Гаплогруппа P (Y-ДНК)
| Гаплогруппа P | |
|---|---|
| Тип | Y-ДНК |
| Время появления | ~32 тыс. л. н.[1] (41–27 тыс.л.н.[2]) |
| Место появления | Центральная Азия, Сибирь или Китай |
| Предковая группа | K2b |
| Сестринские группы | K2b1 (далее M и S) |
| Субклады | P1 (далее Q и R), P2 |
| Мутации-маркеры | 92R7_1, 92R7_2, L138, L268, L405, L471/PF5989, L536/PF5860, L721/PF6020, L741, L768/PF5976/YSC0000274, L779/PF5907/YSC0000251, L781/PF5875/YSC0000255, M45/PF5962, M74/N12, P27.1_1/P207, P27.1_2, P69, P226/PF5879, P228/PF5927, P230/PF5925, P235/PF5946, P237/PF5873, P239, P240/PF5897, P243/PF5874, P244, P281/PF5941, P282/PF5932, P283/PF5966, P284, P295/PF5866/S8, Page83, V231 |
В популяционной генетике и геногеографии человека, изучающих Y-хромосомные гаплогруппы, гаплогруппа P-P295 (она же K2b2) — патрилинейная наследственность, возникшая, вероятнее всего, более 30 тысяч лет назад в Азии, определяемая по целому ряду уникальных меток (а именно: 92R7_1, 92R7_2, L138, L268, L405, L471/PF5989, L536/PF5860, L721/PF6020, L741, L768/PF5976/YSC0000274, L779/PF5907/YSC0000251, L781/PF5875/YSC0000255, M45/PF5962, M74/N12, P27.1_1/P207, P27.1_2, P69, P226/PF5879, P228/PF5927, P230/PF5925, P235/PF5946, P237/PF5873, P239, P240/PF5897, P243/PF5874, P244, P281/PF5941, P282/PF5932, P283/PF5966, P284, P295/PF5866/S8, Page83, V231), что может говорить о периоде достаточно долгой её изоляции. Поскольку от неё через P1-M45=K2b2a происходят гаплогруппы R и Q, является макрогруппой.
Потомки гаплогруппы P=K2b2 доминируют среди индоевропейцев (подветвь R) и тюркских народов (подветвь R), и американских индейцев (подветвь Q), а также составляют значительную часть населения Центральной и Южной Азии.
Происхождение
Гаплогруппа P (она же K2b2) произошла от субклады K2b-M1221/P331/PF5911[3] примерно 44 300 л. н. Последний общий предок современных носителей гаплогруппы P жил 31 900 лет назад (даты определены по снипам компанией YFull[4]). Субклада K2b произошла от гаплогруппы K2=K(xLT) (M526), которая в свою очередь происходит от гаплогруппы K (M9). От гаплогруппы P=K2b2 произошла субклада P1=K2b2a (M45).
Древним носителем предковой K2b* был Тяньюаньский человек[англ.], живший ~40 тыс.л.н. недалеко от Пекина. А вот пришли ли его предки северным континентальным путём (через Алтай) или южным береговым (через Индокитай и Китай), и как именно расселялись потомки – на этот счёт в ходу две гипотезы:
- Гаплогруппа P возникает к северу от Гиндукуша, в Сибири, Казахстане, Узбекистане, либо же где-то на протяжении Великого шёлкового пути на территории китайских провинций Синьцзяна, Ганьсу или Нинся́, затем около 35 тыс.л.н. она широко распространяется по тундростепи в северных направлениях. Эта версия поддерживается тем, что P* (P1*) встречается в Средней, Восточной и Северо-Восточной Азии (а также среди американских индейцев), а также что к этому району примыкают и зоны распространения дочерних от P1 групп: — R-M207 (K2b2a2) и Q-M207 (K2b2a1). Древнейший носитель подгруппы P1 (32-30 тыс.л.н.) также обнаружен в Сибири – на Янской стоянке почти у са́мого Ледовитого Океана.
- Движение предковой группы K2b шло южным маршрутом — вдоль Сычуаньских Альп и — уже из района Ганьсу и Нинся́ — вдоль Шёлкового пути на запад и в сторону Чукотки на север-восток. Климат в ту эпоху отличался от современного, так что в Таримской низменности, Монголии и Маньчжурии была более обильная флора и фауна, что могло сделать те края более привлекательными для жизни. Уровень моря был до 112 метров ниже 18 тыс.л.н., и почти столь же низким на протяжении последних 100 тысячелетий, что способствовало быстрому прибрежному расселению гаплогруппы K и K(xLT) по Восточной Азии, через степи севернее Пекина на запад к Таримской впадине и на восток в Маньчжурию. Эта версия подкрепляется фактами обнаружения древних линий P* в Южной, Юго-Восточной и Восточной Азии (а также в Океании), а также тем, что именно в юго-восточной Азии и Океании распространена и сестринская для P группа K2b1 (её линии M (K2b1b) и S (K2b1a)).
Возможно, что большинство или многие из описанных случаев P* на Востоке в действительности относятся к недавно открытой кладе P2, тогда группа распадается на "сибирский кластер" P1 и "филиппинский кластер" P2. Но в любом случае, присутствие в китайском регионе P* и MS оставляет хорошую поддержку версии о возникновении группы P именно где-то на юго-востоке Азии. С другой стороны, родственная группа NO1 (K2a2), хоть и подверглась наибольшей дивергенции в монгольском регионе, самые древние линии N* и O* также в Сычуани, а её предковая K2a* – у Усть-Ишимского человека и (K2a1) на древних Карпатах и (единичные случаи) в Индии. Так что возможен почти любой сценарий перемещения групп в эпоху 40-30 тыс.л.н. между Восточной Азией и Сибирью: это было время огромной мобильности маленьких популяций.
Палеогенетика
- У двух фрагментированных молочных зубов, найденных на Янской стоянке (возраст ~31,6 тыс. лет назад), определена Y-хромосомная гаплогруппа P1 (K2b2)[5].
- Гаплогруппа P1 была обнаружена у представителя потаповской культуры I0246/SVP41 (могильник Утёвка VI), жившего 2200—1800 лет до н. э.[6], и у обитателя неолитического поселения Ганджи-Даре в Иранском Курдистане[7]. P1 в немецких образцах Esperstedt (I1542/ESP33) и Tiefbrunn (RISE434) возможно недотипированные образцы.
- Гаплогруппу P-P295 определили у образца HistAndaman historical с Андаманских островов[8].
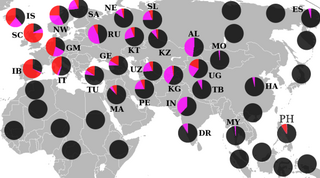
Этногеографическое распределение
Подавляющее большинство носителей M45 относятся к одной из двух её больших ветвей: R или Q, однако парагруппа P* также встречается у сибирских и дальневосточных народов, в Средней и Восточной Азии, а также на хорватском острове Хвар (что могло произойти вследствие миграции аваров), у ашкеназов, индейцев и у народов Южной Азии и Океании. У филиппинского племени аэта базальная гаплогруппа P* достигает 28%, обнаружены субклады P1* (М45) и P2 (Р-B253). У тиморцев Индонезии базальная P* достигает 10%. Остров Лусон является также единственным местом где ветви Р*, Р1* и очень редкая Р2 встречаются вместе[9][10], а также высока доля гаплогруппы K2b1[11].
Примечания
- ↑ Tatiana M. Karafet, Fernando L. Mendez, Monica B. Meilerman, Peter A. Underhill, Stephen L. Zegura, and Michael F. Hammer (2008). New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree
- ↑ Karafet T. M., Mendez F. L., Meilerman M. B., Underhill P. A., Zegura S. L., Hammer M. F. New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree (англ.) // Genome Res. : journal. — 2008. — May (vol. 18, no. 5). — P. 830—838. — doi:10.1101/gr.7172008. — PMID 18385274. — PMC 2336805.
- ↑ Tatiana M. Karafet et all. Improved phylogenetic resolution and rapid diversification of Y-chromosome haplogroup K-M526 in Southeast Asia, 2014
- ↑ P YTree v6.06.17. Дата обращения: 29 октября 2016. Архивировано 30 октября 2016 года.
- ↑ Martin Sikora, Vladimir Pitulko, Qiaomei Fu, Sergey Vasilyev, Elizaveta Veselovskaya, Margarita Gerasimova, Elena Pavlova, Vyacheslav Chasnyk, Pavel Nikolskiy, Pavel Grebenyuk, Alexander Fedorchenko, Alexander Lebedintsev, Boris Malyarchuk et al. The population history of northeastern Siberia since the Pleistocene Архивная копия от 24 октября 2018 на Wayback Machine
- ↑ Iain Mathieson et al. Eight thousand years of natural selection in Europe Архивная копия от 3 марта 2016 на Wayback Machine, 2015
- ↑ Iosif Lazaridis et al. The genetic structure of the world’s first farmers Архивная копия от 16 июля 2018 на Wayback Machine, 2016.
- ↑ J. Víctor Moreno-Mayar et al. Early human dispersals within the Americas Архивная копия от 25 ноября 2018 на Wayback Machine, 2018
- ↑ Tumonggor, Karafet et al., 2014, "Isolation, contact and social behavior shaped genetic diversity in West Timor", Journal of Human Genetics Vol. 59, No. 9 (September), pp. 494–503
- ↑ E. Heyer et al., 2013, "Genetic Diversity of Four Filipino Negrito Populations from Luzon: Comparison of Male and Female Effective Population Sizes and Differential Integration of Immigrants into Aeta and Agta Communities", Human Biology, Vol. 85, Iss. 1, p. 201
- ↑ гаплогруппы K2b1a и М (K2b1d), доминируют у мужчин в Меланезии, а также значительны в смежных частях Океании и Малайского архипелага
| Эволюционное древо гаплогрупп Y-хромосомы человека | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Y-хромосомный Адам | |||||||||||||||||||||||||||||||||
| A00 | A0T | ||||||||||||||||||||||||||||||||
| A0 | A1 | ||||||||||||||||||||||||||||||||
| A1a | A1b | ||||||||||||||||||||||||||||||||
| A1b1 | BT | ||||||||||||||||||||||||||||||||
| B | CT | ||||||||||||||||||||||||||||||||
| DE | CF | ||||||||||||||||||||||||||||||||
| D | E | C | F | ||||||||||||||||||||||||||||||
| F* | GHIJK | ||||||||||||||||||||||||||||||||
| G | HIJK | ||||||||||||||||||||||||||||||||
| H | IJK | ||||||||||||||||||||||||||||||||
| IJ | K | ||||||||||||||||||||||||||||||||
| I | J | LT (K1) | K2 | ||||||||||||||||||||||||||||||
| L (K1a) | T (K1b) | K2a/K2a1/NO/NO1 | K2b | ||||||||||||||||||||||||||||||
| N | O | K2b1 | P (K2b2)/P1 | ||||||||||||||||||||||||||||||
| S (K2b1a) | M (K2b1b) | Q | R | ||||||||||||||||||||||||||||||
| Ключевые понятия |  | |
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||