Бурые водоросли
| Бурые водоросли | |
|---|---|
 Талломы фукоидов Fucus distichus | |
| Научная классификация | |
Домен: Клада: Клада: Клада: Клада: Отдел: Класс: Бурые водоросли | |
| Международное научное название | |
| Phaeophyceae Wettst., 1901 | |
| Порядки | |
| |
Бурые водоросли (лат. Phaeophyceae) — класс охрофитовых водорослей. В жизненном цикле всех представителей присутствуют многоклеточные стадии. Бурые водоросли в хроматофорах содержат бурый пигмент фукоксантин (C42H58O6), который маскирует остальные пигменты.
Бурые водоросли в подавляющем большинстве принадлежат к морским биотопам, их заросли встречаются в литоральной и сублиторальной зонах, до глубин 40—100 м. Представителей класса Phaeophyceae можно увидеть во всех морях земного шара, однако наибольшая их концентрация — в приполярных и умеренных широтах, где они доминируют на глубине от 6 до 15 м. Обычно бурые водоросли растут прикреплённые к скалам и камням разной величины, и только в тихих местах и на большой глубине они могут удерживаться на створках моллюсков или гравии. Среди всего разнообразия бурых водорослей представители только 4 родов — Heribaudiella, Pleurocladia, Bodanella и Sphacelaria — могут обитать в пресных водоемах. Некоторые морские виды могут попадать в прибрежные солоноватые воды, где нередко являются одним из основных компонентов флоры солончаковых болот.
Представители группы играют важную роль в морских экосистемах как один из основных источников органической материи; также, их заросли создают уникальную среду обитания для многих морских организмов. Например, представители рода Macrocystis, которые могут достигать 60 м длиной, формируют большие подводные леса у побережий Америки. Виды рода Sargassum создают окружающую среду тропических вод Саргассова моря, крупнейшего места произрастания бурых водорослей в тропиках. Много видов, за исключением порядка Fucales, можно увидеть в зарослях на подводных скалах вдоль побережья.
Место в системе живого мира
По системе живого мира, которая была предложена S. Adl и коллегами в 2005 году[1], бурые водоросли относятся к царству Chromalveolata Adl et al. отдела Heterokontophyta Hoek класса Phaeophyceae Kjellman.
По системе Adl и коллег, царство Chromalveolata отображает отдельную ветвь развития эукариотических организмов, отделившуюся на древе жизни вследствие исключительного эндосимбиотического события между предковой фотосинтезирующей красной водорослью и фаготрофным эукариотом. С последующей эволюцией некоторые линии живых организмов в царстве Chromalveolata потеряли пластиды, тогда как другие — вторично получили.
Группа Страменопилов объединяет как организмы, способные к фотосинтезу (например диатомей), так и тех, которые потеряли пластиды и питаются гетеротрофно (в частности, это разные группы грибовидных организмов). Всех их объединяет наличие в монадных клетках двух морфологически различных жгутиков. Жгутик, направленный вперёд, — перистый, покрытый двумя рядами специфических трёхчленных волосков — ретронем (мастигонем). Направленный назад жгутик имеет гладкую поверхность. Также общим признаком для всех страменопилов является наличие трубчатых гребней в митохондриях. Кроме класса Phaeophyceae, к группе водорослей-гетероконтов относятся следующие классы водорослей: Chrysophyceae (золотистые), Eustigmatophyceae (эвстигматофициевые), Dictyochophyceae (силикофлагеллаты), Bacillariophyceae (диатомеи), Raphidophyceae (хлоромонады), Xanthophyceae (жёлто-зелёные) и некоторые другие.
Филогенез
Несмотря на то что с начала XX века существовало понимание бурых водорослей как целостной, монофилетической группы по однородному пигментному составу, одинаковым продуктам ассимиляции и схожему строению подвижных стадий, вопрос их филогенеза оставался нерешённым. Сравнивая жгутиковый аппарат и пигментный состав бурых водорослей с золотистыми, Шерффель, а также много других альгологов, высказывались за филогенетическое родство этих двух таксонов[2].
Но уже в конце XX века, благодаря молекулярно-биологическим методам анализа ДНК было установлено, что бурые водоросли в наибольшей степени родственны с группой жёлто-зелёных водорослей (класс Xanthophyceae)[3]. В 1998 году Bailey с коллегами, также основываясь на молекулярных исследованиях ДНК, описал новый небольшой класс водорослей-страменопилов — Phaeothamniophyceae — представители которого, вероятно, и являются прямыми предками бурых водорослей[4].
Бурые водоросли являются сравнительно молодой группой организмов. По разным данным, время их возникновения — от 150[5] до 200[6] миллионов лет назад. С точки зрения эволюции, бурые водоросли являются уникальной группой живых организмов, поскольку они принадлежат к тому небольшому количеству линий развития эукариотов, которым, независимо от других, удалось развить настоящий многоклеточный план строения тела.
Классификация
По современной системе, основываясь на молекулярных, морфологических, онтогенетических критериях, а также на основе различий в ультраструктурном строении клеток, в классе Phaeophyceae выделяют 18 порядков[7]:
| Порядок | Количество видов |
|---|---|
| Ectocarpales Bessey | 693 |
| Fucales Bory de Saint-Vincent | 524 |
| Dictyotales Bory de Saint-Vincent | 239 |
| Laminariales Migula | 130 |
| Sphacelariales Migula | 82 |
| Ralfsiales Nakamura ex Lim & Kawai | 54 |
| Desmarestiales Setchell & Gardner | 33 |
| Sporochnales Sauvageau | 29 |
| Cutleriales Bessey | 11 |
| Tilopteridales Bessey | 9 |
| Scytothamnales Peters & Clayton | 8 |
| Onslowiales Draisma & Prud’homme van Reine | 4 |
| Syringodermatales Henry | 4 |
| Asterocladales Silberfeld et al. | 3 |
| Discosporangiales Schmidt | 3 |
| Ishigeales Cho & Boo | 3 |
| Ascoseirales Petrov | 1 |
| Nemodermatales Parente et al. | 1 |
Типы морфологических структур тела и строение талломов
В пределах класса представлены нитчатый, разнонитчатый и настоящий тканевый типы структур тела водорослей. Все бурые водоросли, за исключением некоторых видов рода Sargassum, ведут прикреплённый образ жизни. Органами прикрепления служат ризоиды или специализированные базальные диски.
У представителей с нитчатой структурой тела талломы созданы системой однорядных разветвлённых нитей. Разнонитчатые водоросли имеют вид довольно толстых шнуров. Такие шнуры имеют одно- или многоосевое строение. Среди бурых водорослей есть представители с эфемерными, однолетними и многолетними талломами. Значительное влияние на продолжительность существования таллома имеют экологические факторы.

Разнообразная форма талломов характерна для представителей с тканевым типом строения тела. Форма талломов таких водорослей может быть в виде кожуры, шарообразной, мешкоподобной, пластинчатой, в виде куста с ребристыми листообразными пластинками. Талломы некоторых представителей могут иметь специальные воздушные пузырьки — пневматофоры, которые удерживают «ветви» в вертикальном положении. В простейших случаях таллом сформирован двумя тканями: корой, которая образована мелкими клетками с окрашенными хлоропластами, и сердцевиной, которая состоит из больших бесцветных клеток, которые выполняют проводящую и запасающую функции.
У более высокоорганизованных представителей таллом образован четырьмя типами тканей: корой, меристодермой, промежуточной тканью и сердечником. Меристодерма — поверхностная ткань, которая выполняет защитную и репродуктивную функции. Её клетки обычно мелкие и способны к активному делению. В клетках промежуточной ткани накапливаются продукты ассимиляции.
Типы роста
Для талломов бурых водорослей характерно 6 типов роста:
- Диффузный — большинство клеток растительного организма способны к делению (напр., Ectocarpus, Petalonia).
- Верхушечный — к делению клетки способны только на верхушках талломов (напр., Sphacelaria, Dictyota).
- Трихоталлический — клетки делятся, формируя волоски над и под поверхностью талломов (напр., Cutleria, Desmarestia).
- Промеристематический — апикальная клетка не способна к делению, но контролирует разделение меристематических клеток, расположенных под ней (напр., Fucus).
- Интеркалярный — в талломе имеется меристематическая зона, клетки которой, делясь, нарастают вверх и вниз от неё (напр., Laminaria).
- Меристодерматический — рост обеспечивается специализированной поверхностной тканью, клетки которой делятся параллельно и перпендикулярно поверхности таллома (напр., Sargassum).
Цитологические и биохимические особенности строения клеток
Клеточные покровы бурых водорослей образованы оболочками, которые состоят из внутреннего каркасного слоя, образованного микрофибрилами целлюлозы, и внешнего аморфного, который содержит в основном соли альгиновой кислоты, пектиновые вещества и белки. Соли альгиновой кислоты — альгинаты — в водных растворах способны к образованию гелей.

(af) передний жгутик; (c) хлоропласт; (e) глазок; (fh) ретронемы (присутствуют вдоль всего жгутика); (fs) базальное утолщение заднего жгутика; (g) комплекс Гольджи; (li) липидное включение; (m) митохондрии; (mb) микротельце; (mt) микротрубочки; (n) ядро; (p) пиреноид; (pf) задний жгутик; (v1) физоды; (v2) запасной продукт; (v3) везикулы со строительным материалом клеточной оболочки
Ещё одним компонентом клеточных оболочек бурых водорослей являются фукоиданы. По своей химической природе они являются сульфатированными полисахаридами сложного строения. Основной мономер фукоиданов — этерифицированные серной кислотой остатки α-L-фукозы. Кроме неё, в состав фукоиданов могут также входить и другие моносахариды. Подобные полисахариды не встречаются ни в одной другой группе водорослей, однако похожие на них соединения были обнаружены у морских беспозвоночных (морские ежи и голотурии)[8].
Благодаря большому количеству альгинатов и пектиновых веществ, клеточные оболочки бурых водорослей способны сильно слизьневеть и находиться в коллоидном (гелеобразном) состоянии.
Клетки бурых водорослей одноядерные. Ядро имеет типичное эукариотическое строение, внутри него имеется большое, хорошо заметное ядрышко. Как и у многих других классов водорослей-страменопилов, которым родственны бурые водоросли, внешняя оболочка ядра переходит в хлоропластный эндоплазматический ретикулум (ЭПР).
Хлоропласты мелкие, многочисленные. Имеют дискообразный вид. У большинства бурых водорослей пиреноиды в хлоропластах очень мелкие, почти незаметные в световой микроскоп. Пластиды бурых водорослей вторично-симбиотические, родофитного типа, окружённые четырьмя мембранами. Две внешние образуют хлоропластный ЭПР, две внутренние являются собственно мембранами пластиды. Между внешними и внутренними мембранами сохраняется перипластидное пространство. Тилакоиды в пластиде преимущественно располагаются лентами по три, на периферии расположена опоясывающая ламелла.
Пигменты пластид являются хлорофиллами а и c, β— и ε-каротинами, а также специфическими ксантофиллами — фукоксантином и виолаксантином. Именно последние и обуславливают характерный буро-желтый цвет талломов водорослей из класса Phaeophyceae. Кроме вышеуказанных соединений, в хлоропластах бурых водорослей также присутствуют другие ксантофиллы: диатоксантин, диадиноксантин, зеаксантин, антераксантин и неоксантин.
За пределами хлоропласта, в виде цитоплазматических пластинок, окружённых собственными мембранами, откладывается основной запасной продукт бурых водорослей — специфический полисахарид ламинарин. Кроме ламинарина, запасным продуктом у бурых водорослей может служить шестиатомный спирт D-маннитол. Так, у некоторых видов ламинарий его концентрация осенью может достигать до 25 % сухой массы.
Ещё одной особенностью бурых водорослей является наличие в их клетках особых вакуолей — физод. В световой микроскоп физоды выглядят как небольшие светопреломляющие включения, в молодых клетках — бесцветные, у стареющих — жёлтого или бурого цвета. Диаметр физод может составлять от 0,1 до 10 мкм, в среднем — 1-4 мкм. На сегодня достоверно установлено, что содержимое физод является совокупностью полифенольных соединений отдельного класса танинов, производных флороглюцина — флоротанинов.
Жгутиковые стадии у бурых водорослей являются исключительно гаметами и зооспорами (у бурых водорослей отсутствуют движущиеся вегетативные клетки). Такие клетки имеют два жгутика, как и остальные страменопилы. Длинный жгутик — порчатый, покрытый трёхчленными мастигонемами, а короткий — гладкий. Также у монадных клеток, в хлоропласте имеется глазок (стигма), состоящий из 40-80 липидных глобул. Глазок функционирует как вогнутое зеркало, фокусируя свет на основе жгутика, где содержится фоторецептор, который отвечает за фототаксисы гамет или зооспор бурых водорослей.
Размножение и жизненный цикл
У бурых водорослей наблюдаются все типы размножения: вегетативное, бесполое, а также половое. Вегетативное размножение происходит при случайной фрагментации талломов, и только род Sphacelaria формирует особые выводковые почки.
Неполовое размножение подвижными зооспорами свойственно подавляющему большинству бурых водорослей, однако у представителей порядков Dictyotales и Tilopteridales можно наблюдать размножение неподвижными тетра- и моноспорами.
Местом образования зооспор служат одно- или многогнёздные зооспорангии. Это значительно увеличенные в размерах клетки, содержимое которых, после серии разделов ядра, распадается на большое количество зооспор. Обычно первое деление ядра является редукционным, поэтому новообразованные зооспоры имеют гаплоидный набор хромосом. Попав в окружающую среду, зооспоры переходят к активному движению, но уже через несколько минут они оседают на субстрат и сбрасывают жгутики. Часть зооспор прорастает в мужской гаметофит, часть — в женский.
Половой процесс у бурых водорослей представлен изо- и оогамией, изредка случается гетерогамия.

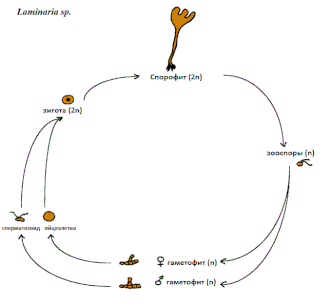
Для всего многообразия бурых водорослей, кроме представителей порядка Fucales, характерен жизненный цикл с чередованием гаплоидного (гаметофит) и диплоидного (спорофит) поколений. Тип жизненного цикла, в котором гаметофит морфологически не отличается от спорофита, называют изоморфным. В гетероморфном жизненном цикле гаметофит бурых водорослей обычно имеет микроскопические размеры и выглядит как система разветвлённых нитей. Продуктами гаметофита являются мужские и женские гаметы, которые сливаясь, дают начало новому спорофиту.
Водоросли из порядка Fucales имеют диплофазный жизненный цикл, без смены поколений. У них отсутствуют зооспоры, мейоз происходит при образовании гамет, а половой процесс представлен преимущественно оогамией. Мужские и женские гаметангии образуются на специализированных плодовитых окончаниях талломов — рецептакулах. Внутри рецептакулов содержатся большие полузакрытые полости, где развиваются мужские антеридии и женские оогонии. Созревшие оогонии и антеридии высвобождают яйцеклетки и сперматозоиды в окружающую среду, где и происходит процесс оплодотворения. Зигота, которая образовалась, сразу же начинает делиться и прорастать в новый диплоидный таллом.
Важную роль в половом размножении бурых водорослей играют феромоны. У бурых водорослей они выполняют две важные функции. Во-первых — это стимуляция высвобождения мужских гамет, а во-вторых, они ответственны за привлечение сперматозоидов к женским гаметам или яйцеклеткам. По химической природе — это летучие, гидрофобные, линейные или циклические непредельные углеводороды. На сегодня выявлено около десятка феромонов в разных родов и видов бурых водорослей.
Охрана бурых водорослей
По состоянию на 2013 год в Красном списке Международного союза охраны природы (МСОП) есть 15 видов бурых водорослей. Все водоросли, занесённые в список МСОП, растут у побережья Галапагосских островов. Четыре из них получили наивысшую категорию уязвимости «В критической опасности». По одному виду имеют категории «В опасности» и «В уязвимом положении», 9 видов получили категорию «Виды с неясным статусом», что свидетельствует о недостаточной исследованности динамики популяций этих видов. МСОП определяет климатические изменения (в частности, температурный феномен Эль-Ниньо) и связанное с ними чрезмерное размножение организмов-фитофагов как основные негативные факторы, влияющие на численность популяций бурых водорослей[9].
Значение для человека
Издавна жители побережий, особенно в Восточной Азии, употребляли их в пищу, часто как основу рациона; сегодня бурые водоросли являются, прежде всего, сырьем для получения альгинатов, используемых в пищевой, текстильной, фармацевтической и биотехнологической отраслях промышленности. В перспективе бурые водоросли и их компоненты также рассматриваются как источник новых лекарств[10].

Пищевая ценность
Как пищевые продукты наибольшей популярностью в некоторых странах (Китай, Япония и Южная Корея) пользуются водоросли комбу (Saccharina japonica) и вакамэ (Undaria pinnatifida). Эти два вида бурых водорослей издавна являются объектами массовой аквакультуры в Китае и Южной Корее. Кроме вышеуказанных видов в Восточной Азии употребляются также водоросли хиджики (Sargassum fusiforme) и араме (Eisenia bicyclis)[11].
С точки зрения диетологии, бурые водоросли — низкокалорийная еда с низким содержанием жиров, обогащённая углеводами, белками и, особенно, минералами. По сравнению с обычными овощами и фруктами, бурые водоросли имеют значительно большее содержание макро- и микроэлементов, таких как натрий, калий, кальций, магний, железо, селен, кобальт, марганец[12]. Также бурые водоросли являются одним из главных источников йода, недостаток которого в организме человека приводит к серьёзным нарушениям развития, снижению трудоспособности, кретинизму. В последнее время доказано, что употребление бурых водорослей и их производных является одним из лучших методов групповой и индивидуальной профилактики йододефицита и йододефицитных заболеваний[13]. Как пищевой продукт, бурые водоросли богаты клетчаткой, в роли которой выступают коллоидные компоненты их клеточных оболочек — альгинаты. Соли альгиновой кислоты также имеют значительные радиационнозащитные свойства[14]. Исследования также показывают, что регулярное употребление бурых водорослей в пищу снижает инсулинорезистентность и, как следствие, риск развития диабета 2-го типа[15].
Значение для науки
В 1907 году японский биохимик Кикунэ Икэда, исследуя вкусовые свойства традиционных блюд японской кухни, приготовленных из бурых водорослей комбу, описал новый тип вкусовых ощущений — умами. Из 40 кг Saccharina japonica ученый выделил 30 г глутаминовой кислоты, которая оказалась ответственной за характерный вкус. В 1908 году компания Икэды начала промышленный выпуск новой приправы — глутамата натрия[16].
В 2010 году международной командой учёных был секвенирован геном бурой водоросли Ectocarpus siliculosus. Благодаря этому стало возможным использовать этот организм как модельный объект для изучения вопросов, связанных с биологией бурых водорослей, в частности возникновения многоклеточности[17].
См. также
Примечания
- ↑ Adl, S. M.; et al. The new higher level classification of eukaryotes with emphasis on the taxonomy of protists (англ.) // Journal of Eukaryotic Microbiology : journal. — 2005. — Vol. 52, no. 5. — P. 399—451. — doi:10.1111/j.1550-7408.2005.00053.x. — PMID 16248873. Архивировано 29 мая 2013 года.
- ↑ Д. К. Зеров. Очерк о филогении бессосудистых растений. — К.: Научная мысль, 1972. — 316 с.
- ↑ Ariztia, E. V.; Andersen, R. A.; Sogin, M. L. A new phylogeny of chromophyte algae using 16S-like rRNA sequences from Mallomonas papillosa (Synurophyceae) and Tribonema aequale (Xanthophyceae) (англ.) // Journal of Phycology[англ.] : journal. — 1991. — Vol. 27, no. 3. — P. 428—436. — doi:10.1111/j.0022-3646.1991.00428.x.
- ↑ Bailey, J.C., R.R. Bidigare, S.J. Christensen and R.A. Andersen. 1998. Phaeothamniophyceae classis nova.: a new lineage of chromophytes based upon photosynthetic pigments, rbcL sequence analysis and ultrastructure. Protist 149: 245—263.
- ↑ Medlin, L. K.; et al. Phylogenetic relationships of the 'golden algae' (haptophytes, heterokont chromophytes) and their plastids (англ.) // Plants Systematics and Evolution : journal. — 1997. — Vol. 11. — P. 187—219. Архивировано 5 октября 2013 года.
- ↑ Lim, B.-L.; Kawai, H.; Hori, H.; Osawa, S. Molecular evolution of 5S ribosomal RNA from red and brown algae (англ.) // Japanese Journal of Genetics : journal. — 1986. — Vol. 61, no. 2. — P. 169—176. — doi:10.1266/jjg.61.169. Архивировано 10 января 2015 года.
- ↑ Guiry, M.D. & Guiry, G.M. 2013. AlgaeBase Архивная копия от 12 октября 2009 на Wayback Machine. World-wide electronic publication, National University of Ireland, Galway
- ↑ А. И. Усов, М. И. Билан. Фукоиданы — сульфатированые полисахариды бурых водорослей. // Успехи химии. — 2009. Том 78. — с. 846—862.
- ↑ Сайт Красного списка МСОП (недоступная ссылка)
- ↑ Albertus J. Smith. Medicinal and pharmaceutical uses of seaweed natural products: A review. // Journal of Applied Phycology. — 2004. Vol. 16. — pp. 245 − 262.
- ↑ Dennis J. McHugh. A Guide to the Seaweed Industry. — Food and Agriculture Organization of the United Nations, 2003. — 106 p.
- ↑ P. Ruperez. Mineral content of edible marine seaweeds. // Food Chemistry. — 2002. No. 79. — pp. 23 — 26.
- ↑ В. Н. Корзун, А. М. Парац, А. П. Матвиенко. Проблемы и перспективы профилактики йоддефицитных заболеваний у населения Украины. // Эндокринология. — 2006. Т.11, № 2. — с. 187—193.
- ↑ В. Н. Корзун, В. И. Сагло, А. М. Парац. Питание в условиях широкомасштабной аварии и её последствиях. // Украинский медицинский журнал. — 2002. № 6 (32). — с. 99 — 105.
- ↑ Brown seaweed emerges as a potent ally in diabetes control (англ.). News-Medical.net (4 декабря 2023). Дата обращения: 6 декабря 2023. Архивировано 5 декабря 2023 года.
- ↑ Вещество с умами: вкус. — Популярная механика. Дата обращения: 22 января 2014. Архивировано из оригинала 30 января 2014 года.
- ↑ J. M. Cock et al. The Ectocarpus genome and the independent evolution of multicellularity in brown algae. // Nature — 2010. Дата обращения: 22 января 2014. Архивировано 5 октября 2013 года.
Литература
- Белякова Г. А. Водоросли и грибы // Ботаника: в 4 тт. / Белякова Г. А., Дьяков Ю. Т., Тарасов К. Л. — М.: Издательский центр «Академия», 2006. — Т. 2. — 320 с. — 3000 экз. — ISBN 5-7695-2750-1.
Ссылки
- Бурые водоросли // Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров. — 3-е изд. — М. : Советская энциклопедия, 1969—1978.