—394. — doi:10.1146/annurev.bi.59.070190.002035. — PMID 2115763.
- ↑ Klocke M., Nettmann E., Bergmann I., et al. Characterization of the methanogenic Archaea within two-phase biogas reactor systems operated with plant biomass (англ.) // Syst. Appl. Microbiol.[англ.] : journal. — 2008. — May (vol. 31, no. 3). — P. 190—205. — doi:10.1016/j.syapm.2008.02.003. — PMID 18501543.
- ↑ Mueller-Cajar O., Badger M. R. New roads lead to Rubisco in archaebacteria (англ.) // BioEssays[англ.] : journal. — 2007. — August (vol. 29, no. 8). — P. 722—724. — doi:10.1002/bies.20616. — PMID 17621634.
- ↑ Berg I. A., Kockelkorn D., Buckel W., Fuchs G. A 3-hydroxypropionate/4-hydroxybutyrate autotrophic carbon dioxide assimilation pathway in Archaea (англ.) // Science : journal. — 2007. — December (vol. 318, no. 5857). — P. 1782—1786. — doi:10.1126/science.1149976. — . — PMID 18079405.
- ↑ Thauer R. K. Microbiology. A fifth pathway of carbon fixation (англ.) // Science. — 2007. — December (vol. 318, no. 5857). — P. 1732—1733. — doi:10.1126/science.1152209. — PMID 18079388.
- ↑ Bryant D. A., Frigaard N. U. Prokaryotic photosynthesis and phototrophy illuminated (англ.) // Trends Microbiol.[англ.] : journal. — 2006. — November (vol. 14, no. 11). — P. 488—496. — doi:10.1016/j.tim.2006.09.001. — PMID 16997562.
- ↑ Könneke M., Bernhard A. E., de la Torre J. R., Walker C. B., Waterbury J. B., Stahl D. A. Isolation of an autotrophic ammonia-oxidizing marine archaeon (англ.) // Nature : journal. — 2005. — September (vol. 437, no. 7058). — P. 543—546. — doi:10.1038/nature03911. — . — PMID 16177789.
- ↑ 1 2 Francis C. A., Beman J. M., Kuypers M. M. New processes and players in the nitrogen cycle: the microbial ecology of anaerobic and archaeal ammonia oxidation (англ.) // ISME J[англ.] : journal. — 2007. — May (vol. 1, no. 1). — P. 19—27. — doi:10.1038/ismej.2007.8. — PMID 18043610.
- ↑ Основано на PDB 1FBB Архивная копия от 3 марта 2016 на Wayback Machine. Данные опубликованы в Subramaniam S., Henderson R. Molecular mechanism of vectorial proton translocation by bacteriorhodopsin (англ.) // Nature : journal. — 2000. — August (vol. 406, no. 6796). — P. 653—657. — doi:10.1038/35020614. — PMID 10949309.
- ↑ Lanyi J. K. Bacteriorhodopsin (англ.) // Annu. Rev. Physiol.. — 2004. — Vol. 66. — P. 665—688. — doi:10.1146/annurev.physiol.66.032102.150049. — PMID 14977418.
- ↑ Galagan J. E., Nusbaum C., Roy A., ; Allen, N; Naylor, J; Stange-Thomann, N; Dearellano, K; Johnson, R; Linton, L; Mcewan, P; Mckernan, K; Talamas, J; Tirrell, A; Ye, W; Zimmer, A; Barber, RD; Cann, I; Graham, DE; Grahame, DA; Guss, AM; Hedderich, R; Ingram-Smith, C; Kuettner, HC; Krzycki, JA; Leigh, JA; Li, W; Liu, J; Mukhopadhyay, B; Reeve, JN; Smith, K; Springer, TA; Umayam, LA; White, O; White, RH; Conway De Macario, E; Ferry, JG; Jarrell, KF; Jing, H; Macario, AJ; Paulsen, I; Pritchett, M; Sowers, KR; Swanson, RV; Zinder, SH; Lander, E; Metcalf, WW; Birren, B. et al. The genome of M. acetivorans reveals extensive metabolic and physiological diversity (англ.) // Genome Res. : journal. — 2002. — April (vol. 12, no. 4). — P. 532—542. — doi:10.1101/gr.223902. — PMID 11932238. — PMC 187521.
- ↑ Waters E., ; Lin, X; Mathur, E; Ni, J; Podar, M; Richardson, T; Sutton, GG; Simon, M; Soll, D; Stetter, KO; Short, JM; Noordewier, M. et al. The genome of Nanoarchaeum equitans: insights into early archaeal evolution and derived parasitism (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2003. — Vol. 100, no. 22. — P. 12984—12988. — doi:10.1073/pnas.1735403100. — . — PMID 14566062. — PMC 240731. Архивировано 16 октября 2019 года.
- ↑ Schleper C., Holz I., Janekovic D., Murphy J., Zillig W. A multicopy plasmid of the extremely thermophilic archaeon Sulfolobus effects its transfer to recipients by mating (англ.) // American Society for Microbiology[англ.] : journal. — 1995. — 1 August (vol. 177, no. 15). — P. 4417—4426. — PMID 7635827. — PMC 177192. Архивировано 29 мая 2012 года.
- ↑ Sota M; Top E. M. Horizontal Gene Transfer Mediated by Plasmids // Plasmids: Current Research and Future Trends (англ.). — Caister Academic Press[англ.], 2008.
- ↑ Xiang X., Chen L., Huang X., Luo Y., She Q., Huang L. Sulfolobus tengchongensis spindle-shaped virus STSV1: virus-host interactions and genomic features (англ.) // J. Virol.[англ.] : journal. — 2005. — Vol. 79, no. 14. — P. 8677—8686. — doi:10.1128/JVI.79.14.8677-8686.2005. — PMID 15994761. — PMC 1168784. Архивировано 16 октября 2019 года.
- ↑ Prangishvili D., Forterre P., Garrett R. A. Viruses of the Archaea: a unifying view (англ.) // Nat. Rev. Microbiol. : journal. — 2006. — Vol. 4, no. 11. — P. 837—848. — doi:10.1038/nrmicro1527. — PMID 17041631.
- ↑ Prangishvili D., Garrett R. A. Exceptionally diverse morphotypes and genomes of crenarchaeal hyperthermophilic viruses (англ.) // Biochem. Soc. Trans.[англ.] : journal. — 2004. — Vol. 32, no. Pt 2. — P. 204—208. — doi:10.1042/BST0320204. — PMID 15046572. Архивировано 18 апреля 2006 года.
- ↑ Pietilä M. K., Roine E., Paulin L., Kalkkinen N., Bamford D. H. An ssDNA virus infecting archaea; A new lineage of viruses with a membrane envelope (англ.) // Microbiology[англ.] : journal. — Microbiology Society[англ.], 2009. — March (vol. 72, no. 2). — P. 307—319. — doi:10.1111/j.1365-2958.2009.06642.x. — PMID 19298373.
- ↑ Mojica F. J., Díez-Villaseñor C., García-Martínez J., Soria E. Intervening sequences of regularly spaced prokaryotic repeats derive from foreign genetic elements (англ.) // J. Mol. Evol.[англ.] : journal. — 2005. — Vol. 60, no. 2. — P. 174—182. — doi:10.1007/s00239-004-0046-3. — PMID 15791728.
- ↑ Makarova K. S., Grishin N. V., Shabalina S. A., Wolf Y. I., Koonin E. V. A putative RNA-interference-based immune system in prokaryotes: computational analysis of the predicted enzymatic machinery, functional analogies with eukaryotic RNAi, and hypothetical mechanisms of action (англ.) // Biol. Direct[англ.] : journal. — 2006. — Vol. 1. — P. 7. — doi:10.1186/1745-6150-1-7. — PMID 16545108. — PMC 1462988.
- ↑ Graham D. E., Overbeek R., Olsen G. J., Woese C. R. An archaeal genomic signature (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2000. — Vol. 97, no. 7. — P. 3304—3308. — doi:10.1073/pnas.050564797. — . — PMID 10716711. — PMC 16234.
- ↑ 1 2 Gaasterland T. Archaeal genomics (англ.) // Curr. Opin. Microbiol.. — 1999. — Vol. 2, no. 5. — P. 542—547. — doi:10.1016/S1369-5274(99)00014-4. — PMID 10508726.
- ↑ Allers T., Mevarech M. Archaeal genetics — the third way (англ.) // Nat. Rev. Genet. : journal. — 2005. — Vol. 6, no. 1. — P. 58—73. — doi:10.1038/nrg1504. — PMID 15630422.
- ↑ Werner F. Structure and function of archaeal RNA polymerases (англ.) // Microbiology[англ.] : journal. — Microbiology Society[англ.], 2007. — September (vol. 65, no. 6). — P. 1395—1404. — doi:10.1111/j.1365-2958.2007.05876.x. — PMID 17697097.
- ↑ Aravind L., Koonin E. V. DNA-binding proteins and evolution of transcription regulation in the archaea (англ.) // Nucleic Acids Res. : journal. — 1999. — Vol. 27, no. 23. — P. 4658—4670. — doi:10.1093/nar/27.23.4658. — PMID 10556324. — PMC 148756. Архивировано 16 октября 2019 года.
- ↑ Lykke-Andersen J., Aagaard C., Semionenkov M., Garrett R. A. Archaeal introns: splicing, intercellular mobility and evolution (англ.) // Trends Biochem. Sci.[англ.] : journal. — 1997. — September (vol. 22, no. 9). — P. 326—331. — doi:10.1016/S0968-0004(97)01113-4. — PMID 9301331.
- ↑ Watanabe Y., Yokobori S., Inaba T., et al. Introns in protein-coding genes in Archaea (англ.) // FEBS Lett.[англ.] : journal. — 2002. — January (vol. 510, no. 1—2). — P. 27—30. — doi:10.1016/S0014-5793(01)03219-7. — PMID 11755525.
- ↑ Yoshinari S., Itoh T., Hallam S. J., et al. Archaeal pre-mRNA splicing: a connection to hetero-oligomeric splicing endonuclease (англ.) // Biochem. Biophys. Res. Commun.[англ.] : journal. — 2006. — August (vol. 346, no. 3). — P. 1024—1032. — doi:10.1016/j.bbrc.2006.06.011. — PMID 16781672.
- ↑ 1 2 Bernander R. Archaea and the cell cycle (англ.) // Microbiology[англ.] : journal. — Microbiology Society[англ.], 1998. — Vol. 29, no. 4. — P. 955—961. — doi:10.1046/j.1365-2958.1998.00956.x. — PMID 9767564.
- ↑ Kelman L. M., Kelman Z. Multiple origins of replication in archaea (англ.) // Trends Microbiol.[англ.] : journal. — 2004. — Vol. 12, no. 9. — P. 399—401. — doi:10.1016/j.tim.2004.07.001. — PMID 15337158.
- ↑ Onyenwoke R. U., Brill J. A., Farahi K., Wiegel J. Sporulation genes in members of the low G+C Gram-type-positive phylogenetic branch ( Firmicutes) (англ.) // Arch. Microbiol.[англ.] : journal. — 2004. — Vol. 182, no. 2—3. — P. 182—192. — doi:10.1007/s00203-004-0696-y. — PMID 15340788.
- ↑ Kostrikina N. A., Zvyagintseva I. S., Duda V. I. Cytological peculiarities of some extremely halophilic soil archaeobacteria (англ.) // Arch. Microbiol.[англ.] : journal. — 1991. — Vol. 156, no. 5. — P. 344—349. — doi:10.1007/BF00248708.
- ↑ DeLong E. F., Pace N. R. Environmental diversity of bacteria and archaea (англ.) // Syst. Biol. : journal. — 2001. — Vol. 50, no. 4. — P. 470—478. — doi:10.1080/106351501750435040. — PMID 12116647.
- ↑ 1 2 Pikuta E. V., Hoover R. B., Tang J. Microbial extremophiles at the limits of life (англ.) // Crit. Rev. Microbiol.[англ.] : journal. — 2007. — Vol. 33, no. 3. — P. 183—209. — doi:10.1080/10408410701451948. — PMID 17653987.
- ↑ Madigan M. T., Martino J. M. Brock Biology of Microorganisms (англ.). — 11th. — Pearson, 2006. — P. 136. — ISBN 0-13-196893-9.
- ↑ Takai K., Nakamura K., Toki T., Tsunogai U., Miyazaki M., Miyazaki J., Hirayama H., Nakagawa S., Nunoura T., Horikoshi K. Cell proliferation at 122 °C and isotopically heavy CH4 production by a hyperthermophilic methanogen under high-pressure cultivation (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2008. — Vol. 105, no. 31. — P. 10949—10954. — doi:10.1073/pnas.0712334105. — . — PMID 18664583. — PMC 2490668.
- ↑ Ciaramella M., Napoli A., Rossi M. Another extreme genome: how to live at pH 0 (англ.) // Trends Microbiol.[англ.] : journal. — 2005. — February (vol. 13, no. 2). — P. 49—51. — doi:10.1016/j.tim.2004.12.001. — PMID 15680761.
- ↑ Javaux E. J. Extreme life on Earth—past, present and possibly beyond (англ.) // Res. Microbiol. : journal. — 2006. — Vol. 157, no. 1. — P. 37—48. — doi:10.1016/j.resmic.2005.07.008. — PMID 16376523.
- ↑ Nealson K. H. Post-Viking microbiology: new approaches, new data, new insights (англ.) // Orig Life Evol Biosph : journal. — 1999. — January (vol. 29, no. 1). — P. 73—93. — doi:10.1023/A:1006515817767. — PMID 11536899. Архивировано 16 октября 2019 года.
- ↑ Davies P. C. The transfer of viable microorganisms between planets (англ.) // Ciba Found. Symp. : journal. — 1996. — Vol. 202. — P. 304—314. — PMID 9243022.
- ↑ López-García P., López-López A., Moreira D., Rodríguez-Valera F. Diversity of free-living prokaryotes from a deep-sea site at the Antarctic Polar Front (англ.) // FEMS Microbiol. Ecol.[англ.] : journal. — 2001. — July (vol. 36, no. 2—3). — P. 193—202. — PMID 11451524.
- ↑ Karner M. B., DeLong E. F., Karl D. M. Archaeal dominance in the mesopelagic zone of the Pacific Ocean (англ.) // Nature : journal. — 2001. — Vol. 409, no. 6819. — P. 507—510. — doi:10.1038/35054051. — PMID 11206545.
- ↑ Giovannoni S. J., Stingl U. Molecular diversity and ecology of microbial plankton (англ.) // Nature : journal. — 2005. — Vol. 427, no. 7057. — P. 343—348. — doi:10.1038/nature04158. — . — PMID 16163344.
- ↑ DeLong E. F., Karl D. M. Genomic perspectives in microbial oceanography (англ.) // Nature. — 2005. — September (vol. 437, no. 7057). — P. 336—342. — doi:10.1038/nature04157. — . — PMID 16163343.
- ↑ Konneke M., Bernhard A. E., de la Torre J. R., Walker C. B., Waterbury J. B., Stahl DA. Isolation of an autotrophic ammonia-oxidizing marine archaeon (англ.) // Nature : journal. — 2005. — Vol. 437, no. 7057. — P. 543—546. — doi:10.1038/nature03911. — . — PMID 16177789.
- ↑ Agogué H., Maaike B., Dinasquet J., Herndl GJ.; Agogué, H; Brink, M; Dinasquet, J; Herndl, G.J. Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic (англ.) // Nature : journal. — 2008. — Vol. 456, no. 7223. — P. 788—791. — doi:10.1038/nature07535. — . — PMID 19037244.
- ↑ Teske A., Sørensen K. B. Uncultured archaea in deep marine subsurface sediments: have we caught them all? (англ.) // ISME J[англ.] : journal. — 2008. — January (vol. 2, no. 1). — P. 3—18. — doi:10.1038/ismej.2007.90. — PMID 18180743.
- ↑ Lipp J. S., Morono Y., Inagaki F., Hinrichs K. U. Significant contribution of Archaea to extant biomass in marine subsurface sediments (англ.) // Nature : journal. — 2008. — July (vol. 454, no. 7207). — P. 991—994. — doi:10.1038/nature07174. — . — PMID 18641632.
- ↑ Cabello P., Roldán M. D., Moreno-Vivián C. Nitrate reduction and the nitrogen cycle in archaea (англ.) // Microbiology[англ.] : journal. — Microbiology Society[англ.], 2004. — November (vol. 150, no. Pt 11). — P. 3527—3546. — doi:10.1099/mic.0.27303-0. — PMID 15528644. Архивировано 7 июня 2011 года.
- ↑ Mehta M. P., Baross J. A. Nitrogen fixation at 92 degrees C by a hydrothermal vent archaeon (англ.) // Science : journal. — 2006. — December (vol. 314, no. 5806). — P. 1783—1786. — doi:10.1126/science.1134772. — . — PMID 17170307.
- ↑ Coolen M. J., Abbas B., van Bleijswijk J., et al. Putative ammonia-oxidizing Crenarchaeota in suboxic waters of the Black Sea: a basin-wide ecological study using 16S ribosomal and functional genes and membrane lipids (англ.) // Environ. Microbiol. : journal. — 2007. — April (vol. 9, no. 4). — P. 1001—1016. — doi:10.1111/j.1462-2920.2006.01227.x. — PMID 17359272.
- ↑ Leininger S., Urich T., Schloter M., et al. Archaea predominate among ammonia-oxidizing prokaryotes in soils (англ.) // Nature : journal. — 2006. — August (vol. 442, no. 7104). — P. 806—809. — doi:10.1038/nature04983. — . — PMID 16915287.
- ↑ Baker, B. J.; Banfield, J. F. Microbial communities in acid mine drainage (англ.) // FEMS Microbiology Ecology[англ.] : journal. — 2003. — Vol. 44, no. 2. — P. 139—152. — doi:10.1016/S0168-6496(03)00028-X. — PMID 19719632. (недоступная ссылка)
- ↑ Schimel J. Playing scales in the methane cycle: from microbial ecology to the globe (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2004. — August (vol. 101, no. 34). — P. 12400—12401. — doi:10.1073/pnas.0405075101. — . — PMID 15314221. — PMC 515073. Архивировано 16 октября 2019 года.
- ↑ EDGAR 3.2 Fast Track 2000. Дата обращения: 26 июня 2008. Архивировано 21 мая 2008 года.
- ↑ Annual Greenhouse Gas Index (AGGI) Indicates Sharp Rise in Carbon Dioxide and Methane in 2007 (23 апреля 2008). Дата обращения: 26 июня 2008. Архивировано 14 мая 2008 года.
- ↑ Trace Gases: Current Observations, Trends, and Budgets. Climate Change 2001. United Nations Environment Programme. Архивировано 5 августа 2012 года.
- ↑ Eckburg P., Lepp P., Relman D. Archaea and their potential role in human disease (англ.) // Infect Immun[англ.] : journal. — 2003. — Vol. 71, no. 2. — P. 591—596. — doi:10.1128/IAI.71.2.591-596.2003. — PMID 12540534. — PMC 145348.
- ↑ Cavicchioli R., Curmi P., Saunders N., Thomas T. Pathogenic archaea: do they exist? (англ.) // BioEssays[англ.] : journal. — 2003. — Vol. 25, no. 11. — P. 1119—1128. — doi:10.1002/bies.10354. — PMID 14579252.
- ↑ Lepp P., Brinig M., Ouverney C., Palm K., Armitage G., Relman D. Methanogenic Archaea and human periodontal disease (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2004. — Vol. 101, no. 16. — P. 6176—6181. — doi:10.1073/pnas.0308766101. — . — PMID 15067114. — PMC 395942.
- ↑ Vianna M. E., Conrads G., Gomes B. P., Horz H. P. Identification and quantification of archaea involved in primary endodontic infections (англ.) // J. Clin. Microbiol. : journal. — 2006. — April (vol. 44, no. 4). — P. 1274—1282. — doi:10.1128/JCM.44.4.1274-1282.2006. — PMID 16597851. — PMC 1448633. Архивировано 16 октября 2019 года.
- ↑ Waters E., Hohn M. J., Ahel I., ; Lin, X; Mathur, E; Ni, J; Podar, M; Richardson, T; Sutton, GG; Simon, M; Soll, D; Stetter, KO; Short, JM; Noordewier, M. et al. The genome of Nanoarchaeum equitans: insights into early archaeal evolution and derived parasitism (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2003. — October (vol. 100, no. 22). — P. 12984—12988. — doi:10.1073/pnas.1735403100. — . — PMID 14566062. — PMC 240731. Архивировано 16 октября 2019 года.
- ↑ Jahn U., Gallenberger M., Paper W., et al. Nanoarchaeum equitans and Ignicoccus hospitalis: new insights into a unique, intimate association of two archaea (англ.) // American Society for Microbiology[англ.] : journal. — 2008. — March (vol. 190, no. 5). — P. 1743—1750. — doi:10.1128/JB.01731-07. — PMID 18165302. — PMC 2258681. Архивировано 27 мая 2020 года.
- ↑ 1 2 3 рус. Архееподобные ацидофильные наноорганизмы Ричмондских рудников
- ↑ Baker B. J., Comolli L. R., Dick G. J., Hauser L. J., Hyatt D., Dill B. D., Land M. L., VerBerkmoes N. C., Hettich R. L., Banfield J. F. Enigmatic, ultrasmall, uncultivated Archaeaa (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2010. — May (vol. 107, no. 19). — P. 8806—8811. — doi:10.1073/pnas.0914470107. — PMID 20421484. — PMC 2889320. Архивировано 10 февраля 2021 года.
- ↑ Chaban B., Ng S. Y., Jarrell K. F. Archaeal habitats—from the extreme to the ordinary (англ.) // Canadian Journal of Microbiology[англ.] : journal. — NRC Research Press[англ.], 2006. — February (vol. 52, no. 2). — P. 73—116. — doi:10.1139/w05-147. — PMID 16541146.
- ↑ Schink B. Energetics of syntrophic cooperation in methanogenic degradation (англ.) // Microbiology and Molecular Biology Reviews[англ.] : journal. — American Society for Microbiology[англ.], 1997. — June (vol. 61, no. 2). — P. 262—280. — PMID 9184013. — PMC 232610.
- ↑ Lange, M; Lange, M.; Westermann, P; Westermann, P.; Ahring, BK; Ahring, B. K. Archaea in protozoa and metazoa (англ.) // Applied Microbiology and Biotechnology[англ.] : journal. — Springer, 2005. — Vol. 66, no. 5. — P. 465—474. — doi:10.1007/s00253-004-1790-4. — PMID 15630514.
- ↑ van Hoek A. H., van Alen T. A., Sprakel V. S., et al. Multiple acquisition of methanogenic archaeal symbionts by anaerobic ciliates (англ.) // Molecular Biology and Evolution[англ.] : journal. — Oxford University Press, 2000. — 1 February (vol. 17, no. 2). — P. 251—258. — PMID 10677847. Архивировано 16 октября 2019 года.
- ↑ Preston, C. M.; Wu, K. Y.; Molinski, T. F.; Delong, E. F. A psychrophilic crenarchaeon inhabits a marine sponge: Cenarchaeum symbiosum gen. nov., sp. nov (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1996. — Vol. 93, no. 13. — P. 6241—6246. — doi:10.1073/pnas.93.13.6241. — . — PMID 8692799. — PMC 39006.
- ↑ Eckburg P. B., Bik E. M., Bernstein C. N., et al. Diversity of the human intestinal microbial flora (англ.) // Science. — 2005. — June (vol. 308, no. 5728). — P. 1635—1638. — doi:10.1126/science.1110591. — . — PMID 15831718. — PMC 1395357.
- ↑ Samuel B. S., Gordon J. I. A humanized gnotobiotic mouse model of host-archaeal-bacterial mutualism (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2006. — June (vol. 103, no. 26). — P. 10011—10016. — doi:10.1073/pnas.0602187103. — . — PMID 16782812. — PMC 1479766.
- ↑ Wegley, L; Wegley, L.; Yu; Yu, Y.; Breitbart; Breitbart, M.; Casas; Casas, V.; Kline; Kline, D. I.; Rohwer; Rohwer, F. Coral-associated Archaea (англ.) // Marine Ecology Progress Series[англ.] : journal. — 2004. — Vol. 273. — P. 89—96. — doi:10.3354/meps273089. Архивировано 11 сентября 2008 года. Архивированная копия. Дата обращения: 21 июля 2012. Архивировано из оригинала 11 сентября 2008 года.
- ↑ Chelius M. K., Triplett E. W. The Diversity of Archaea and Bacteria in Association with the Roots of Zea mays L (англ.) // Microb. Ecol. : journal. — 2001. — April (vol. 41, no. 3). — P. 252—263. — doi:10.1007/s002480000087. — PMID 11391463.
- ↑ Simon H. M., Dodsworth J. A., Goodman R. M. Crenarchaeota colonize terrestrial plant roots (англ.) // Environ. Microbiol.. — 2000. — October (vol. 2, no. 5). — P. 495—505. — doi:10.1046/j.1462-2920.2000.00131.x. — PMID 11233158.
- ↑ Gevers D., Dawyndt P., Vandamme P., et al. Stepping stones towards a new prokaryotic taxonomy (англ.) // Philos. Trans. R. Soc. Lond., B, Biol. Sci. : journal. — 2006. — Vol. 361, no. 1475. — P. 1911—1916. — doi:10.1098/rstb.2006.1915. — PMID 17062410. — PMC 1764938. Архивировано 20 декабря 2012 года.
- ↑ 1 2 Robertson C. E., Harris J. K., Spear J. R., Pace N. R. Phylogenetic diversity and ecology of environmental Archaea (англ.) // Curr. Opin. Microbiol. : journal. — 2005. — Vol. 8, no. 6. — P. 638—642. — doi:10.1016/j.mib.2005.10.003. — PMID 16236543.
- ↑ Huber H., Hohn M. J., Rachel R., Fuchs T., Wimmer V. C., Stetter K. O. A new phylum of Archaea represented by a nanosized hyperthermophilic symbiont (англ.) // Nature : journal. — 2002. — Vol. 417, no. 6884. — P. 27—28. — doi:10.1038/417063a. — PMID 11986665.
- ↑ Barns S. M., Delwiche C. F., Palmer J. D., Pace N. R. Perspectives on archaeal diversity, thermophily and monophyly from environmental rRNA sequences (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1996. — Vol. 93, no. 17. — P. 9188—9193. — doi:10.1073/pnas.93.17.9188. — . — PMID 8799176. — PMC 38617. Архивировано 8 мая 2020 года.
- ↑ Elkins J. G., Podar M., Graham D. E., ; Goltsman, E; Barry, K; Koonin, EV; Hugenholtz, P; Kyrpides, N; Wanner, G; Richardson, P; Keller, M; Stetter, K.O. et al. A korarchaeal genome reveals insights into the evolution of the Archaea (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2008. — June (vol. 105, no. 23). — P. 8102—8107. — doi:10.1073/pnas.0801980105. — . — PMID 18535141. — PMC 2430366. Архивировано 27 мая 2020 года.
- ↑ Baker B. J., Tyson G. W., Webb R. I., Flanagan J., Hugenholtz P. and Banfield J. F. Lineages of acidophilic Archaea revealed by community genomic analysis. Science (англ.) // Science : journal. — 2006. — Vol. 314, no. 6884. — P. 1933—1935. — doi:10.1126/science.1132690. — . — PMID 17185602.
- ↑ Baker B. J., Comolli L. R., Dick G. J., et al. Enigmatic, ultrasmall, uncultivated Archaea (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2010. — May (vol. 107, no. 19). — P. 8806—8811. — doi:10.1073/pnas.0914470107. — . — PMID 20421484. — PMC 2889320.
- ↑ de Queiroz K. Ernst Mayr and the modern concept of species (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2005. — Vol. 102, no. Suppl 1. — P. 6600—6607. — doi:10.1073/pnas.0502030102. — . — PMID 15851674. — PMC 1131873. Архивировано 16 октября 2019 года.
- ↑ Eppley J. M., Tyson G. W., Getz W. M., Banfield J. F. Genetic exchange across a species boundary in the archaeal genus ferroplasma (англ.) // Genetics : journal. — 2007. — Vol. 177, no. 1. — P. 407—416. — doi:10.1534/genetics.107.072892. — PMID 17603112. — PMC 2013692. Архивировано 16 октября 2019 года.
- ↑ Papke R. T., Zhaxybayeva O., Feil E. J., Sommerfeld K., Muise D., Doolittle W. F. Searching for species in haloarchaea (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2007. — Vol. 104, no. 35. — P. 14092—14097. — doi:10.1073/pnas.0706358104. — . — PMID 17715057. — PMC 1955782. Архивировано 16 октября 2019 года.
- ↑ Kunin V., Goldovsky L., Darzentas N., Ouzounis C. A. The net of life: reconstructing the microbial phylogenetic network (англ.) // Genome Res. : journal. — 2005. — Vol. 15, no. 7. — P. 954—959. — doi:10.1101/gr.3666505. — PMID 15965028. — PMC 1172039. Архивировано 16 октября 2019 года.
- ↑ Hugenholtz P. Exploring prokaryotic diversity in the genomic era (англ.) // Genome Biol.[англ.]. — 2002. — Vol. 3, no. 2. — P. REVIEWS0003. — doi:10.1186/gb-2002-3-2-reviews0003. — PMID 11864374. — PMC 139013. Архивировано 15 марта 2020 года.
- ↑ Rappé M. S., Giovannoni S. J. The uncultured microbial majority (англ.) // Annu. Rev. Microbiol.. — 2003. — Vol. 57. — P. 369—394. — doi:10.1146/annurev.micro.57.030502.090759. — PMID 14527284.
- ↑ Морозова О. В. Загадки архей и их фагов // Вестник ВОГиС. — 2005. — Т. 9, № 1. — С. 55—66.
- ↑ Spang A., Hatzenpichler R., Brochier-Armanet C., Rattei T., Tischler P., Spieck E., Streit W., Stahl D. A., Wagner M., Schleper C. Distinct gene set in two different lineages of ammonia-oxidizing archaea supports the phylum Thaumarchaeota (англ.) // Trends Microbiol.[англ.] : journal. — 2010. — Vol. 18, no. 8. — P. 331—340. — PMID 20598889. Архивировано 2 июня 2013 года.
- ↑ 1 2 Spang A., Saw J. H., Jørgensen S. L., Zaremba-Niedzwiedzka K., Martijn J., Lind A. E., van Eijk R., Schleper C., Guy L., Ettema T. J. Complex archaea that bridge the gap between prokaryotes and eukaryotes. (англ.) // Nature. — 2015. — doi:10.1038/nature14447. — PMID 25945739.
- ↑ Seitz K. W., Lazar C. S., Hinrichs K. U., Teske A. P., Baker B. J. Genomic reconstruction of a novel, deeply branched sediment archaeal phylum with pathways for acetogenesis and sulfur reduction. (англ.) // The ISME journal. — 2016. — Vol. 10, no. 7. — P. 1696—1705. — doi:10.1038/ismej.2015.233. — PMID 26824177.
- ↑ Breithaupt H. The hunt for living gold. The search for organisms in extreme environments yields useful enzymes for industry (англ.) // EMBO Rep.[англ.] : journal. — 2001. — Vol. 2, no. 11. — P. 968—971. — doi:10.1093/embo-reports/kve238. — PMID 11713183. — PMC 1084137.
- ↑ 1 2 Egorova K., Antranikian G. Industrial relevance of thermophilic Archaea (англ.) // Curr. Opin. Microbiol.. — 2005. — Vol. 8, no. 6. — P. 649—655. — doi:10.1016/j.mib.2005.10.015. — PMID 16257257.
- ↑ Synowiecki J., Grzybowska B., Zdziebło A. Sources, properties and suitability of new thermostable enzymes in food processing (англ.) // Crit Rev Food Sci Nutr[англ.] : journal. — 2006. — Vol. 46, no. 3. — P. 197—205. — doi:10.1080/10408690590957296. — PMID 16527752.
- ↑ Jenney F. E., Adams M. W. The impact of extremophiles on structural genomics (and vice versa) (англ.) // Extremophiles : journal. — 2008. — January (vol. 12, no. 1). — P. 39—50. — doi:10.1007/s00792-007-0087-9. — PMID 17563834.
- ↑ Schiraldi C., Giuliano M., De Rosa M. Perspectives on biotechnological applications of archaea (англ.) // Archaea : journal. — 2002. — Vol. 1, no. 2. — P. 75—86. — doi:10.1155/2002/436561. — PMID 15803645. — PMC 2685559. Архивировано 26 августа 2013 года. Архивированная копия. Дата обращения: 21 июля 2012. Архивировано из оригинала 26 августа 2013 года.
- ↑ Norris P. R., Burton N. P., Foulis N. A. Acidophiles in bioreactor mineral processing (англ.) // Extremophiles. — 2000. — Vol. 4, no. 2. — P. 71—6. — doi:10.1007/s007920050139. — PMID 10805560.
- ↑ O'Connor E. M., Shand R. F. Halocins and sulfolobicins: the emerging story of archaeal protein and peptide antibiotics (англ.) // J. Ind. Microbiol. Biotechnol. : journal. — 2002. — January (vol. 28, no. 1). — P. 23—31. — doi:10.1038/sj/jim/7000190. — PMID 11938468.
- ↑ Shand R. F.; Leyva K. J. Archaeal Antimicrobials: An Undiscovered Country // Archaea: New Models for Prokaryotic Biology (англ.) / Blum P (ed.). — Caister Academic Press[англ.], 2008. — ISBN 978-1-904455-27-1.
Литература
- Воробьева Л. В. Археи: Учебное пособие для вузов. — М.: Академкнига, 2007. — 447 с.
- Громов Б. В. Удивительный мир архей // СОЖ. — 1997. — № 4. — С. 23—26.
- Морозова О. В. Загадки архей и их фагов // Вестник ВОГиС. — 2005. — Том 9. — № 1. — С. 55—66
- Thomas Cavalier-Smith. Cell evolution and Earth history: stasis and revolution. — 2006.
Ссылки
- Волкова, Ольга. Закинули археи эволюционный невод и вытянули... // Сайт Biomolecula.ru (22 января 2015). Дата обращения: 5 апреля 2018.
- Старокадомский, Пётр. Карл Вёзе (1928–2012). // Сайт Biomolecula.ru (7 февраля 2013). Дата обращения: 5 апреля 2018.
- Панов, Андрей. Как составлялся геном эукариот: эндосимбиоз VS. непрерывный горизонтальный перенос. // Сайт Biomolecula.ru (22 сентября 2015). Дата обращения: 5 апреля 2018.
- Конышев, Илья. Археи «хамят» и помогают. // Сайт Biomolecula.ru (3 сентября 2015). Дата обращения: 5 апреля 2018.
- Кондратенко, Юлия. Между бактериями и археями, окисляющими метан, обнаружена «электропроводка». // Сайт Biomolecula.ru (22 декабря 2015). Дата обращения: 5 апреля 2018.
| Таксономия | ||||
|---|---|---|---|---|