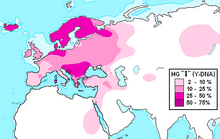
Гаплогруппа I (Y-ДНК)
| Гаплогруппа I | |
|---|---|
 Распространение гаплогруппы I в Европе (в северной зоне преобладает I1, в южной — I2) Распространение гаплогруппы I в Европе (в северной зоне преобладает I1, в южной — I2) | |
| Тип | Y-ДНК |
| Время появления | ~ 25-20.000 до н.э. |
| Место появления | Европа или Передняя Азия |
| Предковая группа | IJ |
| Субклады | I1, I2 |
| Мутации-маркеры | L41/PF3787, M170/PF3715, M258, P19_1, P19_2, P19_3, P19_4, P19_5, P38, P212, U179 |
Гаплогруппа I — гаплогруппа Y-хромосомы человека характеризуемая мутациями L41/PF3787, M170/PF3715, M258, P19_1, P19_2, P19_3, P19_4, P19_5, P38, P212 и U179. Предполагалось, что гаплогруппа I возникла в Европе в эпоху последнего наступления ледников, 25—20 тыс. лет назад, из гипотетической макрогруппы IJ. Однако, гаплогруппа I происходит от мутации гаплогруппы IJ, произошедшей у мужчины, жившего ок. 42 900 лет назад. Последний общий предок современных носителей гаплогруппы I жил 27 500 лет назад (даты определены по снипам компанией YFull[1]).
Её носители могут быть названы прямыми потомками европейского палеолита (кроманьонцев).
Предполагается, что I составляла большинство населения Европы в период до индоевропейского вторжения (около 5 тыс. лет назад). В настоящее время она всё ещё является одной из самых распространённых европейских гаплогрупп, носители различных сохранившихся её ветвей в совокупности составляют до 20 % европейцев (от 10 до 45 % в большинстве европейских стран). Исследование Клио дер Саркиссян о генетике Италии показало, что в распределении гаплогруппы I есть значительная лакуна в Центральной Европе при наличии двух самостоятельных ареалов — скандинавского (I1) и балканского (I2)[2]. Кроме того, носители группы были зафиксированы на Ближнем Востоке, Кавказе и в Центральной Азии. После европейской колонизации мира представители гаплогруппы I живут также и на других континентах.
Происхождение
Согласно современным представлениям, люди современного типа заселили Европу в пределах временного промежутка между 45 и 30 тыс. лет назад, то есть во время последнего межледниковья, когда временное потепление расширило степную зону, в которой охотился ближневосточный Homo sapiens sapiens, до Анатолии[3].
Расселение кроманьонцев по Европе совпало с началом очередного похолодания («последний ледниковый максимум»), которое оказалось одним из самых сильных за несколько последних сотен тысяч лет. Природные условия значительно ухудшились, ледники увеличились, пригодные для жизни территории уменьшились по всему миру, популяция людей сократилась и оказалась в основном ограничена так называемыми ледниковыми убежищами[англ.]. В их числе в Европе граветтская культура. Таким образом, генетическое разнообразие в это время было в очередной раз сокращено, и, вероятно, единственной европейской гаплогруппой, миновавшей образовавшуюся «популяционную горловину» и сохранившейся до наших дней, стала одна из ветвей группы IJ, оформившаяся 25—20 тыс. лет назад, которая и получила имя гаплогруппы I. Разумеется, снова нельзя исключать, что существовали ещё какие-то ветви, пережившие оледенение в Европе, но исчезнувшие в течение следующих тысячелетий[4].
Уже 20—15 тыс. лет назад в группе I дифференцировалась ветвь pre-I1, около 17 тыс. лет назад — ветвь I2, то есть ещё до окончания ледникового периода — возможно, в различных убежищах. Группа pre-I1 могла появиться где угодно от Иберии до Причерноморья, но пережила одну или, скорее всего, несколько «популяционных горловин», так что всё сегодняшнее генетическое разнообразие внутри группы I1 ограничивается пятью-семью последними тысячами лет и исходит из мезолита Северо-Западной Европы. Группа I2 возникла, возможно, на Балканах (или даже в Анатолии, где в 2011 году были обнаружены древнейшие побочные ответвления этой клады) и распространилась по Южной, Центральной и Восточной Европе.
Когда 14—12 тыс. лет назад началось стремительное аллерёдское потепление, ледники отступают на север, меняется расположение климатических зон, люди из «ледниковых убежищ» расселяются на ставшие доступными территории, распространяя по ним гаплогруппу I. Эту эпоху кардинальных перемен в образе жизни, приспособления к новым природным условиям называют эпипалеолитом, переходом к мезолиту. Таким образом, предполагается, что создатели европейских мезолитических культур преимущественно были носителями гаплогруппы I. К культурам, ассоциируемым с группами пред-I1 и I1, относят культуры Эртебёлле, Конгемозе, воронковидных кубков, ямочной керамики, с приходом индоевропейцев бронзового века из культуры боевых топоров они пережили серьёзное сокращение численности и в качестве гипотетического «догерманского субстрата» вошли в основу прагерманского этноса. Распространение группы I2 связывают с последовательностью культур балканского неолита, культурами импрессо, колоколовидных кубков, линейно-ленточной керамики, трипольской.
Также считается, что эту гаплогруппу несли создатели древних европейских мегалитов (5000 — 1200 гг. до н. э.)[], хотя к этому времени в Западной Европе появляется уже и заметная доля неолитических поселенцев с Ближнего Востока, несущих гаплогруппы G2a и E1b1b. Ранее, когда считалось, что группа R1b имеет европейское происхождение, исследователи обращали внимание в частности на то, что область распространения мегалитов в Западной Европе коррелирует с областью современного наибольшего распространения R1b, однако в свете последних исследований это считается совпадением, R1b — группой, пришедшей, вероятно, с индоевропейцами и бронзой, или, по крайней мере, с неолитом, а западноевропейские мегалиты — частью культурной традиции, уходящей в европейский мезолит (хотя конкретный маршрут миграции R1b в западную Европу остаётся ещё предметом споров).
Y-хромосомная гаплогруппа I определена у образца Paglicci 133 (34—31 тыс. л. н.) из пещеры Пальиччи (граветт Италии), образцов Hohle Fels 49 из пещеры Холе-Фельс, Burkhardtshöhle (ок. 15 тыс. л. н.) из Германии (мадленская культура), образца Rochedane (ок. 13 тыс. л. н.) из Франции (эпипалеолит), образцов Chudardes 1, Derry Au Bac (7—8 тыс. л. н.) из Франции (мезолит)[5].
Подгруппы
Поскольку наука о популяционной генетике человека пребывает ещё в процессе становления, родословное древо и номенклатура гаплогрупп подвергается периодическим пересмотрам и уточнениям. В связи с этим следует обращать особое внимание на год публикаций и, соответственно, используемые в них обозначения подгрупп, которые следует проверять по определяющим их мутациям-маркерам. Это особенно актуально для гаплогруппы I, так как её субклады подвергались пересмотру особенно часто (в этом отношении она может сравниться только с другой распространённой в Европе гаплогруппой — R1b).
В гаплогруппе I выделяют две древнейшие подгруппы: I1 (преобладающая в северо-западной Европе) и I2 (преобладающая в южной и восточной Европе).
Гаплогруппа I1
Ветвь I1 (основные мутации M253, M307, P30 и P40) отделена от других пятнадцатью уникальными маркерами, что указывает на её обособление уже 20—15 тысяч лет назад, однако либо она до момента около 5 тысяч лет назад была распространена в совершенно изолированной небольшой популяции (что маловероятно), либо примерно в то же время эта группа пережила серьёзную «горловину», скорее всего, приведшую к исчезновению всех её подветвей, кроме одной, локализованной в северной Европе. Последний общий предок всех зафиксированных на настоящий момент носителей I1 жил 10—7 тысяч лет назад, вероятно, на юго-западном побережье Анцилового озера или на Доггерланде.
Иногда для наименования I1 как древней гаплогруппы (возраста 20-15 тысяч лет) используют термин гаплогруппа пред-I1 (англ. pre-I1). В этом случае ей противопоставляется гаплогруппа I1 как общность всех ныне живущих её потомков, которая в таком случае предстаёт единственной существующей ветвью пред-I1 (возраста 10-5 тысяч лет). В связи с особенностями группы пред-I1 определить район её возникновения не представляется возможным, можно лишь приблизительно обозначить его как Европу, в то время как происхождение «новой» группы I1 можно достаточно уверенно связывать с северо-западом Европы.
Ветвь I1a1-M227 сосредоточена в Восточной Европе и на Балканах и возникла 3700 лет назад[6]. Она зафиксирована в Германии, Чехии, Польше, Эстонии, Украине, Швейцарии, Словении, Боснии, Македонии, Хорватии и в Ливане.
Гаплогруппа I1 была обнаружена у представителя эпипалеолитической азильской культуры BAL0051 из Balma Guianya в Пиренеях (испанская провинция Льейда)[7], у мезолитического обитателя острова Стура-Карлсё (Лен Готланд, Швеция)[8], у представителя культуры линейно-ленточной керамики (неолит Венгрии), жившего ок. 7000 лет назад[9][10] и у представителя бронзового века Швеции (Angmollan, 1493—1302 года до н. э.)[11].
Гаплогруппа I2
Гаплогруппа I2 представляет, по современным предположениям, прямых потомков палеолитических народов Западной, Центральной и Юго-Восточной Европы: от северной Испании до Карпат и от Британских островов до Балкан. Она могла возникнуть около 17 тыс. лет назад в Центральной или Юго-Восточной Европе или даже в Анатолии, поскольку представители корневой парагруппы I2* зафиксированы пока только в Турции, Армении и Грузии.
Шесть крупнейших ветвей включают в себя: I2a1a (M26), I2a1b (M423), I2a2a (M223), I2a2b (L38), I2b (L416) и I2c (L596).
Субклад I2a обнаружен у представителя азильской культуры из Бишонского грота в Швейцарии, жившего 13 560—13 770 тыс. лет назад[12], у представителя неолитической культуры Лендьель, жившего примерно 4360—4490 лет назад, и у представителя культуры Кёрёш (Старчево-Кришская культура), жившего примерно 5650—5780 лет назад[13]. Гаплогруппы I2 и I2a были обнаружены у представителей энеолитической культуры Ремеделло[14], гаплогруппа I2a2a — у обитателя Атапуэрки (Испания), жившего в медном веке[15], гаплогруппа I2a1 — у представителя культуры ямочной керамики Ajvide 58 (2800—2000 BC)[16], гаплогруппа I2a1 — у «Человека из Лошбура» (5998 лет до нашей эры) с мезолитической стоянки Хеффинген-Лошбур близ Хеффингена (Люксембург)[17]. Гаплогруппа I2b2 была обнаружена в скелетных останках из пещеры Лихтенштейн (бронзовый век, Германия), где также были найдены артефакты культуры полей погребальных урн[18]. Гаплогруппы I2a1 и I2c2 известны из раскопок в шведской Мутале (мезолит)[19]. У представителей мегалитической культуры были обнаружены гаплогруппы I2a2a1 (Ла Мина, Испания)[19] и I2a1 (дольмен La Pierre Fritte, Франция)[20]. I2a2 (xI2a2a, I2a2b) или I2a1b*-Y10705>I2a1b2a-S2524>S2524*[21] (ISOGG 2018) определена у темнокожего и голубоглазого человека из Чеддара[22].
Субклады I2a2a1-CTS9183, I2a2a1a2a1a-S391, I2a2a1b2a2-S390, I2a2a1b2a2a2-ZS20 найдены у варваров VI века с кладбища лангобардов в Соладе (Szólád, Венгрия)[23].
I2a1
I2a1a (M26, L158, L159.1/S169.1) до 2005 г. обозначалась I1b2, I1b1b в 2006—2007 гг., и I2a1 с 2008 по 2010 гг. Главным образом, обнаружена среди сардинцев (39 %[24]) и басков, а также встречается на Иберийском полуострове, в западной Франции, на западном побережье Италии и средиземноморском побережье Магриба. Она составляет до 40 % всех Y-хромосомных гаплогрупп на Сардинии. Возраст I2a1a оценивается в 8 тыс. лет.
I2a1b (M423, L178) до 2007 года обозначалась как I1b, с 2008 по 2010 год обозначалась I2a2. Типична для населения Балкан и Карпат, чаще всего наблюдается у динарских славян (сербы, хорваты и боснийцы), а также в Молдове, Беларуси и на юго-западной Украине. Также с заметной частотой обнаруживается в Албании, северной Греции, Болгарии, Словакии, на восточной Украине, в Беларуси и юго-западной России. Эта группа редка, но встречается на Британских островах. Так называемый «динарский» (Dinaric) субклад I2a1b2a1a-CTS10228 выделяется своей численностью. Его современный пик на Динарском нагорье является следствием недавних демографических событий во время Великого Переселения Народов, второй пик в Полесье маркирует место, где начался рост данного субклада. При анализе ископаемой ДНК на Балканах не обнаружено ветвей гаплогруппы I2, к которым мог бы восходить субклад I2a1b2a1a-CTS10228. Единственными древними находками этого субклада в Восточной Европе являются образцы домонгольского времени — Сунгирь-6, VK542 (предполагаемые останки Глеба Святославича, внука Ярослава Мудрого) и образец I18719 из пещеры Безданяча (Bezdanjača Cave) в Хорватии, который археологически датируется поздним бронзовым веком (3200 л. н., Croatia_MBA_LBA)[25][26].
Современный разброс I2a1a и I2a1b (Иллирия, Италия, Сардиния, средиземноморское побережье Франции и Испании) совпадает с распространением неолитической культуры кардиальной керамики (5000 — 1500 гг. до н. э.), появление которой связывают с приходом земледельцев и скотоводов из района Фессалии (гаплогруппы E-V13 и G2a). За нею следовала культура бронзового века Террамаре (1500—1000 гг. до н. э.). Предполагается, что итало-кельтские носители Y-хромосомной гаплогрупы R1b перешли Альпы и заселили Италию около 1000 года до н. э., потеснив большинство местного населения с гаплогрупами I2a1, G2a и E-V13 (особенно в северной части).
I2a2
I2a2 (S33/M436/P214, P216/S30, P217/S23, P218/S32, L35/S150, L37/S153, L181) или I2a1b в ISOGG 2019-2020 (до 2005 г. обозначалась I1c, а с 2005 по 2010 гг. I2b). Связана с докельто-германским населением Северо-Западной Европы, таким как строители мегалитов (5000 — 1200 гг. до н. э.). Широкое разнообразие КТП-меток внутри I2a2a может говорить о её возрасте в 13 тыс. лет.
I2a2 (I2a1b) находят по всей Западной Европе, но очевидно, что она лучше всего пережила индоевропейское вторжение в Северной Германии и оттуда была повторно разнесена с германскими миграциями. В настоящее время чаще всего I2a2 (I2a1b) встречается в центральной и северной Германии (10—20 %), Бенилюксе (10—15 %) и в северной Швеции. Также зафиксирована с частотами от 3 до 10 % в Дании, восточной Англии и северной Франции. Она редка в Норвегии, что согласуется с тем фактом, что миграции из северной Германии не были направлены в эту сторону.`
Выделены две основные подгруппы: I2a2a (I2a1b1-M223 в ISOGG 2019-2020) и I2a2b (I2a1b2a-L38/S154 в ISOGG 2019-2020), каждая из них включает, по меньшей мере, ещё по четыре подгруппы, однако пока они исследованы очень мало. Ветвь I2a2a1 (I2a1b1a1a-M284 в ISOGG 2019-2020) имеет место практически исключительно в Британии, где она, вероятно, и возникла около 3 тыс. лет назад.
I2b
I2b (L416, L417, L418) чрезвычайно немногочисленная, реликтовая ветвь, пока обнаруженная лишь в нескольких образцах из Германии, Италии и Шотландии.
I2c
I2c (I2a2-L596/L597 в ISOGG 2019-2020), вероятнее всего, возникла в районе Рейна. В ней выделяют три подгруппы (кластера): A, B и C.
Кластер I2c-A географически ограничен Германией, Швейцарией, Британией и Ирландией. Предполагается, что он распространился с кельтами железного века. Кластер I2c-C распространён на той же территории, что I2c-A, но также ещё во Франции, Италии и Норвегии. Предполагается, что он распространялся в основном с кельтами и ранними германцами бронзового века. Распространение I2c-A и I2c-C коррелирует с R1b-S116/P312.
Кластер I2c-B с низкой частотой зафиксирован в восточной Европе и Закавказье, а именно: в Польше, Литве, России, на Украине, Молдове, Румынии, Болгарии, Турции, Грузии, Армении, Азербайджане, в северо-западной части Южного Азербайджана и на Крите. Вероятное место происхождения — Балканы.
Примечания
- ↑ I YTree. Дата обращения: 24 февраля 2017. Архивировано 25 февраля 2017 года.
- ↑ Археогенетика Европы: новые исследования Архивная копия от 25 февраля 2017 на Wayback Machine, 17 декабря 2013
- ↑ В ту эпоху география европейского континента заметно отличалась от современной. Белого, Балтийского, Северного и Азовского морей не существовало, Британские острова соединялись с материком Доггерлендом, весь север континента от Коми до района Берлина и до Шотландии был занят ледником, крупные ледники также располагались на Альпах и Пиренеях, а также в других гористых районах. Не существовало Босфора, Чёрное море было бессточным водоёмом, подобно нынешнему Каспийскому, его уровень был значительно ниже, а размеры меньше. Благодаря тому, что и уровень Мирового океана был значительно ниже, многие прибрежные территории являлись сушей, например почти половина Адриатики, многие острова Эгейского и Адриатического морей соединялись с материком, как и Сицилия. Средиземноморское побережье, Пиренеи, Апеннины, Балканы были покрыты редкими лесами, но уже с территории нынешних Франции, Словении, Румынии и на север до самого ледника простиралась тундростепь.
- ↑ Интересно, что по меньшей мере 5 тысячелетий люди современного типа соседствовали в Европе с неандертальцами, которые, хотя и были физически лучше приспособлены к холодному климату и уже не раз переживали периоды наступления ледников, в этот раз вымерли. Представляется, что прямо или косвенно это связано с появлением в Европе кроманьонцев. Уже 30 тыс. лет назад неандертальцы исчезли с большей части своего ареала, возраст самых поздних подтверждённых неандертальских останков в пещере в Гибралтаре оцениваются в 24 тысячи лет. Характер взаимоотношений двух подвидов гоминид, возможность их культурного и генетического взаимовлияния активно изучается и служит предметом научных споров и околонаучных спекуляций. В 2010 году появилось исследование, доказывающее факт, по крайней мере, одной гибридизации Homo sapiens sapiens и Homo sapiens neanderthalensis на Ближнем Востоке около 60 тыс. лет назад, а также ещё одной гибридизации неоантропа с представителем другой популяции Homo erectus в Азии около 40 тыс. лет назад. Таким образом, возможность гибридизации людей современного вида с европейскими неандертальцами так же существует, тем более что многократно обращалось внимание на фенотипическую близость неандертальцев и европейцев, однако на настоящее время факт такого обмена генами не доказан. В этом, гипотетическом пока, случае среди представителей гаплогруппы I носителей следов неандертальской наследственности найти будет особенно вероятно.
- ↑ Qiaomei Fu et al. The genetic history of Ice Age Europe, 2016.
- ↑ [1] Архивная копия от 23 марта 2023 на Wayback Machine // YFull. YTree
- ↑ Vanessa Villalba-Mouco et al. Survival of Late Pleistocene Hunter-Gatherer Ancestry in the Iberian Peninsula Архивная копия от 17 августа 2019 на Wayback Machine, March 14, 2019
- ↑ Torsten Günther et al. Population genomics of Mesolithic Scandinavia: Investigating early postglacial migration routes and high-latitude adaptation Архивная копия от 15 ноября 2020 на Wayback Machine, 2018
- ↑ Szécsényi-Nagy et al. (2015), Tracing the genetic origin of Europe’s first farmers reveals insights into their social organization, Proceedings of the Royal Society B, vol. 282, no. 1805, 20150339. (Previously published online 2014 elsewhere before print). Архивная копия от 1 апреля 2016 на Wayback Machine
- ↑ Szécsényi-Nagy (2015), Molecular genetic investigation of the Neolithic population history in the western Carpathian Basin, PhD thesis Johannes Gutenberg-Universität in Mainz. Архивная копия от 21 июля 2015 на Wayback Machine
- ↑ Allentoft M. et al. (2015), Population genomics of Bronze Age Eurasia, Nature, 522, 167?172 (11 June 2015).
- ↑ Jones, E. R. et al. Upper Palaeolithic genomes reveal deep roots of modern Eurasians, 2015. Дата обращения: 18 ноября 2015. Архивировано 21 августа 2016 года.
- ↑ Genome flux and stasis in a five millennium transect of European prehistory. Дата обращения: 1 мая 2015. Архивировано 29 апреля 2015 года.
- ↑ Morten E. Allentoft et al. «Population genomics of Bronze Age Eurasia». Дата обращения: 11 сентября 2015. Архивировано 30 апреля 2016 года.
- ↑ Ancient genomes link early farmers from Atapuerca in Spain to modern-day Basques. Дата обращения: 11 сентября 2015. Архивировано 20 октября 2019 года.
- ↑ Skoglund, P. et al. (2014), Genomic diversity and admixture differs for Stone-Age Scandinavian foragers and farmers, Science
- ↑ «Ancient human genomes suggest three ancestral populations for present-day Europeans». Дата обращения: 11 сентября 2015. Архивировано 16 марта 2021 года.
- ↑ Lichtenstein Cave Data Analysis Архивная копия от 6 марта 2009 на Wayback Machine | Ken Nordtvedt: «The Lichtenstein cave ydna haplotypes show three from the new S23+(xM223) I2b* (ISOGG 2008) tree» Архивная копия от 5 мая 2009 на Wayback Machine
- ↑ 1 2 Haak, W. et al. (2015), Massive migration from the steppe is a source for Indo-European languages in Europe
- ↑ Lacan, M. et al. (2011), Ancient DNA suggests the leading role played by men in the Neolithic dissemination
- ↑ I-S2524 YTree. Дата обращения: 1 января 2022. Архивировано 1 января 2022 года.
- ↑ Selina Brace et al. Ancient genomes indicate population replacement in Early Neolithic Britain Архивная копия от 17 апреля 2019 на Wayback Machine, 2019
- ↑ Carlos Eduardo G. Amorim, Krishna R. Veeramah et al. Understanding 6th-century barbarian social organization and migration through paleogenomics Архивная копия от 7 ноября 2018 на Wayback Machine, 2018
- ↑ Charleston W. K. Chiang et al. Genomic history of the Sardinian population
- ↑ I-CTS10228 YTree. Дата обращения: 27 декабря 2021. Архивировано 27 декабря 2021 года.
- ↑ Nick Patterson et al. Large-scale migration into Britain during the Middle to Late Bronze Age Архивная копия от 1 января 2022 на Wayback Machine // Nature, 22 December 2021
Ссылки
- ISOGG 2016 Y-DNA Haplogroup I
- Русскоязычный ДНК форум «Родство»
- Русскоязычный ДНК форум «Молген»
- Карта гаплогрупп y-хромосомы
| Эволюционное древо гаплогрупп Y-хромосомы человека | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Y-хромосомный Адам | |||||||||||||||||||||||||||||||||
| A00 | A0T | ||||||||||||||||||||||||||||||||
| A0 | A1 | ||||||||||||||||||||||||||||||||
| A1a | A1b | ||||||||||||||||||||||||||||||||
| A1b1 | BT | ||||||||||||||||||||||||||||||||
| B | CT | ||||||||||||||||||||||||||||||||
| DE | CF | ||||||||||||||||||||||||||||||||
| D | E | C | F | ||||||||||||||||||||||||||||||
| F* | GHIJK | ||||||||||||||||||||||||||||||||
| G | HIJK | ||||||||||||||||||||||||||||||||
| H | IJK | ||||||||||||||||||||||||||||||||
| IJ | K | ||||||||||||||||||||||||||||||||
| I | J | LT (K1) | K2 | ||||||||||||||||||||||||||||||
| L (K1a) | T (K1b) | K2a/K2a1/NO/NO1 | K2b | ||||||||||||||||||||||||||||||
| N | O | K2b1 | P (K2b2)/P1 | ||||||||||||||||||||||||||||||
| S (K2b1a) | M (K2b1b) | Q | R | ||||||||||||||||||||||||||||||
| Ключевые понятия |  | |
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||