
Мейо́з, или редукционное деление — деление ядра эукариотической клетки с уменьшением числа хромосом в два раза. Происходит в два этапа. В результате мейоза образуются гаметы, споры и другие зародышевые клетки.

Кроссинго́вер — процесс обмена участками гомологичных хромосом во время конъюгации в профазе первого деления мейоза, которое происходит, например, при образовании гамет или спор. Помимо мейотического, описан также митотический кроссинговер.

Центромера — участок хромосомы, который связывает сестринские хроматиды, играет важную роль в процессе деления клеточного ядра и участвует в контроле экспрессии генов. Характеризуется специфическими последовательностью нуклеотидов и структурой.

Рекомбинация — перераспределение генетического материала путём разрыва и соединения разных молекул, приводящее к появлению новых комбинаций генов или других нуклеотидных последовательностей. В широком смысле слова включает в себя не только рекомбинацию между молекулами ДНК, но и перекомбинацию (сортировку) генетического материала на уровне целых хромосом или ядер, а также обмен плазмидами между клетками.
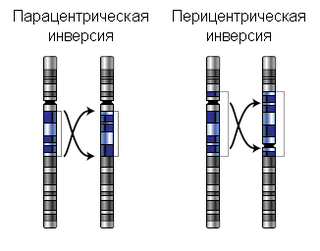
Инве́рсия — хромосомная перестройка, при которой происходит поворот участка хромосомы на 180°. Инверсии являются сбалансированными внутрихромосомными перестройками. Различают парацентрические и перицентрические инверсии. Инверсии играют роль в эволюционном процессе, видообразовании и в нарушениях фертильности.

Гетерохромати́н — участки хроматина, находящиеся в течение клеточного цикла в конденсированном (компактном) состоянии. Особенностью гетерохроматиновой ДНК является крайне низкая транскрибируемость.
Псевдоаутосо́мные о́бласти — гомологичные участки половых хромосом различного типа; у млекопитающих они, соответственно, находятся на X-хромосоме и Y-хромосоме. Все гены, расположенные в этих областях, есть у обоих полов и наследуются так же, как и любые аутосомные гены, отсюда и название областей.

Си́напсис — конъюгация хромосом, попарное временное сближение гомологичных хромосом, во время которого между ними может произойти обмен гомологичными участками. Происходит в профазе I мейоза. Когда гомологичные хромосомы синаптируют, их концы прикрепляются к ядерной оболочке. Потом такие концевые мембранные комплексы перемещаются при содействии ядерного цитоскелета, пока соответственные концы хромосом не объединятся в пары. После этого и межконцевые участки хромосом начинают сближаться, при этом они могут соединяться посредством РНК-белкового комплекса, называемого синаптонемальным комплексом. Синапсису подвергаются только аутосомы в процессе мейоза, половые же хромосомы остаются неспаренными.

Хромосомы типа ламповых щеток, впервые обнаруженные В. Флеммингом в 1882 году, — это специальная форма хромосом, которую они приобретают в растущих ооцитах большинства животных, за исключением млекопитающих.
Хромоме́ры — утолщённые, плотно спирализованные участки нитей ДНК (хромонем), из которых состоит хромосома.

Когези́н — это мультибелковый комплекс, который регулирует процесс разделения сестринских хроматид в ходе деления клетки.

Хиа́зма — в генетике точка, в которой две гомологичные несестринские хроматиды обмениваются генетическим материалом в ходе кроссинговера в течение мейоза. Хиазмы становятся видны в фазе диплотены профазы I мейоза, но сам кроссинговер происходит в предыдущей фазе — пахитене. Когда каждая тетрада, состоящая из двух пар сестринских хроматид, начинает распадаться, хиазмы остаются единственными точками контакта.
Гомологи́чные хромосо́мы — пара хромосом с одинаковым набором генов и сходной морфологии в кариотипе диплоидного организма. Одна из гомологичных хромосом получена при оплодотворении от материнского родительского организма, другая — от отцовского. Гомологичные хромосомы конъюгируют во время профазы первого деления мейоза в процессе кроссинговера.
Синаптоне́мный ко́мплекс (СК) — белковый комплекс, формирующийся между гомологичными хромосомами в ходе мейоза. Синаптонемный комплекс предположительно является связующим звеном между хромосомами во время спаривания (синапсиса).

Пахитена, или пахинема — это одна из стадий профазы первого деления мейоза, называемая также стадией толстых нитей, во время которой завершается формирование синаптонемного комплекса между гомологичными хромосомами, и происходит кроссинговер. Пахитене предшествуют такие стадии профазы I мейоза, как лептотена и зиготена. После неё следует диплотена и диакинез.

SMC5 или белок структурной поддержки хромосом номер пять — это белок, который у человека кодируется геном SMC5.

SMC6 или белок структурной поддержки хромосом номер шесть — это белок, который у человека кодируется геном SMC6.
Балансёрная хромосома — элемент генетического инструментария, специально модифицированная хромосома, используемая для поддержания некоторого гена в искусственной популяции в гетерозиготном состоянии и быстрого поиска подобных особей. Также используется для компенсации повреждённого гена и предотвращения рекомбинации (кроссинговера) между гомологичными хромосомами в мейозе. Наиболее часто используется в генетике Drosophila melanogaster, что позволяет поддерживать в популяции мутацию в состоянии гетерозиготы без постоянного контроля её наличия; также может использоваться для мышей. Балансерная хромосома имеет три важных свойства: она подавляет рекомбинацию с гомологичной хромосомой, несёт в себе доминантные маркеры, негативно влияет на репродуктивную функцию в состоянии гомозиготы.

Контрольная точка мейотической рекомбинации отслеживает мейотическую рекомбинацию во время мейоза и блокирует вход в метафазу I, если рекомбинация не обрабатывается эффективно.
