Бактерии
![Разнообразие бактерий. Сверху: Escherichia coli, Enterococcus sp., Treponema pallidum, Streptococcus pneumoniae; Посередине: Campylobacter jejuni[англ.], бактерия группы CPR[англ.], Oscillatoria[англ.] sp., представитель Atribacterota[англ.]; Снизу: Myxococcus xanthus, Streptomyces sp., Bacillus cereus, Vibrio vulnificus[англ.]](https://upload.wikimedia.org/wikipedia/commons/thumb/4/4f/Bacteria_collage.jpg/275px-Bacteria_collage.jpg)
Бакте́рии (лат. Bacteria, от др.-греч. βακτήριον — «палочка») — домен прокариотических микроорганизмов. Бактерии обычно достигают нескольких микрометров в длину, их клетки могут иметь разнообразную форму: от шарообразной до палочковидной и спиралевидной. Бактерии — одна из первых форм жизни на Земле и встречаются почти во всех земных местообитаниях. Они населяют почву, пресные и морские водоёмы, кислые горячие источники, радиоактивные отходы[2] и глубинные слои земной коры. Бактерии часто являются симбионтами и паразитами растений и животных. Большинство бактерий к настоящему времени не описано, и представители лишь половины отделов бактерий могут быть выращены в лаборатории[3]. Бактерии изучает наука бактериология — раздел микробиологии.
Один грамм почвы в среднем содержит 40 миллионов бактериальных клеток, а в миллилитре свежей воды можно найти миллион клеток бактерий. На Земле насчитывается около 5⋅1030 бактерий[4], и их биомасса превышает суммарную биомассу животных и растений[5]. Они играют важную роль в круговороте питательных веществ[англ.], например, именно бактерии осуществляют фиксацию атмосферного азота. Они также разлагают останки животных и растений посредством гниения[6]. Экстремофильные бактерии, обитающие рядом с холодными[англ.] и горячими гидротермальными источниками, вырабатывают энергию из нерастворимых соединений, таких как сероводород и метан. Предполагается, что бактерии живут и в Марианской впадине, имеющей глубину 11 километров[7][8]. Имеются сообщения о бактериях, обитающих в каменистых породах на 580 метров глубже морского дна на глубине 2,6 км около северо-востока США[7][9].
Человеческую микрофлору составляют 39 триллионов бактериальных клеток (само тело человека состоит из около 30 триллионов клеток)[10]. Наиболее многочисленна кишечная микрофлора. Кожа также заселена многими бактериями[11]. Большинство бактерий, обитающих в человеческом теле, безвредно за счёт сдерживающего действия иммунной системы или приносят пользу (микрофлора человека). Ряд бактерий патогенен для человека. Такие инфекционные болезни, как холера, сифилис, сибирская язва, проказа и бубонная чума, вызываются бактериями. Наибольшее число смертей вызвано бактериальными респираторными инфекциями[англ.], и один лишь туберкулёз ежегодно убивает 2 миллиона человек (преимущественно в Африке южнее Сахары)[12]. В развитых странах антибиотики используются не только для лечения заболеваний человека, но и в животноводстве, из-за чего проблема устойчивости к антибиотикам становится всё более актуальной. В промышленности бактерии используют в очистке сточных вод, для ликвидации разливов нефти, при получении сыра и йогурта, восстановлении золота, палладия, меди и других металлов из руд[13], а также в биотехнологии, для получения антибиотиков и других соединений[14].
Первоначально бактерии поместили в царство растений в составе класса Schizomycetes. Сейчас известно, что бактерии, в отличие от растений и других эукариот, не имеют оформленного ядра и, как правило, мембранных органелл. Традиционно бактериями называли всех прокариот, однако в 1970-х годах было показано, что прокариоты представлены двумя независимыми доменами — бактериями и археями (эукариоты составляют третий домен)[15].
Этимология
Слово «бактерия» происходит от лат. bacterium, производного от др.-греч. βακτήριον, что означает «палочка», так как первые описанные бактерии были палочковидными[16][17].
Происхождение и ранняя эволюция
Предки современных бактерий были одноклеточными микроорганизмами, которые стали одной из первых форм жизни на Земле, появившись около 4 миллиардов лет назад. Почти три миллиарда лет вся жизнь на Земле была микроскопической[18][19]. Хотя для бактерий известны ископаемые останки (например, строматолиты), их морфология очень однообразна, что не позволяет идентифицировать отдельные виды. Однако для реконструкции филогении бактерий можно использовать последовательности генов, и именно с их помощью было показано, что бактерии отделились раньше архей и эукариот[20]. Ближайший общий предок бактерий и архей, вероятнее всего, был гипертермофилом, который жил 3—2,5 млрд лет назад[21][22].
Бактерии сыграли важнейшую роль в появлении эукариот. Считается, что эукариотическая клетка возникла, когда бактерии стали эндосимбионтами одноклеточных организмов, вероятно, близких к современным археям[23][24]. Иными словами, прото-эукариотическая клетка проглотила клетку α-протеобактерии, которая дала начало митохондриям и гидрогеносомам. На данный момент неизвестны эукариоты, лишённые и митохондрий, и гидрогеносом, хотя иногда эти органеллы сильно редуцированы. Впоследствии некоторые из эукариот, уже имеющих митохондрии, проглотили клетки цианобактерий, которые стали пластидами растений и водорослей[25][26].
Морфология

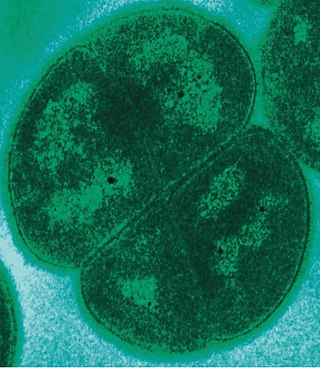
Бактериальные клетки имеют чрезвычайно разнообразную морфологию (то есть форму и размер). Как правило, бактериальные клетки в десять раз мельче эукариотических и достигают 0,5—5 мкм в длину. При этом есть и бактерии, видимые невооружённым глазом: так, Thiomargarita namibiensis достигает половины миллиметра в длину[27], а длина Epulopiscium fishelsoni может составлять 0,7 мм[28]. Thiomargarita magnifica вырастает до 2 см в длину, отличается необычно сложной клеткой, фактически стирая границу между прокариотами и эукариотами и, по одному из предположений, даже является недостающим звеном в эволюции живого от одноклеточных организмов[29]. К числу самых мелких бактерий можно отнести представителей рода Mycoplasma, длина клеток которых не превышает 0,3 мкм, что сравнимо по размерам с вирионами некоторых вирусов[30] и лишь немного превышает предел разрешающей способности оптического микроскопа. Существуют ещё более мелкие бактерии (ультрамикробактерии[англ.]), однако они плохо изучены[31].
Большинство бактерий имеют шарообразную (кокки) или палочковидную (бациллы) форму[32]. Некоторые бактерии, называемые вибрионами[англ.], выглядят как слегка закрученные палочки или запятые; спириллы имеют спиральную форму, а спирохеты имеют длинные плотно закрученные клетки. Описаны и бактерии с другими необычными формами клеток, например, клетками в форме звезды[33]. Разнообразие форм бактериальных клеток обусловлено особенностями их клеточных стенок и цитоскелета. Форма бактериальной клетки обусловливает их способность поглощать питательные вещества, прикрепляться к поверхностям, плавать в жидкостях и ускользать от питающихся бактериями организмов[34].

Многие виды бактерий существуют в виде одиночных клеток, однако у некоторых видов клетки образуют характерные скопления: например, клетки Neisseria объединены в пары, у Streptococcus — в цепочки, у Staphylococcus — в скопления в виде грозди винограда. Некоторые бактерии могут формировать более сложные многоклеточные структуры. Так, Actinobacteria формируют длинные филаменты (внутриклеточные нитевидные образования), Myxococcales образуют плодовые тела, а Streptomyces образуют ветвящиеся нити[35]. Иногда такие сложные структуры появляются только при некоторых условиях. Например, при нехватке аминокислот клетки Myxococcales определяют расположение соседних клеток того же вида с помощью чувства кворума, движутся навстречу друг другу и формируют плодовые тела до 500 мкм длиной, состоящие из около 100 тысяч бактериальных клеток[36]. Бактериальные клетки в составе плодовых тел выполняют различные функции: десятая часть всех клеток мигрирует к верхней части плодового тела и превращается в особую покоящуюся форму, называемую миксоспорой, которая более устойчива к высыханию и другим неблагоприятным условиям внешней среды[37].
Бактерии часто прикрепляются к какой-либо поверхности и формируют плотные скопления, известные как биоплёнки, или более крупные скопления — бактериальные маты. Толщина биоплёнок и матов может составлять от нескольких микрометров до полуметра, в их состав могут входить бактерии разных видов, а также археи и протисты. В биоплёнках наблюдается сложное расположение клеток и внеклеточных компонентов, которые формируют вторичные структуры, известные как микроколонии, через которые проходит сеть каналов, обеспечивающая лучшую диффузию питательных веществ[38][39]. В таких местообитаниях, как почва и поверхность растений, большинство бактерий, прикреплённых к поверхностям, входят в состав биоплёнок[40]. Биоплёнки имеют важное значение для медицины, потому что они часто образуются при хронических бактериальных инфекциях или инфекциях, связанных с инородными имплантатами. Более того, бактерии в составе биоплёнок гораздо сложнее убить, чем отдельные бактериальные клетки[41].
Строение клетки

Внутриклеточные структуры
Бактериальная клетка окружена мембраной, состоящей в основном из фосфолипидов. Мембрана окружает всё содержимое клетки и выступает в роли барьера для удержания в клетке питательных веществ, белков и других компонентов цитоплазмы[42]. В отличие от клеток эукариот, у бактерий, как правило, отсутствуют крупные мембранные органеллы, такие как ядро, митохондрии, хлоропласты[43]. Однако у некоторых бактерий имеются органеллы с белковой оболочкой, в которых протекают определённые метаболические процессы[44][45], например, карбоксисомы[46]. Кроме того, у бактерий имеется многокомпонентный цитоскелет, который контролирует локализацию нуклеиновых кислот и белков внутри клетки и управляет клеточным делением[47][48][49].
Многие важные биохимические реакции, такие как образование АТФ, происходят за счёт градиента концентрации определённых ионов по разные стороны мембраны, что создаёт разность потенциалов, как в батарейке. Поскольку у бактерий нет мембранных органелл, такие реакции (например, перенос электронов) протекают при участии мембраны бактериальной клетки, обращённой во внешнюю среду в случае грамположительных бактерий или в периплазматическое пространство в случае грамотрицательных бактерий[50]. Однако у многих фотосинтезирующих бактерий мембрана образует многочисленные складки, которые заполняют почти всё внутреннее пространство клетки[51]. На этих складках располагаются светопоглощающие комплексы, однако у некоторых бактерий, например, зелёных серных бактерий, светопоглощающие комплексы находятся внутри особых мембранных пузырьков — хлоросом[52].
У большинства бактерий нет ядра, окружённого мембранами, и их генетический материал, в большинстве случаев представленный единственной кольцевой молекулой ДНК[англ.], находится в цитоплазме в составе нуклеоида, имеющего неправильную форму[53]. Нуклеоид содержит не только геномную ДНК, но также взаимодействующие с ней белки и РНК. Как все живые организмы, бактерии имеют рибосомы, которые обеспечивают синтез белков, однако размеры и структура рибосом бактерий отличаются от таковой у рибосом архей и эукариот[54].
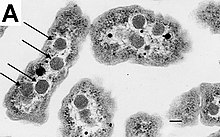
У некоторых бактерий в цитоплазме имеются гранулы, запасающие питательные вещества, такие как гликоген[55], полифосфат[56], сера[57] или полигидроксиалканоаты[58]. Ряд бактерий, например, фотосинтезирующие цианобактерии, имеют газовые вакуоли, с помощью которых они регулируют свою плавучесть, благодаря чему могут перемещаться между слоями воды с разным содержанием питательных веществ и уровнем освещённости[59].
Внеклеточные структуры
Поверх мембраны бактериальной клетки располагается клеточная стенка. Клеточная стенка бактерий состоит из пептидогликана, также известного как муреин, который состоит из полисахаридных цепочек, связанных пептидными линкерами из D-аминокислот[60]. По химическому составу бактериальная клеточная стенка отличается от клеточной стенки растений и грибов, у которых она состоит из целлюлозы и хитина соответственно[61]. Клеточная стенка архей также не содержит пептидогликана. Клеточная стенка жизненно необходима для многих видов бактерий, и некоторые антибиотики, такие как пенициллин, подавляют биосинтез пептидогликана и тем самым убивают бактерию[61].
В широком смысле по составу клеточной стенки бактерий принято делить на грамположительные и грамотрицательные. Название этих типов связано с их дифференциальной окраской по методу Грама, который долгое время используется для классификации бактерий[62]. У грамположительных бактерий имеется толстая клеточная стенка, состоящая из многих слоёв пептидогликана и тейхоевых кислот. У грамотрицательных бактерий, напротив, клеточная стенка значительно тоньше и включает всего лишь несколько слоёв пептидогликана, а поверх неё залегает вторая мембрана, содержащая липополисахариды и липопротеины. Большинство бактерий грамотрицательны, и только фирмикуты и актинобактерии грамположительны (ранее они были известны как грамположительные бактерии с низким GC-составом и грамположительные бактерии с высоким GC-составом соответственно)[63]. Различия между грамположительными и грамотрицательными бактериями могут обусловливать различную чувствительность к антибиотикам. Например, ванкомицин эффективен только против грамположительных бактерий и не действует на грамотрицательные бактерии[64]. У некоторых бактерий строение клеточной стенки не соответствует в строгом смысле ни грамположительному, ни грамотрицательному типу. Например, у микобактерий имеется толстый слой пептидогликана, как у грамположительных бактерий, который покрыт внешней мембраной, как у грамотрицательных бактерий[65].
У многих бактерий клетка покрыта так называемым S-слоем, состоящим из плотно уложенных молекул белков[66]. S-слой обеспечивает химическую и физическую защиту клетки и может выступать в роли макромолекулярного диффузионного барьера. Функции S-слоя разнообразны, но плохо изучены, однако известно, что у Campylobacter он выступает фактором вирулентности, а у Geobacillus stearothermophilus[англ.] он содержит поверхностные ферменты[67].
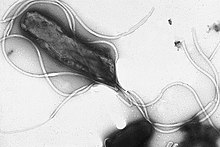
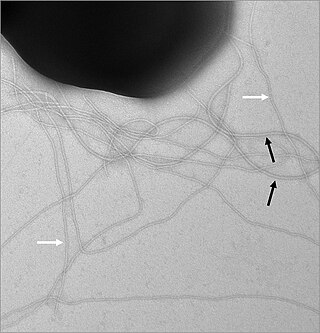
У многих бактерий имеются жгутики, представляющие собой плотные белковые структуры около 20 нм в диаметре и до 20 мкм в длину. Они обеспечивают подвижность клеток и по строению и механизму работы не имеют ничего общего с эукариотическими жгутиками. Движение жгутиков бактерий происходит за счёт энергии, которая высвобождается при движении ионов по электрохимическому градиенту через клеточную мембрану[68].
Нередко клетки бактерий покрыты фимбриями, которые представляют собой белковые филаменты, достигающие 2—10 нм в диаметре и до нескольких мкм в длину. Они покрывают всю поверхность бактериальной клетки и в электронный микроскоп выглядят как волоски. Предполагается, что фимбрии участвуют в прикреплении клеток бактерий к различным поверхностям и друг к другу, а у многих патогенных бактерий они являются факторами вирулентности[69]. Пили — это клеточные белковые придатки, более толстые, чем фимбрии, которые обеспечивают перенос генетического материала от одной бактериальной клетки к другой в ходе конъюгации (половые пили)[70]. Кроме того, пили IV типа участвуют в движении[71].
Многие бактериальные клетки выделяют покрывающий их гликокаликс различной сложности строения: от тонкого неструктурированного слоя внеклеточных полимеров[англ.] до высоко структурированной капсулы. Гликокаликс может защищать бактерию от поглощения эукариотическими клетками, например, макрофагами, входящими в состав иммунной системы[72]. Он также может выступать в роли антигена, который используется для распознавания бактериальных клеток иммунной системой, а также участвовать в формировании биоплёнок и прикреплении бактериальных клеток к поверхностям[73].
Образование внеклеточных структур бактериальной клетки обеспечивается бактериальными системами секреции. Они транспортируют белки из цитоплазмы в периплазматическое пространство или во внешнюю среду. Известно несколько типов бактериальных систем секреции, кроме того, бактериальные системы секреции нередко выступают в роли факторов вирулентности[74].
Эндоспоры
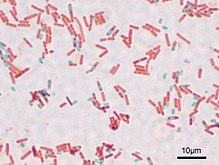
Представители нескольких родов грамположительных бактерий, таких как Bacillus, Clostridium, Sporohalobacter[англ.], Anaerobacter[англ.] и Heliobacterium, образуют покоящиеся структуры, обладающие повышенной устойчивостью к неблагоприятным факторам внешней среды и называемые эндоспорами[75]. Эндоспоры образуются в цитоплазме клетки, и, как правило, в одной клетке может сформироваться только одна эндоспора. Каждая эндоспора содержит ДНК и рибосомы, окружённые поверхностным слоем цитоплазмы, поверх которого залегает плотная многослойная оболочка, состоящая из пептидогликана и разнообразных белков[76].
Внутри эндоспор не протекают метаболические процессы, и они могут выживать при сильнейших неблагоприятных физических и химических воздействиях, таких как интенсивное УФ-излучение, γ-излучение, детергенты, дезинфицирующие агенты, замораживание, давление и высыхание[англ.][77]. Эндоспоры могут сохранять жизнеспособность в течение миллионов лет[78][79], и с их помощью бактерии могут оставаться живыми даже в условиях вакуума и космического излучения[80]. Некоторые бактерии, формирующие эндоспоры, патогенны. Так, сибирская язва развивается после вдыхания спор грамположительной бактерии Bacillus anthracis, а попадание эндоспор Clostridium tetani в глубокие открытые раны может привести к столбняку[81].
Метаболизм
У бактерий наблюдается колоссальное разнообразие видов метаболизма[82]. Традиционно таксономия бактерий строилась на основе их метаболических особенностей, однако она во многом не совпадает с современной классификацией, построенной на геномных последовательностях[83]. Бактерии делятся на три типа питания в зависимости от ключевых черт метаболизма: источника энергии, донора электронов и источника углерода[84].
Бактерии получают энергию двумя способами: поглощая свет в ходе фотосинтеза или окисляя химические соединения (хемосинтез)[85]. Хемотрофы используют в качестве источника энергии химические вещества, перенося электроны с имеющегося донора на конечный акцептор электронов в ходе окислительно-восстановительной реакции. Высвобождающаяся при этой реакции энергия далее используется для нужд метаболизма. В зависимости от того, какое вещество используется как донор электронов, хемотрофы подразделяются ещё на несколько групп. Бактерии, использующие неорганические вещества, такие как водород, угарный газ или аммиак, называются литотрофами, а бактерии, окисляющие органические соединения, называются органотрофами. Бактерий также классифицируют в зависимости от веществ, выступающих акцепторами электронов. У аэробов акцептором электронов выступает кислород, а анаэробы используют для этого другие соединения, такие как нитрат, сульфат и углекислый газ[85].
Многие бактерии удовлетворяют свои потребности в углероде за счёт органических соединений; такие бактерии называются гетеротрофами. Другие бактерии, например, цианобактерии и некоторые пурпурные бактерии, являются автотрофами, то есть получают углерод, фиксируя углекислый газ[86]. В некоторых условиях метанотрофные бактерии используют метан и как источник электронов, и как источник углерода[87].
| Тип питания | Источник энергии | Источник углерода | Примеры |
|---|---|---|---|
| Фототрофы | Солнечный свет | Органические вещества (фотогетеротрофы) или фиксированный углекислый газ (фотоавтотрофы) | Цианобактерии, зелёные серные бактерии, Chloroflexi[англ.], пурпурные бактерии |
| Литотрофы | Неорганические соединения | Органические вещества (литогетеротрофы) или фиксированный углекислый газ (литоавтотрофы) | Thermodesulfobacteria[англ.], Hydrogenophilaceae[англ.], Nitrospirae[англ.] |
| Органотрофы | Органические соединения | Органические вещества (хемогетеротрофы) или фиксированный углекислый газ (хемоавтотрофы) | Bacillus, Clostridium, Enterobacteriaceae |
Метаболизм бактерий имеет огромное значение для экологической стабильности и деятельности человека. Например, некоторые бактерии являются единственными фиксаторами атмосферного азота (с помощью фермента нитрогеназы)[88]. Другими важными для окружающей среды химическими процессами, осуществляемыми бактериями, являются денитрификация, восстановление сульфата и ацетогенез[89][90]. Метаболические процессы бактерий также могут служить источниками загрязнения. Так, сульфатредуцирующие бактерии образуют высокотоксичные соединения ртути (метил- и диметилртуть)[91]. Ряд анаэробных бактерий осуществляет брожение для получения энергии, и его побочные продукты (например, этанол при спиртовом брожении) попадают в окружающую среду. Факультативные анаэробы могут переключаться между получением энергии с помощью брожения и получением её с помощью дыхания с различными акцепторами электронов в зависимости от условий окружающей среды[92].
Рост и размножение

В отличие от многоклеточных организмов, у одноклеточных организмов, в том числе бактерий, увеличение клетки в размерах и размножение путём деления клеток тесно связаны[93]. Бактериальные клетки достигают определённого размера и после этого делятся бинарным делением. В оптимальных условиях бактерии растут и делятся очень быстро, описан пример морской псевдомонады, популяция которой может удваиваться каждые 9,8 минуты[94]. При бинарном делении образуются две дочерние клетки, идентичные материнской. Некоторые бактерии, хотя и размножаются простым делением, образуют более сложные структуры, предназначенные для распространения дочерних клеток. Примером могут служить плодовые тела миксобактерий и воздушные гифы стрептомицетов. Некоторые бактерии способны к почкованию, когда дочерняя клетка образует вырост на материнской, который впоследствии отделяется и переходит к самостоятельной жизни[95].

В лаборатории бактерий выращивают на твёрдых или жидких средах. Твёрдые среды, такие как агар, используются для изоляции[англ.] чистых культур бактериальных штаммов. Жидкие среды используются, когда необходимо измерять скорость роста или получить большое количество клеток. При выращивании бактерий в жидкой среде с перемешиванием получаются однородные клеточные культуры, однако сложно заметить загрязнение другими бактериями. Для идентификации отдельных бактерий используются селективные среды, содержащие антибиотики, специфические питательные вещества или, наоборот, лишённые каких-то соединений[97].
Для большинства лабораторных методов выращивания бактерий необходимы большие количества питательных веществ, чтобы обеспечить быстрое получение больших объёмов клеток. Однако в естественных условиях питательные вещества ограничены, и бактерии не могут размножаться бесконечно. Из-за ограниченного количества питательных веществ в ходе эволюции появились различные стратегии роста. Некоторые виды растут чрезвычайно быстро, когда питательные вещества доступны, например, цианобактерии нередко вызывают цветение водоёмов, насыщенных органикой[98]. Другие организмы адаптированы к жёстким условиям окружающей среды, например, бактерии рода Streptomyces выделяют антибиотики, которые подавляют рост конкурирующих бактерий[99]. В природе многие виды бактерий живут сообществами (например, в виде биоплёнок), которые обеспечивают каждую клетку необходимым питанием и защищают от неблагоприятных условий[40]. Некоторые организмы и группы организмов растут только в составе сообществ и не могут быть выделены в чистую культуру[100].
Динамику роста бактериальной популяции можно подразделить на четыре фазы. Когда популяция бактерий попадает в среду, богатую питательными веществами, клетки начинают адаптироваться к новым условиям. Первая фаза роста называется лаг-фазой, это период медленного роста, когда клетки адаптируются к среде, богатой питательными веществами, и готовятся к быстрому росту. Во время лаг-фазы происходит интенсивный синтез белков[101]. За лаг-фазой следует логарифмическая, или экспоненциальная фаза, во время которой происходит быстрый экспоненциальный рост. Скорость, с которой клетки растут во время этой фазы, называют скоростью роста, а время, которое необходимо для удвоения клеточной популяции, называется временем генерации. В ходе лог-фазы питательные вещества потребляются с максимальной скоростью, до тех пор пока одно из необходимых соединений не кончится и не начнёт подавлять рост. Третья фаза роста называется стационарной, она начинается при нехватке питательных веществ для быстрого роста. Скорость метаболизма падает, и клетки начинают расщеплять белки, не являющиеся строго необходимыми. Во время стационарной фазы экспрессируются гены, белковые продукты которых участвуют в репарации ДНК, метаболизме антиоксидантов и транспорте питательных веществ[102]. Финальная фаза роста — фаза смерти, при которой запас питательных веществ исчерпывается и бактерии погибают[103].
Генетика
У большинства бактерий геном представлен единственной кольцевой молекулой ДНК (её иногда называют хромосомой), а размер генома варьирует от 160 тысяч пар оснований (п. о.) у эндосимбиотической бактерии Carsonella ruddii[англ.][104] до примерно 13 миллионов п. о. у почвенной бактерии Sorangium cellulosum[англ.][105]. Впрочем, у ряда представителей родов Streptomyces и Borrelia геном представлен единственной линейной хромосомой[106][107], а у некоторых видов рода Vibrio имеется более одной хромосомы[108]. Многие бактерии также содержат плазмиды — маленькие внехромосомные молекулы ДНК, которые содержат несколько генов, обеспечивающих своих обладателей разными полезными свойствами: устойчивостью к антибиотикам, новыми метаболическими реакциями и разнообразными факторами вирулентности[109].
В бактериальных геномах, как правило, содержится от нескольких сотен до нескольких тысяч генов. Бактериальные гены, за редкими исключениями, лишены интронов, а если они и есть, то очень короткие[110].
При делении бактериальной клетки дочерние клетки наследуют идентичные копии её генома и, по сути, являются её клонами. Однако в геномах бактерий постоянно происходят мутации, лучшие из которых поддерживаются отбором, кроме того, иногда у бактерий происходит и генетическая рекомбинация. Мутации возникают из-за ошибок ферментов, удваивающих ДНК, а также под действием мутагенов. Скорость мутирования значительно различается не только у бактерий разных видов, но даже и у разных клонов, относящихся к одному виду[111]. Изменения в бактериальных геномах происходят также благодаря случайным мутациям и так называемым стресс-направленным мутациям (то есть гены, отвечающие за какой-либо процесс, сдерживающий рост, мутируют особенно часто)[112].
У некоторых бактерий клетки могут обмениваться друг с другом генетическим материалом. Существует три способа обмена генетической информацией между бактериями. Во-первых, бактериальные клетки могут поглощать экзогенную ДНК из окружающей среды в ходе процесса, называемого трансформацией[113]. Некоторые бактерии поглощают ДНК извне в нормальном состоянии, а другие начинают захватывать чужеродную ДНК после химического воздействия, то есть они должны прежде стать компетентными[англ.][114]. В природе компетентность развивается при стрессовых условиях и выступает в роли адаптивного механизма, потому что захваченный извне фрагмент ДНК может оказаться полезным для переживания неблагоприятных условий[115]. Во-вторых, бактерия может получить чужеродную ДНК в ходе трансдукции, когда бактериофаг не только сам вставляется в бактериальную хромосому, но и приносит с собой фрагмент генома другой бактерии. Существует множество бактериофагов, но для всех них существует два типа жизненного цикла: литический цикл, когда клетка-хозяин разрушается вскоре после заражения, высвобождая наружу новые вирусные частицы, и лизогенный цикл, когда бактериофаг встраивается в геном бактерии и до некоторого момента мирно сосуществует с ней[116]. У бактерий существует ряд механизмов защиты от бактериофагов, в частности, система рестрикции-модификации[117] и система CRISPR/Cas. Система CRISPR/Cas, по сути, играет роль адаптивного иммунитета, потому что фрагменты геномов фагов, заразивших клетку, встраиваются в локус CRISPR, и при повторном заражении их репликация подавляется за счёт РНК-интерференции[118][119]. В-третьих, бактериальные клетки обмениваются генетическим материалом в ходе конъюгации, когда ДНК передаётся из одной клетки в другую при непосредственном контакте. Обычно передача генетического материала любым из трёх механизмов подразумевает участие бактерий одного вида, однако в некоторых случаях ДНК обмениваются бактерии разных видов. Например, гены устойчивости к антибиотикам передаются от одних видов бактерий к другим[120][121]. Передача генетического материала между разными особями носит название горизонтального переноса генов (в противовес вертикальному переносу генов, то есть от родителей к потомкам)[122].
Поведение
Движение

Многие бактерии подвижны и перемещаются за счёт разных механизмов. Чаще всего для движения используются жгутики — длинные филаменты, которые вращаются, подобно пропеллеру[123], за счёт особого мотора у их основания. Движущей силой для мотора является электрохимический градиент клеточной мембраны[124]. В состав бактериального жгутика входят около 20 белков, а ещё 30 белков необходимы для регуляции его работы и сборки[123].

С помощью жгутиков бактерии могут двигаться по-разному. Многие бактерии, например, кишечная палочка Escherichia coli, могут двигаться вперёд или кувыркаться. За счёт кувыркания клетка меняет направление своего движения, которое представляет собой случайное блуждание в трёхмерном пространстве[125]. Количество и расположение жгутиков различно у разных видов. У некоторых бактерий есть всего один жгутик (монотрихи), у других два жгутика располагаются на двух противоположных концах клетки (амфитрихи), у третьих на полюсах клетки находятся пучки жгутиков (лофотрихи), а у четвёртых жгутики покрывают всю поверхность клетки (перитрихи). У спирохет жгутик находится в периплазматическом пространстве между двумя мембранами. Клетки спирохет имеют характерную извитую форму, которая меняется при движении[123].
Некоторые бактерии способны к так называемым подёргиваниям[англ.] (англ. twitching) за счёт пилей IV типа[126], а также скольжению. При подёргивании на клетке имеется палочковидный пиль, который связывается с субстратом и сокращается, проталкивая клетку вперёд[127].
Для подвижных бактерий характерно движение, направленное навстречу какому-либо стимулу или, напротив, от него — таксис. К числу таких поведенческих программ относятся хемотаксис, фототаксис, энергетический таксис и магнитотаксис[128][129][130]. Клетки бактерий могут кооперироваться с образованием единого скопления за счёт чувства кворума, как, например, миксобактерии при образовании плодовых тел. Некоторые виды родов внутриклеточных паразитов[англ.] Listeria и Shigella движутся внутри клетки-хозяина, используя её цитоскелет, который обычно используется для перемещения клеточных органелл. Стимулируя полимеризацию актина у одного из полюсов своих клеток, эти бактерии формируют своего рода актиновый хвост, который проталкивает их вперёд[131].
Коммуникация
У некоторых бактерий имеются химические системы, испускающие свет. Способность к биолюминесценции часто имеется у бактерий, живущих в симбиозе с глубоководными рыбами, и свет, производимый бактериями, привлекает рыб друг к другу или более крупных животных к рыбам[132].
Бактерии часто формируют многоклеточные скопления, известные как биоплёнки, обмениваясь разнообразными химическими сигналами, за счёт которых их движение становится координированным[133][134]. Формирование многоклеточных скоплений даёт бактериям ряд преимуществ: в них наблюдаются разделение труда между клетками и появление различных функциональных типов клеток, питательные вещества усваиваются более эффективно, обеспечивается более надёжная защита от естественных врагов. Например, бактерии в составе биоплёнок в 500 раз более устойчивы к антибиотикам, чем одиночные планктонные клетки того же вида[134].
Координированное поведение клеток одного и того же вида бактерий часто осуществляется за счёт особых химических веществ. На основе локальной концентрации этих веществ бактерия определяет плотность клеток-сородичей вокруг себя (чувство кворума). За счёт чувства кворума бактерии могут координировать экспрессию генов и начинают выделять и улавливать аутоиндукторы[англ.] или феромоны, концентрация которых повышается по мере роста популяции[135].
Классификация и идентификация
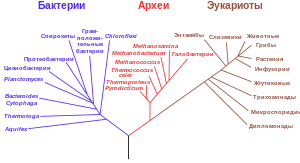
Бактерий можно классифицировать на основе строения клетки, метаболизма, а также различий в химическом составе клеток (наличия или отсутствия некоторых жирных кислот, пигментов, антигенов, хинонов)[97]. В то время как перечисленные характеристики подходят для выделения штаммов, непонятно, можно ли их использовать для разделения видов бактерий. Дело в том, что у большинства бактерий нет отличительных структур, а из-за широко распространённого горизонтального переноса генов родственные виды могут сильно отличаться по морфологии и метаболизму[136]. В связи с этим в настоящее время современная классификация базируется на молекулярной филогенетике. К числу её методов относят определение GC-состава генома, гибридизация геномов, а также секвенирование генов, которые не подверглись интенсивному горизонтальному переносу, такие как гены рРНК[137]. Релевантная классификация бактерий публикуется «Международным журналом систематической бактериологии» (англ. International Journal of Systematic Bacteriology)[138] и руководством по систематической бактериологии Берджи (англ. Bergey's Manual of Systematic Bacteriology). Международный комитет систематики прокариот[англ.] (англ. International Committee on Systematics of Prokaryotes) регулирует международные правила именования таксонов бактерий и определение их рангов согласно правилам Международного кодекса номенклатуры прокариот[англ.] (англ. International Code of Nomenclature of Prokaryotes)[139].
Термин «бактерии» традиционно применяли по отношению к микроскопическим одноклеточным прокариотам. Однако данные молекулярной филогенетики свидетельствуют о том, что в действительности прокариоты подразделяются на два независимых домена, которые первоначально получили названия эубактерии (лат. Eubacteria) и архебактерии (лат. Archaebacteria), но в настоящее время называются бактерии и археи[15]. Эти два домена, наряду с доменом эукариоты, составляют основу трёхдоменной системы, которая является наиболее популярной системой классификации живых организмов[140]. Археи и эукариоты состоят в более близком родстве, чем каждый из этих доменов к бактериям. Впрочем, высказывается мнение, что археи и эукариоты произошли от грамположительных бактерий[141]. Поскольку количество отсеквенированных последовательностей бактериальных геномов очень быстро растёт, классификация бактерий постоянно меняется[3][142].
В медицине идентификация бактерий имеет огромное значение, поскольку от неё зависит схема лечения. По этой причине ещё до эры молекулярной биологии учёные активно разрабатывали методы, позволяющие быстро идентифицировать патогенные бактерии. В 1884 году Ганс Кристиан Грам предложил метод дифференциального окрашивания бактерий на основе строения их клеточной стенки[62]. При окрашивании по Граму грамположительные бактерии с толстым слоем пептидогликана имеют фиолетовый цвет, а грамотрицательные бактерии с тонким слоем пептидогликана окрашены в розовый. Комбинируя окрашивание по Граму и морфотипы, выделяют четыре основные группы бактерий: грамположительные кокки, грамположительные бациллы, грамотрицательные кокки, грамотрицательные бациллы. Однако для идентификации некоторых бактерий больше подходят другие методы окрашивания. Например, микобактерии и бактерии рода Nocardia не обесцвечиваются кислотами[англ.] после окрашивания по Цилю — Нильсену[143]. Некоторых бактерий можно идентифицировать по их росту на специфических средах и при помощи других методов, например, серологии[144].
Методы культивирования бактерий[англ.] разработаны так, чтобы способствовать росту определённых бактерий, но подавлять рост других бактерий из того же образца. Часто эти методы разрабатываются специально для определённых образцов, откуда берутся микробы. Например, для идентификации возбудителя пневмонии для дальнейшего культивирования берут образец мокроты, для идентификации возбудителя диареи для выращивания на селективной среде берут образец стула, причём во всех случаях рост непатогенных бактерий будет подавляться. Образцы, которые в норме стерильны (например, кровь, моча, спинномозговая жидкость), культивируются в условиях, подходящих для роста любых микроорганизмов[97][145]. После изоляции патогенного микроорганизма можно изучать его морфологию, особенности роста (например, аэробный или анаэробный рост), характер гемолиза[англ.], а также окрашивать его разными методами.
Как и для классификации бактерий, молекулярные методы всё чаще применяют и для их идентификации. Диагностика, использующая такие молекулярные методы, как полимеразная цепная реакция (ПЦР), набирает всё большую популярность благодаря своей скорости и специфичности[146]. С помощью этих методов можно обнаруживать и идентифицировать бактерии, которые, хотя и сохраняют метаболическую активность, не делятся и поэтому не могут быть выращены в культуре[147]. Однако даже с помощью молекулярных методов точно определить или хотя бы примерно оценить число существующих видов бактерий невозможно. По состоянию на 2018, год описано несколько тысяч видов бактерий, но лишь около 250 из них являются патогенами человека[148]. Общее число видов бактерий, по разным оценкам, составляет от 107 до 109, но даже эти оценки могут быть на порядки меньше настоящего количества видов[149][150].
Однозначная и точная концепция вида бактерий так и не сформулирована. Это связано с невероятным разнообразием бактерий, широким распространением горизонтального переноса генов, невозможностью культивирования большинства бактерий и рядом других причин. Введение ПЦР и методов секвенирования в микробиологию позволило выделять виды бактерий на основании степени их сходства с геномами уже известных бактерий, однако и этот подход зачастую оказывается неэффективен из-за огромного разнообразия бактерий[151].
Помимо видов, при классификации бактерий иногда используют другие категории. К названию не до конца подтверждённых, а только предполагаемых видов добавляют слово Candidatus[152]. Многие виды подразделяются на так называемые штаммы — морфологические или генетические варианты (подтипы) бактерий в пределах одного вида. Однако ряд специалистов считает категорию «штамм» искусственной[153].
Взаимодействия с другими организмами

Несмотря на видимую простоту, бактерии могут вступать в сложные взаимоотношения с другими организмами. Такие симбиотические отношения можно подразделить на паразитизм, мутуализм и комменсализм, а также хищничество. Из-за небольших размеров бактерии-комменсалы распространены повсеместно и обитают на всевозможных поверхностях, в том числе на растениях и животных. Рост бактерий на теле человека ускоряется от тепла и пота, и их большие популяции придают запах телу[англ.].
Хищники
Некоторые бактерии убивают и поглощают другие микроорганизмы. К числу таких хищных бактерий[156] относится Myxococcus xanthus, формирующая скопления, которые убивают и переваривают любую попавшую на них бактерию[157]. Хищная бактерия Vampirovibrio chlorellavorus[англ.] прикрепляется к своей добыче, после чего постепенно переваривает её и всасывает высвобождающиеся питательные вещества[158]. Daptobacter проникает внутрь других бактериальных клеток и размножается в их цитозоле[159]. Вероятно, хищные бактерии произошли от сапрофагов, питающихся мёртвыми микроорганизмами, после того как приобрели приспособления для ловли и убийства других микробов[160].
Мутуалисты
Некоторые виды бактерий образуют скопления, которые необходимы для их выживания. Одна из таких мутуалистических ассоциаций, известная как межвидовая передача водорода, формируется между кластерами анаэробных бактерий, которые поглощают органические кислоты, такие как масляная и пропионовая кислоты, и выделяют водород, и метаногенными археями, которые используют водород. Бактерии из этой ассоциации не могут поглощать органические кислоты сами по себе, так как в ходе этой реакции образуется водород, накапливающийся вокруг. Только благодаря метаногенным археям концентрация водорода поддерживается достаточно низкой, чтобы позволить бактериям расти[161].
Многие бактерии являются симбионтами людей и других организмов. У человека от бактерий полностью свободны только кровь и лимфа[162]. Например, более тысячи видов бактерий, входящих в состав нормальной кишечной микрофлоры человека, участвуют в работе иммунитета, синтезируют витамины (например, фолиевую кислоту, витамин K и биотин), превращают сахара в молочную кислоту, а также сбраживают сложные неперевариваемые углеводы[163][164][165]. Кроме того, кишечная микрофлора подавляет размножение патогенных организмов за счёт конкурентного исключения. Полезные микроорганизмы кишечной микрофлоры часто продают в виде пробиотических пищевых добавок[166].
Бактерии вступают в сложные мутуалистические отношения с самыми разными животными. Например, в мезохиле[англ.] губок обитает множество бактерий, причём все исследованные к настоящему времени виды губок имеют симбиотические ассоциации с одним или более видами бактериальных симбионтов[167][168][169][170]. Многие моллюски имеют особые светящиеся органы, которые светятся благодаря обитающим в них бактериям. Бактерии получают надёжную защиту и благоприятные условия для питания, а моллюскам свечение помогает в привлечении полового партнёра[171]. Асцидии вступают в симбиотические отношения с цианобактериями рода Prochloron[англ.], который фиксирует CO2, а животное обеспечивает ему защищённое местообитание[172].
У жвачных животных в сложно устроенном желудочно-кишечном тракте обитает множество микроорганизмов, благодаря которым животные могут питаться почти что безбелковой пищей. Разрушать целлюлозу способны лишь некоторые бактерии, в результате деятельности которых образуются органические кислоты (муравьиная, уксусная, пропионовая, масляная), которые и усваиваются животными. Выделяющиеся углекислый газ и водород обитающие тут же метаногены превращают в метан. В одной из секций сложного желудка жвачных, рубце, обитают не только бактерии, разрушающие целлюлозу, но также бактерии, расщепляющие крахмал, пектин, полисахариды и пептиды, сбраживающие разнообразные сахара, спирты, аминокислоты и жирные кислоты[173]. Целлюлозоразрушающие бактерии также населяют заднюю кишку термитов, образуя ацетат, который и усваивается насекомым[174].
В почве бактерии, входящие в состав ризосферы, осуществляют фиксацию азота, превращая его в различные азотсодержащие соединения[175]. Они являются единственной усваиваемой формой азота для многих растений, которые сами не могут фиксировать азот. Множество бактерий обнаруживается на поверхности и внутри семян[176].
Патогены
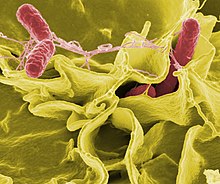
Бактерии, паразитирующие на других организмах, называют патогенами. Патогенные бактерии являются причиной множества человеческих смертей и вызывают такие инфекции, как столбняк, брюшной тиф, дифтерия, сифилис, холера, пищевые отравления, проказа и туберкулёз. Патоген, вызывающий заболевание, может быть описан много лет спустя после описания самой болезни, как, например, произошло с Helicobacter pylori и язвенной болезнью желудка. Бактерии ответственны за многие болезни культурных растений (бактериозы), в числе которых пятнистость листьев[177], ожог плодовых культур и увядание. Бактериальную основу имеют такие заболевания домашнего скота, как паратуберкулёз, мастит, сальмонеллёз и сибирская язва[178][179].
Каждый патоген характеризуется особыми взаимодействиями с организмом хозяина. Некоторые возбудители, такие как виды родов Staphylococcus и Streptococcus, вызывают кожные инфекции, пневмонию, менингит и даже сепсис, системный воспалительный ответ, переходящий в шок, массивную вазодилатацию сосудов и заканчивающийся смертью[180]. При этом те же самые микроорганизмы входят в состав нормальной микрофлоры человека и зачастую обитают на коже и внутри полости носа, не вызывая никакого заболевания. Другие бактерии всегда вызывают болезнь, например, риккетсии, которые являются облигатными внутриклеточными паразитами и могут размножаться только внутри клеток организма-хозяина. Один вид риккетсий вызывает сыпной тиф, другой является возбудителем пятнистой лихорадки Скалистых гор. Другой род облигатных внутриклеточных паразитов, Chlamydia, включает возбудителей пневмонии, инфекций мочевыводящих путей и коронарной недостаточности[181]. Некоторые бактерии, такие как Pseudomonas aeruginosa, Burkholderia cenocepacia[англ.] и Mycobacterium avium, являются оппортунистическими патогенами и вызывают заболевания преимущественно у людей, страдающих от иммунодефицита или муковисцидоза[182][183].
Бактериальные инфекции можно лечить антибиотиками, в числе которых выделяют бактерицидные препараты, которые убивают бактерии, и бактериостатики[англ.], только подавляющие их рост. Существует несколько классов антибиотиков, которые действуют на процессы, которые есть у бактерии-патогена, но нет у организма-хозяина. Так, антибиотики хлорамфеникол и пуромицин подавляют работу бактериальной рибосомы, но не действуют на эукариотические рибосомы[184]. Антибиотики используются не только в медицине, но и в животноводстве для стимуляции роста животных, что стало причиной повсеместного распространения устойчивости к антибиотикам в популяциях бактерий[185].
Хозяйственное значение
Некоторые бактерии, например, молочнокислые бактерии (Lactobacillus и Lactococcus[англ.]), наряду с дрожжами и плесневыми грибками в течение нескольких тысяч лет использовались людьми для приготовления продуктов брожения, в числе которых сыры, квашеная капуста, соевый соус, уксус, вино и йогурт[186][187].
Способность бактерий разлагать разнообразные органические соединения находит применение в переработке отходов и биоремедиации. Бактерии, способные разрушать углеводороды нефти, часто используются для устранения разливов нефти[188]. После выброса нефти из танкера «Эксон Валдез» в 1989 году в проливе Принца Вильгельма на некоторых из близлежащих пляжей были разложены удобрения, чтобы способствовать росту бактерий, разрушающих углеводороды нефти. Эта мера оказалась эффективной для тех пляжей, на которых слой нефти был не слишком толстым. Бактерии также используют для биоремедиации промышленных токсичных отходов[англ.][189]. В химической промышленности бактерии играют наиболее важную роль в производстве чистых энантиомеров химических соединений, которые используются в фармацевтике или агрохимии[190].
Бактерии можно использовать вместо пестицидов для биологической защиты растений. Наиболее часто для этих целей используется грамположительная почвенная бактерия Bacillus thuringiensis. Подвиды этой бактерии входят в состав инсектицидов, эффективных против чешуекрылых насекомых и известных под торговыми названиями Dipel и Thuricide[191]. Благодаря узкой специфичности такие пестициды считаются экологичными и безопасными для человека, диких животных, опылителей и других полезных насекомых[англ.][192][193].
Благодаря способности к быстрому росту и лёгкости манипуляций бактерии стали настоящими «рабочими лошадками» молекулярной биологии, генетики и биохимии. Внося мутации в геномы бактерий и изучая получившиеся фенотипы, учёные могут определять функции генов, ферментов и метаболических путей у бактерий и далее экстраполировать полученные данные на более сложные организмы[194]. Для хорошо изученных бактерий, таких как E. coli, даже получены математические модели их метаболизма[195][196]. Благодаря знаниям о метаболизме и генетике бактерий они могут использоваться в биотехнологии и биоинженерии для получения терапевтически важных белков, таких как инсулин, факторы роста и антитела[197][198].
История изучения

Первым бактерии увидел голландский микроскопист Антони ван Левенгук в 1676 году с помощью микроскопа с одной линзой собственного изобретения[199]. Он описал свои наблюдения в серии писем Лондонскому королевскому обществу[200][201][202]. Размеры бактерий были почти на пределе разрешающей способности простых линз Левенгука. В течение почти сотни лет после открытия Левенгука бактерий никто не видел[203]. Левенгук также описал протистов, которых назвал анималкули[англ.], и интерес к его открытиям появился в свете развития клеточной теории[204].
Термин «бактерии» предложил Христиан Готфрид Эренберг в 1828 году[205]. Описанный им род Bacterium[англ.] на самом деле оказался сборной группой неспорообразующих палочковидных бактерий[206] в противовес спорообразующим палочкам рода Bacillus, который Эренберг описал в 1835 году[207].
В 1859 году Луи Пастер показал, что в основе брожения лежит рост микроорганизмов, причём рост бактерий не был результатом их спонтанного самозарождения. Наряду со своим современником Робертом Кохом, Пастер стал первым приверженцем микробной теории болезней[208].
Роберт Кох, пионер медицинской микробиологии, работал с возбудителями холеры, сибирской язвы и туберкулёза. Коху удалось показать микробную основу туберкулёза, за что в 1905 году ему была присуждена Нобелевская премия по физиологии и медицине[209]. Так называемые постулаты Коха, которые он ввёл как критерии для установления связи между микробом и болезнью, используются и по сей день[210].
Основателем бактериологии считается Фердинанд Кон, занимавшийся изучением бактерий с 1870 года. Он стал автором классификации бактерий на основе их морфологии[211][212].
Хотя в XIX веке стало известно, что многие болезни вызываются бактериями, эффективных антибактериальных препаратов всё ещё не существовало[213]. В 1910 году Пауль Эрлих получил первый антибиотик, превратив красители для специфического окрашивания возбудителя сифилиса Treponema pallidum в соединения, которые селективно убивали бактерию[214]. В 1908 году он был удостоен Нобелевской премии за свои работы в области иммунологии, однако он также стал пионером использования красителей для идентификации бактерий[215].
Положение бактерий в системе живого мира неоднократно менялось. С самого момента открытия их отнесли к растениям как класс Schizomycetes, а ныне выделяемые в отдельный тип цианобактерий (тогда известные как синезелёные водоросли) получили название Schizophyceae. Вместе с Schizomycetes они были выделены в отдел растений под названием Schizophyta[216]. В 1866 году Эрнст Геккель выделил бактерий в тип дробянки (Monera), который рассматривал в составе царства протисты[217]. Термин «прокариоты» для обозначения организмов, не имеющих клеточного ядра, был предложен Эдуардом Шаттоном в 1925 году наряду с термином «эукариоты». Шаттон также поместил цианобактерий в группу прокариоты из группы настоящих водорослей, которые являются эукариотами[218]. В 1938 году Херберт Коупленд выделил дробянок из царства протистов, которое переименовал в протоктисты[219]. В 1969 году Роберт Уиттекер предложил деление живого мира на пять царств: дробянки, протисты, растения, грибы и животные[220]. Колоссальный шаг вперёд в понимании биологии бактерий был совершён в 1977 году, когда Карл Вёзе разделил прокариот (которых по-прежнему нередко называли дробянками) на бактерий и архей[221]. Филогенетическая таксономия, построенная на основе последовательностей гена 16S рРНК, легла в основу предложенной им трёхдоменной системы живого мира[15], которая закрепилась в 1990 году[15].
Примечания
- ↑ List of prokaryotic names with standing in nomenclature. Hierarchical classification of prokaryotes (bacteria). Version 2.2 (англ.). Leibniz Institute DSMZ (22 июня 2019). Дата обращения: 25 мая 2022. Архивировано из оригинала 4 августа 2020 года.
- ↑ Fredrickson J. K., Zachara J. M., Balkwill D. L., Kennedy D., Li S. M., Kostandarithes H. M., Daly M. J., Romine M. F., Brockman F. J. Geomicrobiology of high-level nuclear waste-contaminated vadose sediments at the hanford site, washington state. (англ.) // Applied And Environmental Microbiology. — 2004. — July (vol. 70, no. 7). — P. 4230—4241. — doi:10.1128/AEM.70.7.4230-4241.2004. — PMID 15240306.
- ↑ 1 2 Rappé M. S., Giovannoni S. J. The uncultured microbial majority. (англ.) // Annual Review Of Microbiology. — 2003. — Vol. 57. — P. 369—394. — doi:10.1146/annurev.micro.57.030502.090759. — PMID 14527284.
- ↑ Whitman W. B., Coleman D. C., Wiebe W. J. Prokaryotes: the unseen majority. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 1998. — 9 June (vol. 95, no. 12). — P. 6578—6583. — PMID 9618454.
- ↑ C. Michael Hogan. 2010. Bacteria. Encyclopedia of Earth. Eds. Sidney Draggan and C. J. Cleveland, National Council for Science and the Environment, Washington DC Архивировано 11 мая 2011 года.
- ↑ Forbes S. L. Decomposition Chemistry in a Burial Environment // Soil Analysis in Forensic Taphonomy (англ.) / Tibbett M, Carter DO. — CRC Press, 2008. — P. 203—223. — ISBN 1-4200-6991-8.
- ↑ 1 2 Choi, Charles Q. Microbes Thrive in Deepest Spot on Earth. LiveScience (17 марта 2013). Дата обращения: 17 марта 2013. Архивировано 2 апреля 2013 года.
- ↑ Glud Ronnie N., Wenzhöfer Frank, Middelboe Mathias, Oguri Kazumasa, Turnewitsch Robert, Canfield Donald E., Kitazato Hiroshi. High rates of microbial carbon turnover in sediments in the deepest oceanic trench on Earth (англ.) // Nature Geoscience. — 2013. — 17 March (vol. 6, no. 4). — P. 284—288. — ISSN 1752-0894. — doi:10.1038/ngeo1773.
- ↑ Oskin, Becky Intraterrestrials: Life Thrives in Ocean Floor. LiveScience (14 марта 2013). Дата обращения: 17 марта 2013. Архивировано 2 апреля 2013 года.
- ↑ Sender R., Fuchs S., Milo R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. (англ.) // PLoS Biology. — 2016. — August (vol. 14, no. 8). — P. e1002533—1002533. — doi:10.1371/journal.pbio.1002533. — PMID 27541692.
- ↑ Sears C. L. A dynamic partnership: celebrating our gut flora. (англ.) // Anaerobe. — 2005. — October (vol. 11, no. 5). — P. 247—251. — doi:10.1016/j.anaerobe.2005.05.001. — PMID 16701579.
- ↑ 2002 WHO mortality data. Дата обращения: 20 января 2007. Архивировано 23 октября 2013 года.
- ↑ "Metal-Mining Bacteria Are Green Chemists". Science Daily. 2 Сентябрь 2010. Архивировано 31 августа 2017.
- ↑ Ishige T., Honda K., Shimizu S. Whole organism biocatalysis. (англ.) // Current Opinion In Chemical Biology. — 2005. — April (vol. 9, no. 2). — P. 174—180. — doi:10.1016/j.cbpa.2005.02.001. — PMID 15811802.
- ↑ 1 2 3 4 Woese C. R., Kandler O., Wheelis M. L. Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 1990. — Vol. 87, no. 12. — P. 4576—4579. — PMID 2112744.
- ↑ bacterium Архивировано 27 января 2011 года., on Oxford Dictionaries.
- ↑ bacteria (англ.) в Online Etymology Dictionary[англ.]
- ↑ Schopf J. W. Disparate rates, differing fates: tempo and mode of evolution changed from the Precambrian to the Phanerozoic. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 1994. — 19 July (vol. 91, no. 15). — P. 6735—6742. — PMID 8041691.
- ↑ DeLong E. F., Pace N. R. Environmental diversity of bacteria and archaea. (англ.) // Systematic Biology. — 2001. — August (vol. 50, no. 4). — P. 470—478. — PMID 12116647.
- ↑ Brown J. R., Doolittle W. F. Archaea and the prokaryote-to-eukaryote transition. (англ.) // Microbiology And Molecular Biology Reviews : MMBR. — 1997. — December (vol. 61, no. 4). — P. 456—502. — PMID 9409149.
- ↑ Di Giulio M. The universal ancestor and the ancestor of bacteria were hyperthermophiles. (англ.) // Journal Of Molecular Evolution. — 2003. — December (vol. 57, no. 6). — P. 721—730. — doi:10.1007/s00239-003-2522-6. — PMID 14745541.
- ↑ Battistuzzi F. U., Feijao A., Hedges S. B. A genomic timescale of prokaryote evolution: insights into the origin of methanogenesis, phototrophy, and the colonization of land. (англ.) // BMC Evolutionary Biology. — 2004. — 9 November (vol. 4). — P. 44—44. — doi:10.1186/1471-2148-4-44. — PMID 15535883.
- ↑ Poole A. M., Penny D. Evaluating hypotheses for the origin of eukaryotes. (англ.) // BioEssays : News And Reviews In Molecular, Cellular And Developmental Biology. — 2007. — January (vol. 29, no. 1). — P. 74—84. — doi:10.1002/bies.20516. — PMID 17187354.
- ↑ Dyall S. D., Brown M. T., Johnson P. J. Ancient invasions: from endosymbionts to organelles. (англ.) // Science (New York, N.Y.). — 2004. — 9 April (vol. 304, no. 5668). — P. 253—257. — doi:10.1126/science.1094884. — PMID 15073369.
- ↑ Lang B. F., Gray M. W., Burger G. Mitochondrial genome evolution and the origin of eukaryotes. (англ.) // Annual Review Of Genetics. — 1999. — Vol. 33. — P. 351—397. — doi:10.1146/annurev.genet.33.1.351. — PMID 10690412.
- ↑ McFadden G. I. Endosymbiosis and evolution of the plant cell. (англ.) // Current Opinion In Plant Biology. — 1999. — December (vol. 2, no. 6). — P. 513—519. — PMID 10607659.
- ↑ Schulz H. N., Jorgensen B. B. Big bacteria. (англ.) // Annual review of microbiology. — 2001. — Vol. 55. — P. 105—137. — doi:10.1146/annurev.micro.55.1.105. — PMID 11544351.
- ↑ Williams Caroline. Who are you calling simple? (англ.) // New Scientist. — 2011. — July (vol. 211, no. 2821). — P. 38—41. — ISSN 0262-4079. — doi:10.1016/S0262-4079(11)61709-0.
- ↑ Largest bacterium ever discovered has unexpectedly complex cells. Science (23 февраля 2022). Дата обращения: 24 февраля 2022. Архивировано 23 марта 2022 года.
- ↑ Robertson J., Gomersall M., Gill P. Mycoplasma hominis: growth, reproduction, and isolation of small viable cells. (англ.) // Journal Of Bacteriology. — 1975. — November (vol. 124, no. 2). — P. 1007—1018. — PMID 1102522.
- ↑ Velimirov Branko. Nanobacteria, Ultramicrobacteria and Starvation Forms: A Search for the Smallest Metabolizing Bacterium (англ.) // Microbes and Environments. — 2001. — Vol. 16, no. 2. — P. 67—77. — ISSN 1342-6311. — doi:10.1264/jsme2.2001.67.
- ↑ Dusenbery D. B. Living at Micro Scale. — Cambridge, Massachusetts: Harvard University Press, 2009. — С. 20—25. — ISBN 978-0-674-03116-6.
- ↑ Yang D. C., Blair K. M., Salama N. R. Staying in Shape: the Impact of Cell Shape on Bacterial Survival in Diverse Environments. (англ.) // Microbiology And Molecular Biology Reviews : MMBR. — 2016. — March (vol. 80, no. 1). — P. 187—203. — doi:10.1128/MMBR.00031-15. — PMID 26864431.
- ↑ Young K. D. The selective value of bacterial shape. (англ.) // Microbiology And Molecular Biology Reviews : MMBR. — 2006. — September (vol. 70, no. 3). — P. 660—703. — doi:10.1128/MMBR.00001-06. — PMID 16959965.
- ↑ Claessen D., Rozen D. E., Kuipers O. P., Søgaard-Andersen L., van Wezel G. P. Bacterial solutions to multicellularity: a tale of biofilms, filaments and fruiting bodies. (англ.) // Nature Reviews. Microbiology. — 2014. — February (vol. 12, no. 2). — P. 115—124. — doi:10.1038/nrmicro3178. — PMID 24384602.
- ↑ Shimkets L. J. Intercellular signaling during fruiting-body development of Myxococcus xanthus. (англ.) // Annual Review Of Microbiology. — 1999. — Vol. 53. — P. 525—549. — doi:10.1146/annurev.micro.53.1.525. — PMID 10547700.
- ↑ Kaiser D. Signaling in myxobacteria. (англ.) // Annual Review Of Microbiology. — 2004. — Vol. 58. — P. 75—98. — doi:10.1146/annurev.micro.58.030603.123620. — PMID 15487930.
- ↑ Donlan R. M. Biofilms: microbial life on surfaces. (англ.) // Emerging Infectious Diseases. — 2002. — September (vol. 8, no. 9). — P. 881—890. — doi:10.3201/eid0809.020063. — PMID 12194761.
- ↑ Branda S. S., Vik S., Friedman L., Kolter R. Biofilms: the matrix revisited. (англ.) // Trends In Microbiology. — 2005. — January (vol. 13, no. 1). — P. 20—26. — doi:10.1016/j.tim.2004.11.006. — PMID 15639628.
- ↑ 1 2 Davey M. E., O'toole G. A. Microbial biofilms: from ecology to molecular genetics. (англ.) // Microbiology And Molecular Biology Reviews : MMBR. — 2000. — December (vol. 64, no. 4). — P. 847—867. — PMID 11104821.
- ↑ Donlan R. M., Costerton J. W. Biofilms: survival mechanisms of clinically relevant microorganisms. (англ.) // Clinical Microbiology Reviews. — 2002. — April (vol. 15, no. 2). — P. 167—193. — PMID 11932229.
- ↑ Slonczewski J. L., Foster J. W. Microbiology: An Evolving Science (англ.). — 3rd ed. — New York, N. Y.: W. W. Norton & Company. — P. 82. — ISBN 9780393123678.
- ↑ Lodish H., Berk A., Kaiser C. A., Krieger M., Bretscher A., Ploegh H., Amon A., Scott M. P. Molecular Cell Biology (англ.). — 7th ed. — W. H. Freeman[англ.], 2013. — P. 13. — ISBN 9781429234139.
- ↑ Bobik T. A. Polyhedral organelles compartmenting bacterial metabolic processes. (англ.) // Applied Microbiology And Biotechnology. — 2006. — May (vol. 70, no. 5). — P. 517—525. — doi:10.1007/s00253-005-0295-0. — PMID 16525780.
- ↑ Yeates T. O., Kerfeld C. A., Heinhorst S., Cannon G. C., Shively J. M. Protein-based organelles in bacteria: carboxysomes and related microcompartments. (англ.) // Nature reviews. Microbiology. — 2008. — Vol. 6, no. 9. — P. 681—691. — doi:10.1038/nrmicro1913. — PMID 18679172.
- ↑ Kerfeld C. A., Sawaya M. R., Tanaka S., Nguyen C. V., Phillips M., Beeby M., Yeates T. O. Protein structures forming the shell of primitive bacterial organelles. (англ.) // Science (New York, N.Y.). — 2005. — Vol. 309, no. 5736. — P. 936—938. — doi:10.1126/science.1113397. — PMID 16081736.
- ↑ Gitai Z. The new bacterial cell biology: moving parts and subcellular architecture. (англ.) // Cell. — 2005. — Vol. 120, no. 5. — P. 577—586. — doi:10.1016/j.cell.2005.02.026. — PMID 15766522.
- ↑ Shih Y. L., Rothfield L. The bacterial cytoskeleton. (англ.) // Microbiology and molecular biology reviews : MMBR. — 2006. — Vol. 70, no. 3. — P. 729—754. — doi:10.1128/MMBR.00017-06. — PMID 16959967.
- ↑ Norris V., den Blaauwen T., Cabin-Flaman A., Doi R. H., Harshey R., Janniere L., Jimenez-Sanchez A., Jin D. J., Levin P. A., Mileykovskaya E., Minsky A., Saier M. Jr., Skarstad K. Functional taxonomy of bacterial hyperstructures. (англ.) // Microbiology and molecular biology reviews : MMBR. — 2007. — Vol. 71, no. 1. — P. 230—253. — doi:10.1128/MMBR.00035-06. — PMID 17347523.
- ↑ Harold F. M. Conservation and transformation of energy by bacterial membranes. (англ.) // Bacteriological Reviews. — 1972. — June (vol. 36, no. 2). — P. 172—230. — PMID 4261111.
- ↑ Bryant D. A., Frigaard N. U. Prokaryotic photosynthesis and phototrophy illuminated. (англ.) // Trends in microbiology. — 2006. — Vol. 14, no. 11. — P. 488—496. — doi:10.1016/j.tim.2006.09.001. — PMID 16997562.
- ↑ Psencík J., Ikonen T. P., Laurinmäki P., Merckel M. C., Butcher S. J., Serimaa R. E., Tuma R. Lamellar organization of pigments in chlorosomes, the light harvesting complexes of green photosynthetic bacteria. (англ.) // Biophysical journal. — 2004. — Vol. 87, no. 2. — P. 1165—1172. — doi:10.1529/biophysj.104.040956. — PMID 15298919.
- ↑ Thanbichler M., Wang S. C., Shapiro L. The bacterial nucleoid: a highly organized and dynamic structure. (англ.) // Journal of cellular biochemistry. — 2005. — Vol. 96, no. 3. — P. 506—521. — doi:10.1002/jcb.20519. — PMID 15988757.
- ↑ Poehlsgaard J., Douthwaite S. The bacterial ribosome as a target for antibiotics. (англ.) // Nature Reviews. Microbiology. — 2005. — November (vol. 3, no. 11). — P. 870—881. — doi:10.1038/nrmicro1265. — PMID 16261170.
- ↑ Yeo M., Chater K. The interplay of glycogen metabolism and differentiation provides an insight into the developmental biology of Streptomyces coelicolor. (англ.) // Microbiology (Reading, England). — 2005. — March (vol. 151, no. Pt 3). — P. 855—861. — doi:10.1099/mic.0.27428-0. — PMID 15758231.
- ↑ Shiba T., Tsutsumi K., Ishige K., Noguchi T. Inorganic polyphosphate and polyphosphate kinase: their novel biological functions and applications. (англ.) // Biochemistry. Biokhimiia. — 2000. — March (vol. 65, no. 3). — P. 315—323. — PMID 10739474.
- ↑ Brune D. C. Isolation and characterization of sulfur globule proteins from Chromatium vinosum and Thiocapsa roseopersicina. (англ.) // Archives Of Microbiology. — 1995. — June (vol. 163, no. 6). — P. 391—399. — PMID 7575095.
- ↑ Kadouri D., Jurkevitch E., Okon Y., Castro-Sowinski S. Ecological and agricultural significance of bacterial polyhydroxyalkanoates. (англ.) // Critical Reviews In Microbiology. — 2005. — Vol. 31, no. 2. — P. 55—67. — doi:10.1080/10408410590899228. — PMID 15986831.
- ↑ Walsby A. E. Gas vesicles. (англ.) // Microbiological Reviews. — 1994. — March (vol. 58, no. 1). — P. 94—144. — PMID 8177173.
- ↑ van Heijenoort J. Formation of the glycan chains in the synthesis of bacterial peptidoglycan. (англ.) // Glycobiology. — 2001. — March (vol. 11, no. 3). — P. 25—36. — PMID 11320055.
- ↑ 1 2 Koch A. L. Bacterial wall as target for attack: past, present, and future research. (англ.) // Clinical Microbiology Reviews. — 2003. — October (vol. 16, no. 4). — P. 673—687. — PMID 14557293.
- ↑ 1 2 Gram H. C. Über die isolierte Färbung der Schizomyceten in Schnitt- und Trockenpräparaten (нем.) // Fortschritte der Medizin. — Berlin, 1884. — Bd. 2. — S. 185—189.
- ↑ Hugenholtz P. Exploring prokaryotic diversity in the genomic era. (англ.) // Genome biology. — 2002. — Vol. 3, no. 2. — P. 0003. — PMID 11864374.
- ↑ Walsh F. M., Amyes S. G. Microbiology and drug resistance mechanisms of fully resistant pathogens. (англ.) // Current Opinion In Microbiology. — 2004. — October (vol. 7, no. 5). — P. 439—444. — doi:10.1016/j.mib.2004.08.007. — PMID 15451497.
- ↑ Alderwick L. J., Harrison J., Lloyd G. S., Birch H. L. The Mycobacterial Cell Wall--Peptidoglycan and Arabinogalactan. (англ.) // Cold Spring Harbor Perspectives In Medicine. — 2015. — 27 March (vol. 5, no. 8). — P. 021113—021113. — doi:10.1101/cshperspect.a021113. — PMID 25818664.
- ↑ Engelhardt H., Peters J. Structural research on surface layers: a focus on stability, surface layer homology domains, and surface layer-cell wall interactions. (англ.) // Journal Of Structural Biology. — 1998. — 15 December (vol. 124, no. 2-3). — P. 276—302. — doi:10.1006/jsbi.1998.4070. — PMID 10049812.
- ↑ Beveridge T. J., Pouwels P. H., Sára M., Kotiranta A., Lounatmaa K., Kari K., Kerosuo E., Haapasalo M., Egelseer E. M., Schocher I., Sleytr U. B., Morelli L., Callegari M. L., Nomellini J. F., Bingle W. H., Smit J., Leibovitz E., Lemaire M., Miras I., Salamitou S., Béguin P., Ohayon H., Gounon P., Matuschek M., Koval S. F. Functions of S-layers. (англ.) // FEMS Microbiology Reviews. — 1997. — June (vol. 20, no. 1-2). — P. 99—149. — PMID 9276929.
- ↑ Kojima S., Blair D. F. The bacterial flagellar motor: structure and function of a complex molecular machine. (англ.) // International Review Of Cytology. — 2004. — Vol. 233. — P. 93—134. — doi:10.1016/S0074-7696(04)33003-2. — PMID 15037363.
- ↑ Beachey E. H. Bacterial adherence: adhesin-receptor interactions mediating the attachment of bacteria to mucosal surface. (англ.) // The Journal Of Infectious Diseases. — 1981. — March (vol. 143, no. 3). — P. 325—345. — PMID 7014727.
- ↑ Silverman P. M. Towards a structural biology of bacterial conjugation. (англ.) // Molecular Microbiology. — 1997. — February (vol. 23, no. 3). — P. 423—429. — PMID 9044277.
- ↑ Costa T. R., Felisberto-Rodrigues C., Meir A., Prevost M. S., Redzej A., Trokter M., Waksman G. Secretion systems in Gram-negative bacteria: structural and mechanistic insights. (англ.) // Nature Reviews. Microbiology. — 2015. — June (vol. 13, no. 6). — P. 343—359. — doi:10.1038/nrmicro3456. — PMID 25978706.
- ↑ Stokes R. W., Norris-Jones R., Brooks D. E., Beveridge T. J., Doxsee D., Thorson L. M. The glycan-rich outer layer of the cell wall of Mycobacterium tuberculosis acts as an antiphagocytic capsule limiting the association of the bacterium with macrophages. (англ.) // Infection And Immunity. — 2004. — October (vol. 72, no. 10). — P. 5676—5686. — doi:10.1128/IAI.72.10.5676-5686.2004. — PMID 15385466.
- ↑ Daffé M., Etienne G. The capsule of Mycobacterium tuberculosis and its implications for pathogenicity. (англ.) // Tubercle And Lung Disease : The Official Journal Of The International Union Against Tuberculosis And Lung Disease. — 1999. — Vol. 79, no. 3. — P. 153—169. — doi:10.1054/tuld.1998.0200. — PMID 10656114.
- ↑ Finlay B. B., Falkow S. Common themes in microbial pathogenicity revisited. (англ.) // Microbiology And Molecular Biology Reviews : MMBR. — 1997. — June (vol. 61, no. 2). — P. 136—169. — PMID 9184008.
- ↑ Nicholson W. L., Munakata N., Horneck G., Melosh H. J., Setlow P. Resistance of Bacillus endospores to extreme terrestrial and extraterrestrial environments. (англ.) // Microbiology And Molecular Biology Reviews : MMBR. — 2000. — September (vol. 64, no. 3). — P. 548—572. — PMID 10974126.
- ↑ McKenney P. T., Driks A., Eichenberger P. The Bacillus subtilis endospore: assembly and functions of the multilayered coat. (англ.) // Nature Reviews. Microbiology. — 2013. — January (vol. 11, no. 1). — P. 33—44. — doi:10.1038/nrmicro2921. — PMID 23202530.
- ↑ Nicholson W. L., Fajardo-Cavazos P., Rebeil R., Slieman T. A., Riesenman P. J., Law J. F., Xue Y. Bacterial endospores and their significance in stress resistance. (англ.) // Antonie Van Leeuwenhoek. — 2002. — August (vol. 81, no. 1-4). — P. 27—32. — PMID 12448702.
- ↑ Vreeland R. H., Rosenzweig W. D., Powers D. W. Isolation of a 250 million-year-old halotolerant bacterium from a primary salt crystal. (англ.) // Nature. — 2000. — 19 October (vol. 407, no. 6806). — P. 897—900. — doi:10.1038/35038060. — PMID 11057666.
- ↑ Cano R. J., Borucki M. K. Revival and identification of bacterial spores in 25- to 40-million-year-old Dominican amber. (англ.) // Science (New York, N.Y.). — 1995. — 19 May (vol. 268, no. 5213). — P. 1060—1064. — PMID 7538699.
- ↑ Nicholson W. L., Schuerger A. C., Setlow P. The solar UV environment and bacterial spore UV resistance: considerations for Earth-to-Mars transport by natural processes and human spaceflight. (англ.) // Mutation Research. — 2005. — 1 April (vol. 571, no. 1-2). — P. 249—264. — doi:10.1016/j.mrfmmm.2004.10.012. — PMID 15748651.
- ↑ Hatheway C. L. Toxigenic clostridia. (англ.) // Clinical Microbiology Reviews. — 1990. — January (vol. 3, no. 1). — P. 66—98. — PMID 2404569.
- ↑ Nealson K. H. Post-Viking microbiology: new approaches, new data, new insights. (англ.) // Origins of life and evolution of the biosphere : the journal of the International Society for the Study of the Origin of Life. — 1999. — Vol. 29, no. 1. — P. 73—93. — PMID 11536899.
- ↑ Xu J. Microbial ecology in the age of genomics and metagenomics: concepts, tools, and recent advances. (англ.) // Molecular Ecology. — 2006. — June (vol. 15, no. 7). — P. 1713—1731. — doi:10.1111/j.1365-294X.2006.02882.x. — PMID 16689892.
- ↑ Zillig W. Comparative biochemistry of Archaea and Bacteria. (англ.) // Current Opinion In Genetics & Development. — 1991. — December (vol. 1, no. 4). — P. 544—551. — PMID 1822288.
- ↑ 1 2 Slonczewski J. L., Foster J. W. Microbiology: An Evolving Science (англ.). — 3rd ed. — New York, N. Y.: W. W. Norton & Company. — P. 491—494. — ISBN 9780393123678.
- ↑ Hellingwerf K. J., Crielaard W., Hoff W. D., Matthijs H. C., Mur L. R., van Rotterdam B. J. Photobiology of bacteria. (англ.) // Antonie Van Leeuwenhoek. — 1994. — Vol. 65, no. 4. — P. 331—347. — PMID 7832590.
- ↑ Dalton H. The Leeuwenhoek Lecture 2000 the natural and unnatural history of methane-oxidizing bacteria. (англ.) // Philosophical Transactions Of The Royal Society Of London. Series B, Biological Sciences. — 2005. — 29 June (vol. 360, no. 1458). — P. 1207—1222. — doi:10.1098/rstb.2005.1657. — PMID 16147517.
- ↑ Zehr J. P., Jenkins B. D., Short S. M., Steward G. F. Nitrogenase gene diversity and microbial community structure: a cross-system comparison. (англ.) // Environmental Microbiology. — 2003. — July (vol. 5, no. 7). — P. 539—554. — PMID 12823187.
- ↑ Zumft W. G. Cell biology and molecular basis of denitrification. (англ.) // Microbiology And Molecular Biology Reviews : MMBR. — 1997. — December (vol. 61, no. 4). — P. 533—616. — PMID 9409151.
- ↑ Drake H. L., Daniel S. L., Küsel K., Matthies C., Kuhner C., Braus-Stromeyer S. Acetogenic bacteria: what are the in situ consequences of their diverse metabolic versatilities? (англ.) // BioFactors (Oxford, England). — 1997. — Vol. 6, no. 1. — P. 13—24. — PMID 9233536.
- ↑ Morel François M. M., Kraepiel Anne M. L., Amyot Marc. THE CHEMICAL CYCLE AND BIOACCUMULATION OF MERCURY (англ.) // Annual Review of Ecology and Systematics. — 1998. — November (vol. 29, no. 1). — P. 543—566. — ISSN 0066-4162. — doi:10.1146/annurev.ecolsys.29.1.543.
- ↑ Нетрусов, Котова, 2012, с. 108.
- ↑ Koch A. L. Control of the bacterial cell cycle by cytoplasmic growth. (англ.) // Critical Reviews In Microbiology. — 2002. — Vol. 28, no. 1. — P. 61—77. — doi:10.1080/1040-840291046696. — PMID 12003041.
- ↑ EAGON RG. Pseudomonas natriegens, a marine bacterium with a generation time of less than 10 minutes. (англ.) // Journal Of Bacteriology. — 1962. — April (vol. 83). — P. 736—737. — PMID 13888946.
- ↑ Hirsch P. Budding Bacteria (англ.) // Annual Review of Microbiology. — 1974. — October (vol. 28, no. 1). — P. 391—440. — ISSN 0066-4227. — doi:10.1146/annurev.mi.28.100174.002135.
- ↑ Stewart E. J., Madden R., Paul G., Taddei F. Aging and death in an organism that reproduces by morphologically symmetric division. (англ.) // PLoS Biology. — 2005. — February (vol. 3, no. 2). — P. e45—45. — doi:10.1371/journal.pbio.0030045. — PMID 15685293.
- ↑ 1 2 3 Thomson Jr. R. B., Bertram H. Laboratory diagnosis of central nervous system infections. (англ.) // Infectious Disease Clinics Of North America. — 2001. — December (vol. 15, no. 4). — P. 1047—1071. — PMID 11780267.
- ↑ Paerl H. W., Fulton RS 3rd., Moisander P. H., Dyble J. Harmful freshwater algal blooms, with an emphasis on cyanobacteria. (англ.) // TheScientificWorldJournal. — 2001. — 4 April (vol. 1). — P. 76—113. — doi:10.1100/tsw.2001.16. — PMID 12805693.
- ↑ Challis G. L., Hopwood D. A. Synergy and contingency as driving forces for the evolution of multiple secondary metabolite production by Streptomyces species. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2003. — 25 November (vol. 100 Suppl 2). — P. 14555—14561. — doi:10.1073/pnas.1934677100. — PMID 12970466.
- ↑ Kooijman S. A., Auger P., Poggiale J. C., Kooi B. W. Quantitative steps in symbiogenesis and the evolution of homeostasis. (англ.) // Biological Reviews Of The Cambridge Philosophical Society. — 2003. — August (vol. 78, no. 3). — P. 435—463. — PMID 14558592.
- ↑ Prats C., López D., Giró A., Ferrer J., Valls J. Individual-based modelling of bacterial cultures to study the microscopic causes of the lag phase. (англ.) // Journal Of Theoretical Biology. — 2006. — 21 August (vol. 241, no. 4). — P. 939—953. — doi:10.1016/j.jtbi.2006.01.029. — PMID 16524598.
- ↑ Hecker M., Völker U. General stress response of Bacillus subtilis and other bacteria. (англ.) // Advances In Microbial Physiology. — 2001. — Vol. 44. — P. 35—91. — PMID 11407115.
- ↑ Slonczewski J. L., Foster J. W. Microbiology: An Evolving Science (англ.). — 3rd ed. — New York, N. Y.: W. W. Norton & Company. — P. 143. — ISBN 9780393123678.
- ↑ Pradella S., Hans A., Spröer C., Reichenbach H., Gerth K., Beyer S. Characterisation, genome size and genetic manipulation of the myxobacterium Sorangium cellulosum So ce56. (англ.) // Archives Of Microbiology. — 2002. — December (vol. 178, no. 6). — P. 484—492. — doi:10.1007/s00203-002-0479-2. — PMID 12420170.
- ↑ Schneiker S., Perlova O., Kaiser O., Gerth K., Alici A., Altmeyer M. O., Bartels D., Bekel T., Beyer S., Bode E., Bode H. B., Bolten C. J., Choudhuri J. V., Doss S., Elnakady Y. A., Frank B., Gaigalat L., Goesmann A., Groeger C., Gross F., Jelsbak L., Jelsbak L., Kalinowski J., Kegler C., Knauber T., Konietzny S., Kopp M., Krause L., Krug D., Linke B., Mahmud T., Martinez-Arias R., McHardy A. C., Merai M., Meyer F., Mormann S., Muñoz-Dorado J., Perez J., Pradella S., Rachid S., Raddatz G., Rosenau F., Rückert C., Sasse F., Scharfe M., Schuster S. C., Suen G., Treuner-Lange A., Velicer G. J., Vorhölter F. J., Weissman K. J., Welch R. D., Wenzel S. C., Whitworth D. E., Wilhelm S., Wittmann C., Blöcker H., Pühler A., Müller R. Complete genome sequence of the myxobacterium Sorangium cellulosum. (англ.) // Nature Biotechnology. — 2007. — November (vol. 25, no. 11). — P. 1281—1289. — doi:10.1038/nbt1354. — PMID 17965706.
- ↑ Hinnebusch J., Tilly K. Linear plasmids and chromosomes in bacteria. (англ.) // Molecular Microbiology. — 1993. — December (vol. 10, no. 5). — P. 917—922. — PMID 7934868.
- ↑ Lin Y. S., Kieser H. M., Hopwood D. A., Chen C. W. The chromosomal DNA of Streptomyces lividans 66 is linear. (англ.) // Molecular Microbiology. — 1993. — December (vol. 10, no. 5). — P. 923—933. — PMID 7934869.
- ↑ Val M. E., Soler-Bistué A., Bland M. J., Mazel D. Management of multipartite genomes: the Vibrio cholerae model. (англ.) // Current Opinion In Microbiology. — 2014. — December (vol. 22). — P. 120—126. — doi:10.1016/j.mib.2014.10.003. — PMID 25460805.
- ↑ Kado C. I. Historical Events That Spawned the Field of Plasmid Biology. (англ.) // Microbiology Spectrum. — 2014. — October (vol. 2, no. 5). — doi:10.1128/microbiolspec.PLAS-0019-2013. — PMID 26104369.
- ↑ Belfort M., Reaban M. E., Coetzee T., Dalgaard J. Z. Prokaryotic introns and inteins: a panoply of form and function. (англ.) // Journal Of Bacteriology. — 1995. — July (vol. 177, no. 14). — P. 3897—3903. — PMID 7608058.
- ↑ Denamur E., Matic I. Evolution of mutation rates in bacteria. (англ.) // Molecular Microbiology. — 2006. — May (vol. 60, no. 4). — P. 820—827. — doi:10.1111/j.1365-2958.2006.05150.x. — PMID 16677295.
- ↑ Wright B. E. Stress-directed adaptive mutations and evolution. (англ.) // Molecular Microbiology. — 2004. — May (vol. 52, no. 3). — P. 643—650. — doi:10.1111/j.1365-2958.2004.04012.x. — PMID 15101972.
- ↑ Chen I., Dubnau D. DNA uptake during bacterial transformation. (англ.) // Nature reviews. Microbiology. — 2004. — Vol. 2, no. 3. — P. 241—249. — doi:10.1038/nrmicro844. — PMID 15083159.
- ↑ Johnsborg O., Eldholm V., Håvarstein L. S. Natural genetic transformation: prevalence, mechanisms and function. (англ.) // Research In Microbiology. — 2007. — December (vol. 158, no. 10). — P. 767—778. — doi:10.1016/j.resmic.2007.09.004. — PMID 17997281.
- ↑ Bernstein H., Bernstein C., Michod R. E. DNA repair as the primary adaptive function of sex in bacteria and eukaryotes (англ.) / Sakura Kimura and Sora Shimizu. — N. Y.: Nova Science Publishers, Hauppauge, 2012. — Vol. Chapter 1. — P. 49. — ISBN 978-1-62100-808-8.
- ↑ Brüssow H., Canchaya C., Hardt W. D. Phages and the evolution of bacterial pathogens: from genomic rearrangements to lysogenic conversion. (англ.) // Microbiology And Molecular Biology Reviews : MMBR. — 2004. — September (vol. 68, no. 3). — P. 560—602. — doi:10.1128/MMBR.68.3.560-602.2004. — PMID 15353570.
- ↑ Bickle T. A., Krüger D. H. Biology of DNA restriction. (англ.) // Microbiological Reviews. — 1993. — June (vol. 57, no. 2). — P. 434—450. — PMID 8336674.
- ↑ Barrangou R., Fremaux C., Deveau H., Richards M., Boyaval P., Moineau S., Romero D. A., Horvath P. CRISPR provides acquired resistance against viruses in prokaryotes // Science. — 2007. — Vol. 315, no. 5819. — P. 1709—1712. — doi:10.1126/science.1138140. — PMID 17379808.
- ↑ Brouns S. J., Jore M. M., Lundgren M., Westra E. R., Slijkhuis R. J., Snijders A. P., Dickman M. J., Makarova K. S., Koonin E. V., van der Oost J. Small CRISPR RNAs guide antiviral defense in prokaryotes. (англ.) // Science (New York, N.Y.). — 2008. — 15 August (vol. 321, no. 5891). — P. 960—964. — doi:10.1126/science.1159689. — PMID 18703739.
- ↑ Michod R. E., Bernstein H., Nedelcu A. M. Adaptive value of sex in microbial pathogens. (англ.) // Infection, genetics and evolution : journal of molecular epidemiology and evolutionary genetics in infectious diseases. — 2008. — Vol. 8, no. 3. — P. 267—285. — doi:10.1016/j.meegid.2008.01.002. — PMID 18295550.
- ↑ Hastings P. J., Rosenberg S. M., Slack A. Antibiotic-induced lateral transfer of antibiotic resistance. (англ.) // Trends In Microbiology. — 2004. — September (vol. 12, no. 9). — P. 401—404. — doi:10.1016/j.tim.2004.07.003. — PMID 15337159.
- ↑ Davison J. Genetic exchange between bacteria in the environment. (англ.) // Plasmid. — 1999. — September (vol. 42, no. 2). — P. 73—91. — doi:10.1006/plas.1999.1421. — PMID 10489325.
- ↑ 1 2 3 Bardy S. L., Ng S. Y., Jarrell K. F. Prokaryotic motility structures. (англ.) // Microbiology (Reading, England). — 2003. — February (vol. 149, no. Pt 2). — P. 295—304. — doi:10.1099/mic.0.25948-0. — PMID 12624192.
- ↑ Macnab R. M. The bacterial flagellum: reversible rotary propellor and type III export apparatus. (англ.) // Journal Of Bacteriology. — 1999. — December (vol. 181, no. 23). — P. 7149—7153. — PMID 10572114.
- ↑ Wu M., Roberts J. W., Kim S., Koch D. L., DeLisa M. P. Collective bacterial dynamics revealed using a three-dimensional population-scale defocused particle tracking technique. (англ.) // Applied And Environmental Microbiology. — 2006. — July (vol. 72, no. 7). — P. 4987—4994. — doi:10.1128/AEM.00158-06. — PMID 16820497.
- ↑ Mattick J. S. Type IV pili and twitching motility. (англ.) // Annual Review Of Microbiology. — 2002. — Vol. 56. — P. 289—314. — doi:10.1146/annurev.micro.56.012302.160938. — PMID 12142488.
- ↑ Merz A. J., So M., Sheetz M. P. Pilus retraction powers bacterial twitching motility. (англ.) // Nature. — 2000. — 7 September (vol. 407, no. 6800). — P. 98—102. — doi:10.1038/35024105. — PMID 10993081.
- ↑ Lux R., Shi W. Chemotaxis-guided movements in bacteria. (англ.) // Critical Reviews In Oral Biology And Medicine : An Official Publication Of The American Association Of Oral Biologists. — 2004. — 1 July (vol. 15, no. 4). — P. 207—220. — PMID 15284186.
- ↑ Schweinitzer T., Josenhans C. Bacterial energy taxis: a global strategy? (англ.) // Archives Of Microbiology. — 2010. — July (vol. 192, no. 7). — P. 507—520. — doi:10.1007/s00203-010-0575-7. — PMID 20411245.
- ↑ Frankel R. B., Bazylinski D. A., Johnson M. S., Taylor B. L. Magneto-aerotaxis in marine coccoid bacteria. (англ.) // Biophysical Journal. — 1997. — August (vol. 73, no. 2). — P. 994—1000. — doi:10.1016/S0006-3495(97)78132-3. — PMID 9251816.
- ↑ Goldberg M. B. Actin-based motility of intracellular microbial pathogens. (англ.) // Microbiology And Molecular Biology Reviews : MMBR. — 2001. — December (vol. 65, no. 4). — P. 595—626. — doi:10.1128/MMBR.65.4.595-626.2001. — PMID 11729265.
- ↑ Dusenbery, David B. Life at Small Scale. — Scientific American Library. — 1996. — ISBN 0-7167-5060-0.
- ↑ Shapiro J. A. Thinking about bacterial populations as multicellular organisms. (англ.) // Annual Review Of Microbiology. — 1998. — Vol. 52. — P. 81—104. — doi:10.1146/annurev.micro.52.1.81. — PMID 9891794.
- ↑ 1 2 Costerton J. W., Lewandowski Z., Caldwell D. E., Korber D. R., Lappin-Scott H. M. Microbial biofilms. (англ.) // Annual Review Of Microbiology. — 1995. — Vol. 49. — P. 711—745. — doi:10.1146/annurev.mi.49.100195.003431. — PMID 8561477.
- ↑ Miller M. B., Bassler B. L. Quorum sensing in bacteria. (англ.) // Annual Review Of Microbiology. — 2001. — Vol. 55. — P. 165—199. — doi:10.1146/annurev.micro.55.1.165. — PMID 11544353.
- ↑ Boucher Y., Douady C. J., Papke R. T., Walsh D. A., Boudreau M. E., Nesbø C. L., Case R. J., Doolittle W. F. Lateral gene transfer and the origins of prokaryotic groups. (англ.) // Annual Review Of Genetics. — 2003. — Vol. 37. — P. 283—328. — doi:10.1146/annurev.genet.37.050503.084247. — PMID 14616063.
- ↑ Olsen G. J., Woese C. R., Overbeek R. The winds of (evolutionary) change: breathing new life into microbiology. (англ.) // Journal Of Bacteriology. — 1994. — January (vol. 176, no. 1). — P. 1—6. — PMID 8282683.
- ↑ IJSEM Home. Ijs.sgmjournals.org (28 октября 2011). Дата обращения: 4 ноября 2011. Архивировано 19 октября 2011 года.
- ↑ Bergey's Manual Trust. Bergeys.org. Дата обращения: 4 ноября 2011. Архивировано 7 ноября 2011 года.
- ↑ Gupta R. S. The natural evolutionary relationships among prokaryotes. (англ.) // Critical Reviews In Microbiology. — 2000. — Vol. 26, no. 2. — P. 111—131. — doi:10.1080/10408410091154219. — PMID 10890353.
- ↑ Cavalier-Smith T. The neomuran origin of archaebacteria, the negibacterial root of the universal tree and bacterial megaclassification. (англ.) // International journal of systematic and evolutionary microbiology. — 2002. — Vol. 52, no. Pt 1. — P. 7—76. — doi:10.1099/00207713-52-1-7. — PMID 11837318.
- ↑ Doolittle R. F. Evolutionary aspects of whole-genome biology. (англ.) // Current Opinion In Structural Biology. — 2005. — June (vol. 15, no. 3). — P. 248—253. — doi:10.1016/j.sbi.2005.04.001. — PMID 15963888.
- ↑ Woods G. L., Walker D. H. Detection of infection or infectious agents by use of cytologic and histologic stains. (англ.) // Clinical Microbiology Reviews. — 1996. — July (vol. 9, no. 3). — P. 382—404. — PMID 8809467.
- ↑ Sutherland John B., Rafii Fatemeh. Cultural, Serological, and Genetic Methods for Identification of Bacteria (англ.) // Identification of Microorganisms by Mass Spectrometry. — 2006. — 3 January. — P. 1—21. — ISBN 9780471654421. — doi:10.1002/0471748641.ch1.
- ↑ Weinstein M. P. Clinical importance of blood cultures. (англ.) // Clinics In Laboratory Medicine. — 1994. — March (vol. 14, no. 1). — P. 9—16. — PMID 8181237.
- ↑ Louie M., Louie L., Simor A. E. The role of DNA amplification technology in the diagnosis of infectious diseases. (англ.) // CMAJ : Canadian Medical Association Journal = Journal De L'Association Medicale Canadienne. — 2000. — 8 August (vol. 163, no. 3). — P. 301—309. — PMID 10951731.
- ↑ Oliver J. D. The viable but nonculturable state in bacteria. (англ.) // Journal Of Microbiology (Seoul, Korea). — 2005. — February (vol. 43 Spec No). — P. 93—100. — PMID 15765062.
- ↑ Talaro K. P., Chess B. Foundations in Microbiology. — New York: McGraw-Hill Education. — P. 111. — ISBN 978-1-259-70521-2.
- ↑ Curtis T. P., Sloan W. T., Scannell J. W. Estimating prokaryotic diversity and its limits. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2002. — 6 August (vol. 99, no. 16). — P. 10494—10499. — doi:10.1073/pnas.142680199. — PMID 12097644.
- ↑ Schloss P. D., Handelsman J. Status of the microbial census. (англ.) // Microbiology And Molecular Biology Reviews : MMBR. — 2004. — December (vol. 68, no. 4). — P. 686—691. — doi:10.1128/MMBR.68.4.686-691.2004. — PMID 15590780.
- ↑ Riley M. A., Lizotte-Waniewski M. Population genomics and the bacterial species concept. (англ.) // Methods In Molecular Biology (Clifton, N.J.). — 2009. — Vol. 532. — P. 367—377. — doi:10.1007/978-1-60327-853-9_21. — PMID 19271196.
- ↑ List of prokaryotic names with standing in nomenclature. Candidatus. Дата обращения: 7 декабря 2018. Архивировано 15 июня 2018 года.
- ↑ Dijkshoorn L., Ursing B. M., Ursing J. B. Strain, clone and species: comments on three basic concepts of bacteriology. (англ.) // Journal Of Medical Microbiology. — 2000. — May (vol. 49, no. 5). — P. 397—401. — doi:10.1099/0022-1317-49-5-397. — PMID 10798550.
- ↑ Fisher, Bruce; Harvey, Richard P.; Champe, Pamela C. Lippincott's Illustrated Reviews: Microbiology (Lippincott's Illustrated Reviews Series) (англ.). — Hagerstown, MD: Lippincott Williams & Wilkins[англ.], 2007. — P. 367—392. — ISBN 0-7817-8215-5.
- ↑ LEF.org > Bacterial Infections Updated: 19 January 2006. Retrieved on 11 April 2009
- ↑ Martin M. O. Predatory prokaryotes: an emerging research opportunity. (англ.) // Journal Of Molecular Microbiology And Biotechnology. — 2002. — September (vol. 4, no. 5). — P. 467—477. — PMID 12432957.
- ↑ Velicer G. J., Stredwick K. L. Experimental social evolution with Myxococcus xanthus. (англ.) // Antonie Van Leeuwenhoek. — 2002. — August (vol. 81, no. 1-4). — P. 155—164. — PMID 12448714.
- ↑ Громов Б. В., Мамкаева К. А. Электронно-микроскопическое изучение паразитизма бактерии Вdellovibrio chlorellavorus на клетках зеленой водоросли Сhlorella pulgaris // Цитология. — Л.: Наука, 1972. — Т. 14, № 2. — С. 256—261.
- ↑ Guerrero R., Pedros-Alio C., Esteve I., Mas J., Chase D., Margulis L. Predatory prokaryotes: predation and primary consumption evolved in bacteria. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 1986. — April (vol. 83, no. 7). — P. 2138—2142. — PMID 11542073.
- ↑ Velicer G. J., Mendes-Soares H. Bacterial predators. (англ.) // Current Biology : CB. — 2009. — 27 January (vol. 19, no. 2). — P. 55—56. — doi:10.1016/j.cub.2008.10.043. — PMID 19174136.
- ↑ Stams A. J., de Bok F. A., Plugge C. M., van Eekert M. H., Dolfing J., Schraa G. Exocellular electron transfer in anaerobic microbial communities. (англ.) // Environmental Microbiology. — 2006. — March (vol. 8, no. 3). — P. 371—382. — doi:10.1111/j.1462-2920.2006.00989.x. — PMID 16478444.
- ↑ Нетрусов, Котова, 2012, с. 289.
- ↑ O'Hara A. M., Shanahan F. The gut flora as a forgotten organ. (англ.) // EMBO Reports. — 2006. — July (vol. 7, no. 7). — P. 688—693. — doi:10.1038/sj.embor.7400731. — PMID 16819463.
- ↑ Zoetendal E. G., Vaughan E. E., de Vos W. M. A microbial world within us. (англ.) // Molecular Microbiology. — 2006. — March (vol. 59, no. 6). — P. 1639—1650. — doi:10.1111/j.1365-2958.2006.05056.x. — PMID 16553872.
- ↑ Gorbach S. L. Lactic acid bacteria and human health. (англ.) // Annals Of Medicine. — 1990. — February (vol. 22, no. 1). — P. 37—41. — PMID 2109988.
- ↑ Salminen S. J., Gueimonde M., Isolauri E. Probiotics that modify disease risk. (англ.) // The Journal Of Nutrition. — 2005. — May (vol. 135, no. 5). — P. 1294—1298. — doi:10.1093/jn/135.5.1294. — PMID 15867327.
- ↑ Althoff K., Schütt C., Steffen R., Batel R., Müller W. E. G. Evidence for a symbiosis between bacteria of the genus Rhodobacter and the marine sponge Halichondria panicea : harbor also for putatively toxic bacteria? (англ.) // Marine Biology. — 1998. — 9 February (vol. 130, no. 3). — P. 529—536. — ISSN 0025-3162. — doi:10.1007/s002270050273.
- ↑ Hentschel U., Usher K. M., Taylor M. W. Marine sponges as microbial fermenters // FEMS Microbiology Ecology. — 2006. — Vol. 55, no. 2. — P. 167—177. — doi:10.1111/j.1574-6941.2005.00046.x. — PMID 16420625.
- ↑ Sarà M., Bavestrello G., Cattaneo-Vietti R., Cerrano C. Endosymbiosis in sponges: Relevance for epigenesis and evolution // Symbiosis. — 1998. — Vol. 25, no. 1. — P. 57—70.
- ↑ Taylor M. W., Radax R., Steger D., Wagner M. Sponge-associated microorganisms: evolution, ecology, and biotechnological potential // Microbiology and Molecular Biology Reviews. — 2007. — Vol. 71, no. 2. — P. 295—347. — doi:10.1128/MMBR.00040-06. — PMID 17554047.
- ↑ Нетрусов, Котова, 2012, с. 284.
- ↑ Нетрусов, Котова, 2012, с. 286.
- ↑ Нетрусов, Котова, 2012, с. 286—287.
- ↑ Нетрусов, Котова, 2012, с. 288.
- ↑ Barea J. M., Pozo M. J., Azcón R., Azcón-Aguilar C. Microbial co-operation in the rhizosphere. (англ.) // Journal Of Experimental Botany. — 2005. — July (vol. 56, no. 417). — P. 1761—1778. — doi:10.1093/jxb/eri197. — PMID 15911555.
- ↑ Нетрусов, Котова, 2012, с. 291.
- ↑ Leaf Spot Diseases of Shade Trees and Ornamentals. Missouri Botanical Garden. Дата обращения: 11 ноября 2014. Архивировано 31 мая 2019 года.
- ↑ Bacterial diseases of animals. Дата обращения: 10 декабря 2018. Архивировано 11 декабря 2018 года.
- ↑ Human bacterial diseases. Дата обращения: 10 декабря 2018. Архивировано 11 декабря 2018 года.
- ↑ Fish D. N. Optimal antimicrobial therapy for sepsis. (англ.) // American Journal Of Health-system Pharmacy : AJHP : Official Journal Of The American Society Of Health-System Pharmacists. — 2002. — 15 February (vol. 59 Suppl 1). — P. 13—19. — PMID 11885408.
- ↑ Belland R. J., Ouellette S. P., Gieffers J., Byrne G. I. Chlamydia pneumoniae and atherosclerosis. (англ.) // Cellular Microbiology. — 2004. — February (vol. 6, no. 2). — P. 117—127. — PMID 14706098.
- ↑ Heise E. R. Diseases associated with immunosuppression. (англ.) // Environmental Health Perspectives. — 1982. — February (vol. 43). — P. 9—19. — doi:10.1289/ehp.82439. — PMID 7037390.
- ↑ Saiman L. Microbiology of early CF lung disease. (англ.) // Paediatric Respiratory Reviews. — 2004. — Vol. 5 Suppl A. — P. 367—369. — PMID 14980298.
- ↑ Yonath A., Bashan A. Ribosomal crystallography: initiation, peptide bond formation, and amino acid polymerization are hampered by antibiotics. (англ.) // Annual Review Of Microbiology. — 2004. — Vol. 58. — P. 233—251. — doi:10.1146/annurev.micro.58.030603.123822. — PMID 15487937.
- ↑ Khachatourians G. G. Agricultural use of antibiotics and the evolution and transfer of antibiotic-resistant bacteria. (англ.) // CMAJ : Canadian Medical Association Journal = Journal De L'Association Medicale Canadienne. — 1998. — 3 November (vol. 159, no. 9). — P. 1129—1136. — PMID 9835883.
- ↑ Johnson M. E., Lucey J. A. Major technological advances and trends in cheese. (англ.) // Journal Of Dairy Science. — 2006. — April (vol. 89, no. 4). — P. 1174—1178. — doi:10.3168/jds.S0022-0302(06)72186-5. — PMID 16537950.
- ↑ Hagedorn S., Kaphammer B. Microbial biocatalysis in the generation of flavor and fragrance chemicals. (англ.) // Annual Review Of Microbiology. — 1994. — Vol. 48. — P. 773—800. — doi:10.1146/annurev.mi.48.100194.004013. — PMID 7826026.
- ↑ Cohen Y. Bioremediation of oil by marine microbial mats. (англ.) // International Microbiology : The Official Journal Of The Spanish Society For Microbiology. — 2002. — December (vol. 5, no. 4). — P. 189—193. — doi:10.1007/s10123-002-0089-5. — PMID 12497184.
- ↑ Neves L. C., Miyamura T. T., Moraes D. A., Penna T. C., Converti A. Biofiltration methods for the removal of phenolic residues. (англ.) // Applied Biochemistry And Biotechnology. — 2006. — Vol. 129-132. — P. 130—152. — PMID 16915636.
- ↑ Liese A., Filho M. V. Production of fine chemicals using biocatalysis. (англ.) // Current Opinion In Biotechnology. — 1999. — December (vol. 10, no. 6). — P. 595—603. — PMID 10600695.
- ↑ Aronson A. I., Shai Y. Why Bacillus thuringiensis insecticidal toxins are so effective: unique features of their mode of action. (англ.) // FEMS Microbiology Letters. — 2001. — 5 February (vol. 195, no. 1). — P. 1—8. — PMID 11166987.
- ↑ Bozsik A. Susceptibility of adult Coccinella septempunctata (Coleoptera: Coccinellidae) to insecticides with different modes of action. (англ.) // Pest Management Science. — 2006. — July (vol. 62, no. 7). — P. 651—654. — doi:10.1002/ps.1221. — PMID 16649191.
- ↑ Chattopadhyay A., Bhatnagar N. B., Bhatnagar R. Bacterial insecticidal toxins. (англ.) // Critical Reviews In Microbiology. — 2004. — Vol. 30, no. 1. — P. 33—54. — doi:10.1080/10408410490270712. — PMID 15116762.
- ↑ Serres M. H., Gopal S., Nahum L. A., Liang P., Gaasterland T., Riley M. A functional update of the Escherichia coli K-12 genome. (англ.) // Genome Biology. — 2001. — Vol. 2, no. 9. — P. 0035—0035. — PMID 11574054.
- ↑ Almaas E., Kovács B., Vicsek T., Oltvai Z. N., Barabási A. L. Global organization of metabolic fluxes in the bacterium Escherichia coli. (англ.) // Nature. — 2004. — 26 February (vol. 427, no. 6977). — P. 839—843. — doi:10.1038/nature02289. — PMID 14985762.
- ↑ Reed J. L., Vo T. D., Schilling C. H., Palsson B. O. An expanded genome-scale model of Escherichia coli K-12 (iJR904 GSM/GPR). (англ.) // Genome biology. — 2003. — Vol. 4, no. 9. — P. 54. — doi:10.1186/gb-2003-4-9-r54. — PMID 12952533.
- ↑ Walsh G. Therapeutic insulins and their large-scale manufacture. (англ.) // Applied Microbiology And Biotechnology. — 2005. — April (vol. 67, no. 2). — P. 151—159. — doi:10.1007/s00253-004-1809-x. — PMID 15580495.
- ↑ Graumann K., Premstaller A. Manufacturing of recombinant therapeutic proteins in microbial systems. (англ.) // Biotechnology Journal. — 2006. — February (vol. 1, no. 2). — P. 164—186. — doi:10.1002/biot.200500051. — PMID 16892246.
- ↑ Porter J. R. Antony van Leeuwenhoek: tercentenary of his discovery of bacteria. (англ.) // Bacteriological Reviews. — 1976. — June (vol. 40, no. 2). — P. 260—269. — PMID 786250.
- ↑ Leewenhoeck A. An Abstract of a Letter from Mr. Anthony Leewenhoeck at Delft, Dated Sep. 17. 1683. Containing Some Microscopical Observations, about Animals in the Scurf of the Teeth, the Substance Call'd Worms in the Nose, the Cuticula Consisting of Scales (англ.) // Philosophical Transactions of the Royal Society of London. — 1684. — 1 January (vol. 14, no. 155-166). — P. 568—574. — ISSN 0261-0523. — doi:10.1098/rstl.1684.0030.
- ↑ van Leeuwenhoek A. Part of a Letter from Mr Antony van Leeuwenhoek, concerning the Worms in Sheeps Livers, Gnats, and Animalcula in the Excrements of Frogs (англ.) // Philosophical Transactions of the Royal Society of London. — 1700. — 1 January (vol. 22, no. 260-276). — P. 509—518. — ISSN 0261-0523. — doi:10.1098/rstl.1700.0013.
- ↑ van Leeuwenhoek A. Part of a Letter from Mr Antony van Leeuwenhoek, F. R. S. concerning Green Weeds Growing in Water, and Some Animalcula Found about Them (англ.) // Philosophical Transactions of the Royal Society of London. — 1702. — 1 January (vol. 23, no. 277-288). — P. 1304—1311. — ISSN 0261-0523. — doi:10.1098/rstl.1702.0042.
- ↑ Asimov I. Asimov's Biographical Encyclopedia of Science and Technology (англ.). — 2nd ed. — Garden City, New York: Doubleday and Company, 1982. — P. 143.
- ↑ [biology.about.com/od/biologydictionary/g/celltheory.htm The modern version of the Cell Theory]. Дата обращения: 25 марта 2018. Архивировано 12 февраля 2015 года.
- ↑ Ehrenberg, C. G. Symbolae physicae animalia evertebrata exclusis insectis. Series prima cum tabularum decade prima continent animalia Africana et Asiatica. Decas Prima // Symbolae physicae, seu Icones adhue ineditae corporum naturalium novorum aut minus cognitorum, quae ex itineribus per Libyam, Aegyptum, Nubiam, Dengalam, Syriam, Arabiam et Habessiniam. Pars Zoologica, 4 / Hemprich F. G. & Ehrenberg C. G. (eds.). — Berlin : Officina Academica, 1828—1831.
- ↑ Breed R. S., Conn H. J. The Status of the Generic Term Bacterium Ehrenberg 1828. (англ.) // Journal Of Bacteriology. — 1936. — May (vol. 31, no. 5). — P. 517—518. — PMID 16559906.
- ↑ Ehrenberg C. G. Dritter Beitrag zur Erkenntniss grosser Organisation in der Richtung des kleinsten Raumes (нем.) // Physikalische Abhandlungen der Koeniglichen Akademie der Wissenschaften zu Berlin aus den Jahren 1833—1835. — Berlin, 1835. — S. 143—336.
- ↑ Pasteur's Papers on the Germ Theory. LSU Law Center's Medical and Public Health Law Site, Historic Public Health Articles. Дата обращения: 23 ноября 2006. Архивировано 18 декабря 2006 года.
- ↑ The Nobel Prize in Physiology or Medicine 1905. Nobelprize.org. Дата обращения: 22 ноября 2006. Архивировано 10 декабря 2006 года.
- ↑ O'Brien S. J., Goedert J. J. HIV causes AIDS: Koch's postulates fulfilled. (англ.) // Current Opinion In Immunology. — 1996. — October (vol. 8, no. 5). — P. 613—618. — PMID 8902385.
- ↑ Chung, King-Thom Ferdinand Julius Cohn (1828-1898): Pioneer of Bacteriology. Department of Microbiology and Molecular Cell Sciences, The University of Memphis. Архивировано 27 июля 2011 года.
- ↑ Drews, Gerhart. Ferdinand Cohn, a founder of modern microbiology (англ.) // ASM News. — 1999. — Vol. 65, no. 8. — P. 547—552. Архивировано 13 июля 2017 года. Архивированная копия. Дата обращения: 15 октября 2018. Архивировано из оригинала 13 июля 2017 года.
- ↑ Thurston A. J. Of blood, inflammation and gunshot wounds: the history of the control of sepsis. (англ.) // The Australian And New Zealand Journal Of Surgery. — 2000. — December (vol. 70, no. 12). — P. 855—861. — PMID 11167573.
- ↑ Schwartz R. S. Paul Ehrlich's magic bullets. (англ.) // The New England Journal Of Medicine. — 2004. — 11 March (vol. 350, no. 11). — P. 1079—1080. — doi:10.1056/NEJMp048021. — PMID 15014180.
- ↑ Biography of Paul Ehrlich. Nobelprize.org. Дата обращения: 26 ноября 2006. Архивировано 28 ноября 2006 года.
- ↑ C. Von Nägeli. Bericht über die Verhandlungen der 33. Versammlung deutscher Naturforscher und Aerzte, gehalten in Bonn von 18 bis 24 September 1857 (нем.) // Botanische Zeitung : magazin / R. Caspary. — 1857. — Bd. 15. — S. 749—776.<https://archive.org/stream/botanischezeitun15mohl#page/372/mode/2up>
- ↑ Haeckel, Ernst. Generelle Morphologie der Organismen (нем.). — Reimer, Berlin, 1867. — ISBN 978-1-144-00186-3.
- ↑ É. Chatton. Pansporella perplexa. Réflexions sur la biologie et la phylogénie des protozoaires (фр.) // Ann. Sci. Nat. Zool. : magazine. — 1925. — Vol. 10—VII. — P. 1—84.
- ↑ Copeland Herbert F. The Kingdoms of Organisms (англ.) // The Quarterly Review of Biology. — 1938. — December (vol. 13, no. 4). — P. 383—420. — ISSN 0033-5770. — doi:10.1086/394568.
- ↑ Whittaker R. H. New concepts of kingdoms or organisms. Evolutionary relations are better represented by new classifications than by the traditional two kingdoms. (англ.) // Science (New York, N.Y.). — 1969. — 10 January (vol. 163, no. 3863). — P. 150—160. — PMID 5762760.
- ↑ Woese C. R., Fox G. E. Phylogenetic structure of the prokaryotic domain: the primary kingdoms. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 1977. — November (vol. 74, no. 11). — P. 5088—5090. — PMID 270744.
Литература
- Пиневич А. В. Микробиология. Биология прокариотов: в 3 т. — СПб.: Издательство С.-Петербургского университета, 2006. — ISBN 5-288-04056-7.
- Нетрусов А. И., Котова И. Б. Микробиология. — 4-е изд., перераб. и доп. — М.: Издательский центр «Академия», 2012. — 384 с. — ISBN 978-5-7695-7979-0.
- Современная микробиология: в 2 т / Под ред. Й. Ленгелера, Г. Древса, Г. Шлегеля. — М.: Мир, 2005.
| Таксономия | ||||
|---|---|---|---|---|
| Патогенные бактерии | |
|---|---|
| Микрофлора человека | |
| Субстратная специфичность | |
| Дыхание | |
| Генетика и размножение | |
| Покоящиеся формы | |
| См. также | |