Гаплогруппа I1 (Y-ДНК)
| Гаплогруппа I1 | |
|---|---|
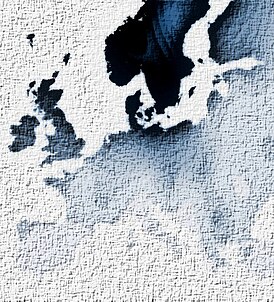 Распространение гаплогруппы I1 Распространение гаплогруппы I1 | |
| Тип | Y-ДНК |
| Время появления | ~ 25.500 до н.э. |
| Место появления | Европа |
| Предковая группа | I |
| Сестринские группы | I2 |
| Мутации-маркеры | M253, M307.1/P203.1, M450/S109, P30, P40, S62, S63, S64, S65, S66, S107, S108, S110, S111 |
В человеческой генетике Гаплогруппа I1 (бывшая I1a) (M253, M307.1/P203.1, M450/S109, P30, P40, S62, S63, S64, S65, S66, S107, S108, S110, S111) — Y-хромосомная гаплогруппа, типичная для популяций Скандинавии и северо-западной Европы, с умеренным распределением всюду по Восточной Европе.
Происхождение
Гаплогруппа I1 происходит от мутации гаплогруппы I, произошедшей у мужчины, жившего ок. 27 500 лет назад. Последний общий предок современных носителей гаплогруппы I1 жил 4600 л. н. (даты определены по снипам компанией YFull[1]).
Гаплогруппа I1 является коренной европейской гаплогруппой[2]. Большинство современных носителей гаплогруппы I1 составляют носители германских языков индоевропейской семьи, хотя первоначально данная гаплогруппа была связана, по-видимому, не с индоевропейскими народами, а с догерманским субстратом.
I1-V5203/Z2744 идентифицируется по меньшей мере 300 уникальными мутациями, что означает, что данная группа либо была совершенно изолирована в течение долгого периода (что маловероятно), либо пережила серьёзное «бутылочное горлышко» в сравнительно недавнем времени.
Субклад I1a-L121 образовался 4600 лет назад. Он имеет три основные сублинии: I1a-CTS12768, I1a-Z17954, I1a-DF29[3].
Субветвь I1a1b1a4a1a1-L258 (ранее I1d-Bothnia, I1a1b3a1) характерна для финнов (16,8%)[4], I1a3-Z63 — для Восточной Европы, также как I1a-Z2337I1a-Z2337 и I1a-L22.
Палеогенетика
- I1 определили у представителя азильской культуры BAL0051 из Balma Guianya в Пиренеях (испанская провинция Льейда)[5].
- Гаплогруппа I1 была обнаружена у мезолитического обитателя острова Стура-Карлсё (Лен Готланд, Швеция)[6].
- Гаплогруппа I1 была обнаружена у представителя культуры линейно-ленточной керамики (неолит Венгрии), жившего ок. 7000 лет назад[7][8].
- Как минимум два представителя бронзового века Швеции имели гаплогруппу I1 (Angmollan, 1493—1302 года до н. э.)[9]
- I1a2a1a1a-Z140 определили у образца CGG106515 (Strandlunden II Gerlev MFG 191/02, EarlyBronzeAge, мтДНК: V10) из Зеландии (Дания)[10]
- Представитель Вельбарской культуры (Польша), живший примерно в 100—300 году н. э., имел гаплогруппу I1[11].
- I1a2a1a1a-Z140 определили у образца CGG106734 (Ods Trundholm Mose, IronAge, мтДНК: T2b9) из Зеландии (Дания)[10]
- Англосакс, живший примерно в 550—650 годах н. э. и похороненный в Северной Британии, имел гаплогруппу I1[12]
- Мужчина из захоронения лангобардов в западной Венгрии, живший примерно в 600—650 годах н. э., имел гаплогруппу I1[13]
- I1-M253 определили у образцов VIII века из корабля «Салме-2»[англ.] (волость Сальме, Эстония)[14]
- Житель средневековой Польши, живший примерно в 1000—1200 гг. н. э., имел гаплогруппу I1 (захоронен вместе с оружием и ценными вещами)[11]
- I1a определили у жителя Гнёздова X—XI веков[14]
- У жителей Старой Ладоги X—XII веков определили Y-хромосомные гаплогруппы I (n=1), I1 (n=4), I1a1 (n=1), I1a1b1 (n=2)[14]
- Гаплогруппа I1 была обнаружена в шести образцах из средневекового захоронения в Исландии. Субклады: I1a2a1a2-F2642, I1b-Z131, I1-M253, I1a1b3b-L813, I1a1b3-Z74, I1-M253[15]
- I1a2a1a1d1b1~-Y4870>Y4870*[16] определили у образца KRA004 из Кракауэр-Берга (Krakauer Berg) в Восточной Германии, датирующегося возрастом примерно 612 л. н. (666—558 л. н.)[17]
- I1a2-Z58 определили у образца № 26 из захоронения в Патриаршем саду Владимира у реки Клязьма, датируемого XIII—XIV веками[18].
Субклады
Субклады[19]
- Z131 + является небольшим субкладом, который был найден в районах, граничащих с древней кельтско-германской границей (Бельгия, Центральная Германия, Богемия), также обнаружен в Швеции и Великобритании.
- Z17925 + небольшой субклад, который можно найти в Германии, Франции и Англии.
- Z19086 + является небольшим субкладом, найденным в Финляндии.
- DF29 + представляет 99% линий I1.
- • CTS6364 + основной северный субклад, вероятно скандинавского происхождения.
- • • Y3866 + встречается в основном в Скандинавии.
- • • • A5338 + встречается чаще всего в Норвегии, но также был найден в Шотландии.
- • • • S11221 + встречается чаще всего в Швеции и Норвегии, но также найдена в Венгрии.
- • • • S4767 + образовался около 4000 лет назад.
- • • • • Y4781 + встречается в основном в скандинавских странах.
- • • • • • M227 + встречается почти во всех странах Европы, и даже на Ближнем Востоке.
- • • • • S7642 + встречается в основном в Швеции.
- • • • • • Y18103 + встречается в основном в Польше.
- • • L22 + (aka S142 +) — очень большая скандинавская ветвь. Имеет очень большое распространение в Великобритании, особенно на восточном побережье, где селились датские викинги, а также в Польше и России (шведские викинги). L22 (I1a1b) характерна для части башкир-сары-мингов и башкир-юрматов
- • • • P109 + В основном южный скандинавский субклад, с присутствием во всех регионах, куда мигрировали датские викинги. Он был обнаружен во многих частях Европы, таких как западная Иберия, северная Италия, Балканы, Литва и Россия.
- • • • • S10891 + встречается в основном в Швеции и Норвегии, но также в Нормандии и Британских островах (происхождение — викинги), а также в северной Италии (субклад FGC21732).
- • • • • S7660 / Y3662 +
- • • • • • Y4045 + находится в Англии и Ирландии (происхождение — викинги).
- • • • • • S14887 + встречается в основном в Дании, Швеции и Финляндии.
- • • • • • • FGC22045 + является основным субкладом I1, найденным на Балканах. Вероятно, норманнского происхождения.
- • • • • Y3664 + незначительный субклад, найденный в Дании, Шлезвиг-Гольштейне, Нормандии и Гернси (происхождение — викинги).
- Y5621 + — незначительная клада, найденная в Швеции, Нормандии, Великобритании и России (происхождение викингов).
- Y14999 + — незначительная клада, найденная во Франции и Великобритании.
- L205 + в основном ограничивается Нидерландами, Францией и Великобританией. Отдельные случаи были также выявлены в Швеции и Испании.
- L287 + является подавляющим финским подклассом (в целом по всей стране), с очень небольшим присутствием в Норвегии, Швеции, Польше и России.
- L300 + — это небольшой субклад, который встречается почти исключительно на юге Финляндии.
- L813 + является преимущественно скандинавским субкладом, особенно распространенным в южной Норвегии, но также найденным в Швеции и в меньшей степени западной Финляндией. Он также встречается в Великобритании (вероятно, спуск викингов) и в северных Нидерландах (но не в Германии).
- Z58 + преимущественно западногерманский, с очень сильным присутствием в Германии, странах с низким и британским. Он также находится в меньшей степени в скандинавских странах и во всей континентальной Европе. Его возраст оценивался приблизительно за 4600 лет до настоящего времени.
- Z138 + (aka Z139 +) — очень разрозненный субклад. Он встречается на очень низкой частоте во всем германском мире, с пиком в Англии и Уэльсе (хотя это может быть просто из-за передискретизации в Британии). Помимо германских стран, он также был найден в Ирландии, Португалии, на юге Италии, Венгрии и Румынии. Z138 + соответствует AS2, AS10, AS1010.2, AS10910, AS1221, AS1414 и Esc-13 в номенклатуре STR на основе FTDNA. Z138 разделен на четыре подклада: S2293 (самый большой), S5619, PF1610 и PF2364.
- S2293 + был образован 4500 лет назад и сразу же разделен на две ветви: S6277 и Z2541. S6277 + находится в Скандинавии, Великобритании, Франции, Германии, Австрии и Италии (Турин, Матера, Реджо-ди-Калабрия).
- Z2541 + соответствует AS1313 в номенклатуре STR на основе FTDNA. Это относительно небольшая клада, распространенная особенно на северо-западе Европы, но также на западе Украины, Румынии и Португалии. Он появился 4,500 лет назад, но имеет TMRCA 3800 лет. Он имеет пять подкладок:
- S2268 + находится в Ирландии, Великобритании, Нидерландах, Германии, Дании, Норвегии, Австрии и Венгрии.
- S19185 + находится в Великобритании, Ирландии, Норвегии и Венгрии.
- Y7043 + — самый большой из субкладов Z2541. Он находится в Норвегии на севере Италии (возможно, Ломбардская связь) через Германию, Бельгию и Швейцарию.
- Y21391 + был найден в Люксембурге. Y29668 + — небольшая клада, найденная в Англии.
- Z59 + является главной ветвью Z58.
- Z2040 + сформировался 4300 лет назад и разбился на две клады около 3900 лет назад. Образцы, отрицательные для L1450 и Z382, были найдены в Норвегии, Финляндии и России.
- L1450 + — небольшая клада, разбросанная по Норвегии, Германии, Польше, Болгарии, Словении, Швейцарии, Великобритании и Ирландии.
- Z382 + наиболее распространен в Южной Швеции, Дании и на Британских островах, встречается в Голландии и Германии, с небольшим присутствием в Норвегии. Отдельные образцы были также идентифицированы в России, Украине, Финляндии, Польше, Франции, Италии, Хорватии и Румынии. Соответствует AS3, AS3-911, AS13 и Sw в номенклатуре STR на основе FTDNA.
- S26361 + встречается в основном в Швеции и Норвегии.
- S16414 + был найден в Норвегии и на Сардинии (YP5417 +, TMRCA 1750 ybp), где он мог быть принесен вандалами.
- Y2170 + наиболее распространен в Южной Швеции, присутствует в Германии, Великобритании и России.
- Y5384 + находится в Дании, Финляндии и Италии (Рим).
- Z60 + встречается во всем германском мире
- Y12342 + — небольшая клада, найденная в основном в Финляндии, но также и в Англии.
- Y22033 + — незначительная клада, найденная в Шотландии.
- Z140 + является явно западногерманским подкладом, который был найден в основном на Британских островах, Нидерландах (одна треть всех I1), северной Франции, центральной и южной Германии и Швейцарии. В скандинавских странах очень редок. Изолированные образцы были найдены в Испании, центральной и южной Италии, Словении, Чехии, Польше, Украине и России. Z140 * соответствует гаплотипам AS5, AS6, AS814 и EE в номенклатуре STR на основе FTDNA. I1-Z140 характерна для башкир-мурзалар (улаште) и тырнаклинцев, части кудейцев. Башкирские линии I1-Z140 отделились от европейских линий I1-Z140 более 2 тыс. лет назад.
- A196 + находится в Швеции, Германии, Швейцарии, Австрии, Венгрии, Франции, Португалии, Англии, Уэльсе, Шотландии, Ирландии, Беларуси и России.
- A1605 + — небольшая клада, найденная в Нидерландах и Великобритании. BY477 / Y15150 + — небольшая клада, найденная в Германии, Великобритании, Ирландии, Португалии и Испании.
- CTS8691 / F2642 / S2169 + — большая клада, особенно в Скандинавии, Германии и Великобритании, но также в Финляндии, Нидерландах, Швейцарии, Польше и Ирландии.
- Y6232 + находится в Центральной Европе, Великобритании и Италии.
- Y7277 + можно найти в Германии, Нидерландах, Англии, Чехии, Швейцарии и Италии. Этот субклад может быть связан с лангобардами.
- Y10890 + встречается особенно в центральной и южной Италии недалеко от мест, расположенных на Ломбардах (Ортона, Алифае, Кампобассо) и Вандалах (Трапани).
- Z2535 + — очень большая клада, найденная в Скандинавии, Великобритании, Германии, Чехии, Польше и Румынии.
- L338 + — большой субклад, найденный в Германии, Нидерландах, Норвегии, Великобритании, Швейцарии, Польше и Румынии. Обычно он имеет значение STR GATA-H4 = 9.
- L338 + делится на несколько подкладок, которые соответствуют AS1, AS1H, AS8, AS114 и AS11616 в номенклатуре STR на основе FTDNA.
- Z73 + (и L1301 +) главным образом северный скандинавский и финский. Он также встречается в России, на восточном побережье Великобритании и на шотландских островах (наследие викингов). Z73 + соответствует AS9 и AS16 в номенклатуре STR на основе FTDNA.
- L1302 + встречается главным образом в Швеции, с несколькими субкладами в Финляндии и Норвегии (BY495) и несколькими отдельными образцами, отобранными в Шотландии и России.
- L573 +, по-видимому, является главным образом северо-восточным германским подкладом, но он также был найден в Бельгии, Франции, Швеции, Польше, Литве и на северо-западе России.
- L1248 + является второстепенным подкладом, который был найден в Швеции, Великобритании, Германии и России.
- L803 + был идентифицирован только в Шотландии.
- Z63 + является сильно континентальным германским подкладом, практически отсутствующим в скандинавских странах. Это наиболее распространено в Центральной Германии, Бенилюксе, Англии, Нижней Шотландии, а также в Польше. Он также был найден в России, на Украине, на Балканах, в Италии, Испании и Португалии.
- S2078 + — широко распространенная клада, найденная в большинстве стран Европы, но относительно редко встречается в скандинавских странах, в Бенилюксе и на Британских островах.
- Y2245.2 + / PR683 составляет большую часть Z63 в России, Украине, Польше, на Балканах, в Италии и в Иберии. Он также находится в Великобритании, Швеции, Франции, Германии, Чехии, Боснии, Албании и Ливане. Некоторые субклады могли быть распространены готами.
- L1237 + находится в Швеции, Польше, Украине, России, Сербии, Боснии, Чехии, Франции и на севере Италии. Может быть готического происхождения. Подклас A8249 встречается главным образом в Великобритании, в то время как очень молодой (850 ybp) подклассу Y8815 находится исключительно в Шотландии.
- Y3968 + / S10360 + находится в Скандинавии, Финляндии, Германии, Англии, Польше, Украине, России, Венгрии, Италии, Испании и Португалии. Не только готика, но, несомненно, найден среди готов.
- Y7234 + / FGC14480 + находится в Польше, Германии, Бельгии, Франции и Испании.
- S2097 / Y4102 +> FGC29230 + встречается особенно в Испании (Каталония, Мадрид, Эстремадура) и Италии (Ломбардия, Сицилия), но и в Германии. Может быть готического происхождения.
- Y6375 + находится в Великобритании.
- Y16435 + находится в Сербии (второй наиболее распространенный подкласс после FGC22045).
- Y24458 + V68 + находится в Беларуси, Сербии и Косово.
- BY351 + — это подкласс, который можно найти в Польше, Украине, Хорватии, Италии (включая Сардинию), Франции, Испании и Португалии. Вероятно, он было распространен вестготами и остготами.
Распространение
Наиболее высокие частоты I1 обнаружены в Скандинавии. Анализ генетического разнообразия указывает на территорию современной Дании и Северной Германии как на место происхождения предка I1[20].
В регионах России до 18 % (Roewer 2008) носителей гаплогруппы I1. Вероятно, что они представляют собой потомков дофинского и доиндоевропейского населения прибалтийского региона; также возможно, что это потомки скандинавских или германских переселенцев, а также потомки ассимилированных остготов, проживавших в Крыму с IV века н. э.
Частоты гаплогруппы I1 в европейской части России.
| Популяция | Частота | Публикация |
|---|---|---|
| Вологодская обл. | 18 % | (Roewer 2008) |
| Архангельск | 14,2 % | (Mirabal 2009) |
| Рязанская обл. | 14 % | (Roewer 2008) |
| Казанские татары | 13,2 % | (Trofimova 2015[21]) |
| Красноборск (Арханг.) | 12,1 % | (Balanovsky 2008) |
| Мокша | 12 % | (Rootsi 2004)* |
| Вологда | 11,6 % | (Balanovsky 2008) |
| Унжа (Костр.) | 11,5 % | (Balanovsky 2008) |
| Кострома | 11,3 % | (Underhill 2007)* |
| Пензенская обл. | 11 % | (Roewer 2008) |
| Ивановская обл. | 10 % | (Roewer 2008) |
| Тамбовская обл. | 10 % | (Roewer 2008) |
| Лезгины | 9 % | (Yunusbayev 2000) |
| Карелы | 8,6 % | (Underhill 2007) |
| Ливны (Орл.) | 8,2 % | (Balanovsky 2008) |
| Тверь | 7,9 % | (Mirabal 2009) |
| Архангельск | 7,6 % | (Underhill 2007) |
| Чуваши | 7,5 % | (Rootsi 2004) |
| Архангельская обл. | 7 % | (Roewer 2008) |
| Брянская обл. | 7 % | (Roewer 2008) |
| Остров (Пск.) | 6,7 % | (Balanovsky 2008) |
| Костромская обл. | 6 % | (Rootsi 2004) |
| Псков | 5,4 % | (Underhill 2007) |
| Русские Адыгеи | 5,1 % | (Rootsi 2004) |
| Тверская обл. | 5 % | (Roewer 2008) |
| Курск | 5 % | (Mirabal 2009) |
| Казаки | 4,5 % | (Underhill 2007) |
| Пристень (Курск.) | 4,4 % | (Balanovsky 2008) |
| Куб. казаки (Адыг.) | 4,4 % | (Balanovsky 2008) |
| Казаки | 4,1 % | (Rootsi 2004) |
| Русские Башкирии | 4 % | (Rootsi 2004) |
| Липецкая обл. | 4 % | (Roewer 2008) |
| Коми | 3,6 % | (Rootsi 2004) |
| Порхов (Пск.) | 3,5 % | (Balanovsky 2008) |
| Белгород | 3,5 % | (Balanovsky 2008) |
| Белгородская обл. | 3,5 % | (Rootsi 2004) |
| Репьевка (Воронеж.) | 3,1 % | (Balanovsky 2008) |
| Воронеж | 3,1 % | (Underhill 2007) |
| Новгородская обл. | 3 % | (Roewer 2008) |
| Кашин (Твер.) | 2,7 % | (Balanovsky 2008) |
| Вепсы | 2,6 % | (Underhill 2007) |
| Смоленская обл. | 2 % | (Roewer 2008) |
| Орловская обл. | 2 % | (Roewer 2008) |
| Рославль (Смол.) | 1,9 % | (Balanovsky 2008) |
| Смоленск | 1,9 % | (Underhill 2007) |
| Смоленская обл. | 1,7 % | (Rootsi 2004) |
| Пинега (Арх.) | 0,9 % | (Balanovsky 2008) |
| Пинега (Арх.) | 0,8 % | (Rootsi 2004) |
| Татары | 0,8 % | (Rootsi 2004) |
| Мезень (Арх.) | 0 % | (Balanovsky 2008) |
- В работе Rootsi 2004 и Underhill 2007 выборка частично собрана из материала лаборатории Балановских.
Известные люди
- Ярл Биргер (Биргер Ма́гнуссон) — правитель Швеции из рода Фолькунгов, ярл Швеции с 1248 года, зять короля Эрика Эрикссона, регент с 1250 до смерти. Основатель Стокгольма. У Биргера и его сына Эрика определена Y-гаплогруппа I1[22].
- Уоррен Баффетт — американский предприниматель, крупнейший в мире и один из наиболее известных инвесторов, состояние которого на сентябрь 2018 года оценивается в 108,4 млрд долларов. В исследованиях компании 23andMe, специализирующейся на ДНК тестах, упоминается, что Баффет генетически близок (по линии отличной от mtDNA) к людям, живущим в северной и центральной Европе, и его предки жили там с древних времён[23]. Это характеристика совпадает с историей и картой распространения Y-гаплогруппы I1. Крупные интернет-порталы, специализирующиеся на истории Европы и генетической генеалогии, интерпретируют эту информацию как наличии у Уоррена Баффетта Y-гаплогруппы I1[].
- Джимми Баффетт — американский музыкант. Упоминается в том же исследовании компании 23andMe[23].
- Пётр Толстой — российский политик, журналист, продюсер и телеведущий. Заместитель председателя Государственной думы Федерального собрания Российской Федерации VII созыва с 5 октября 2016 года и руководитель российской делегации в Парламентской Ассамблее ОБСЕ с 2017 года. Праправнук графа Л. Н. Толстого. В рамках проекта «Моя родословная» его Y-гаплогруппа была определена как I1[24].
- Лев Николаевич Толстой — один из наиболее известных русских писателей и мыслителей, один из величайших писателей-романистов мира. Вероятнее всего имел Y-гаплогруппу I1, так как является предком Петра Толстого[24] по прямой мужской линии (см. выше).
- Стинг — британский музыкант-мультиинструменталист, певец и автор песен, актёр, общественный деятель и филантроп. Лидер группы The Police в 1976—1984 годах. С 1984 года выступает сольно. В восьмом эпизоде второго сезона шоу Finding Your Roots было показано, что он принадлежит к Y-гаплогруппе I1[25].
- Егор Летов — советский и российский поэт, музыкант, певец, звукорежиссёр, художник-оформитель и коллажист. Основатель, лидер и единственный постоянный участник группы «Гражданская оборона», также известен по музыкальным проектам «Егор и Опизденевшие», «Коммунизм» и др. [26]
- Ипполит Антонович Монигетти — русский архитектор и акварелист, представитель архитектурной эклектики, много работавший по заказам царской фамилии и высшей аристократии. Академик и профессор Императорской Академии художеств. Действительный статский советник (1874)[27].
- У Людвига ван Бетховена определили Y-хромосомную гаплогруппу I1a-Z139[28].
Примечания
- ↑ YTree v5.02 at 11 February 2017. Дата обращения: 13 февраля 2017. Архивировано 7 ноября 2017 года.
- ↑ Chiara Batini et al. Large-scale recent expansion of European patrilineages shown by population resequencing Архивная копия от 25 ноября 2017 на Wayback Machine, 2015
- ↑ I-L121 Архивная копия от 23 марта 2023 на Wayback Machine // YFull. YTree v11.01.00
- ↑ Annina Preussner, Jaakko Leinonen, Juha Riikonen, Matti Pirinen, Taru Tukiainen. Y chromosome sequencing data suggests dual paths of haplogroup N1a1 into Finland, February 26, 2024
- ↑ Vanessa Villalba-Mouco et al. Survival of Late Pleistocene Hunter-Gatherer Ancestry in the Iberian Peninsula Архивная копия от 17 августа 2019 на Wayback Machine, March 14, 2019
- ↑ Torsten Günther et al. Population genomics of Mesolithic Scandinavia: Investigating early postglacial migration routes and high-latitude adaptation Архивная копия от 15 ноября 2020 на Wayback Machine, 2018
- ↑ Szécsényi-Nagy et al. (2015), Tracing the genetic origin of Europe’s first farmers reveals insights into their social organization, Proceedings of the Royal Society B, vol. 282, no. 1805, 20150339. (Previously published online 2014 elsewhere before print). Архивная копия от 1 апреля 2016 на Wayback Machine
- ↑ Szécsényi-Nagy (2015), Molecular genetic investigation of the Neolithic population history in the western Carpathian Basin, PhD thesis Johannes Gutenberg-Universität in Mainz. Архивная копия от 21 июля 2015 на Wayback Machine
- ↑ Allentoft M. et al. (2015), Population genomics of Bronze Age Eurasia, Nature, 522, 167?172 (11 June 2015).
- ↑ 1 2 Hugh McColl et al. Steppe Ancestry in western Eurasia and the spread of the Germanic Languages, Marth 14 2024 (Supplementary Tables S1-S5)
- ↑ 1 2 Zenczak M. et al. (2017), Y-chromosome haplogroup assignment through next generation sequencing of enriched ancient DNA libraries Архивная копия от 31 августа 2017 на Wayback Machine
- ↑ Rui Martiniano et al. (19 января 2016), Genomic signals of migration and continuity in Britain before the Anglo-Saxons, Nature Communications volume 7, Article number: 10326 (2016) Архивная копия от 25 марта 2019 на Wayback Machine
- ↑ Carlos Eduardo G. Amorim et al. (11 сентября 2018), Understanding 6th-century barbarian social organization and migration through paleogenomics, Nature Communications volume 9, Article number: 3547 (2018) Архивная копия от 7 ноября 2018 на Wayback Machine
- ↑ 1 2 3 Ashot Margaryan et al. Population genomics of the Viking world Архивная копия от 26 марта 2021 на Wayback Machine, 2020 (bioRxiv Архивная копия от 12 февраля 2020 на Wayback Machine)
- ↑ S. Sunna Ebenesersdóttir et al. (1 июня 2018), Ancient genomes from Iceland reveal the making of a human population Архивная копия от 8 ноября 2018 на Wayback Machine, Science 01 Jun 2018: Vol. 360, Issue 6392, pp. 1028—1032
- ↑ I-Y4870 YTree. Дата обращения: 18 апреля 2021. Архивировано 18 апреля 2021 года.
- ↑ Cody Parker et al. A systematic investigation of human DNA preservation in medieval skeletons Архивная копия от 22 декабря 2021 на Wayback Machine, 26 October 2020 (bioRxiv Архивная копия от 22 апреля 2021 на Wayback Machine, May 22, 2020)
- ↑ Kabaev D. A., Chernyaeva L. L., Chernov S. Z., Goncharova N. N., Semenov A. S. Archaeological DNA Data from the XII-XIV Centuries from Ancient Klyazma Settlements Архивная копия от 1 октября 2022 на Wayback Machine // Историческая информатика. – 2022. – № 3. – С. 1 - 9.
- ↑ ISOGG 2017 Y-DNA Haplogroup I. isogg.org. Дата обращения: 18 сентября 2017. Архивировано 30 августа 2017 года.
- ↑ Underhill et al, New Phylogenetic Relationships for Y-chromosome Haplogroup I: Reappraising its Phylogeography and Prehistory in Rethinking the Human Evolution, Mellars P, Boyle K, Bar-Yosef O, Stringer C, Eds. McDonald Institute for Archaeological Research, Cambridge, UK, pp. 33-42, 2007b.
- ↑ Трофимова Н. В. ИЗМЕНЧИВОСТЬ МИТОХОНДРИАЛЬНОЙ ДНК И Y-ХРОМОСОМЫ В ПОПУЛЯЦИЯХ ВОЛГО-УРАЛЬСКОГО РЕГИОНА, 2015
- ↑ Malmström H. et al. Finding the founder of Stockholm: a kinship study based on Y-chromosomal, autosomal and mitochondrial DNA, Annals of Anatomy — Anatomischer Anzeiger, online 5 April 2011 ahead of print.
- ↑ 1 2 Are Warren and Jimmy Buffett Related? Архивная копия от 3 ноября 2018 на Wayback Machine23andMe
- ↑ 1 2 Петр Толстой. Моя родословная. Выпуск от 18.04.2010 Архивная копия от 3 ноября 2018 на Wayback Machine Первый канал
- ↑ The British Invasion Архивная копия от 3 ноября 2018 на Wayback Machine Finding Your Roots
- ↑ Проект "ДНК замечательных людей". Дата обращения: 13 октября 2021. Архивировано 27 октября 2021 года.
- ↑ [ FTDNA I1-Z140 YDNA Project]
- ↑ Tristan James Alexander Begg et al. Genomic analyses of hair from Ludwig van Beethoven Архивная копия от 30 марта 2023 на Wayback Machine, Received 29 May 2022, Revised 11 October 2022, Accepted 13 February 2023, Available online 22 March 2023. Published: March 22, 2023
Ссылки
Дерево гаплогрупп Y-ДНК человека
| Эволюционное древо гаплогрупп Y-хромосомы человека | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Y-хромосомный Адам | |||||||||||||||||||||||||||||||||
| A00 | A0T | ||||||||||||||||||||||||||||||||
| A0 | A1 | ||||||||||||||||||||||||||||||||
| A1a | A1b | ||||||||||||||||||||||||||||||||
| A1b1 | BT | ||||||||||||||||||||||||||||||||
| B | CT | ||||||||||||||||||||||||||||||||
| DE | CF | ||||||||||||||||||||||||||||||||
| D | E | C | F | ||||||||||||||||||||||||||||||
| F* | GHIJK | ||||||||||||||||||||||||||||||||
| G | HIJK | ||||||||||||||||||||||||||||||||
| H | IJK | ||||||||||||||||||||||||||||||||
| IJ | K | ||||||||||||||||||||||||||||||||
| I | J | LT (K1) | K2 | ||||||||||||||||||||||||||||||
| L (K1a) | T (K1b) | K2a/K2a1/NO/NO1 | K2b | ||||||||||||||||||||||||||||||
| N | O | K2b1 | P (K2b2)/P1 | ||||||||||||||||||||||||||||||
| S (K2b1a) | M (K2b1b) | Q | R | ||||||||||||||||||||||||||||||
| Ключевые понятия |  | |
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||
Формирование славянских народов — генофонд, археология | |||||||
|---|---|---|---|---|---|---|---|
| Основное | |||||||
| Генофонд славян |
| ||||||
| Древнеславянские археологические культуры | |||||||

















