Гаплогруппа U (мтДНК)
| Гаплогруппа U | |
|---|---|
 | |
| Тип | мтДНК |
| Время появления | 60 тыс. лет назад |
| Место появления | Ближний Восток |
| Предковая группа | Гаплогруппа R |
В популяционной генетике человека гаплогруппой U называют одну из гаплогрупп, выявленных при анализе последовательности мутаций в митохондриальной ДНК (mtDNA). Эта гаплогруппа широко распространена в Европе, её носители отделились от макрогруппы R, подгруппы в гаплогруппе N, около 60 тыс. лет назад. За прошедшее с тех пор время гаплогруппа U, кроме Европы, распространилась также в Северной Африке, Аравии, Индии и на Ближнем Востоке[1].
Разделилась на 8 подгрупп, от гаплогруппы U8 отделилась также гаплогруппа К[2].
Гаплогруппу U определили у образца MA-1 со стоянки Мальта́ (23 891—24 423 л. н.)[3].
Гаплогруппу U, содержащую семь «диагностических» позиций, которые отличают её от последовательностей мтДНК других людей (Дольни-Вестонице-14, Лошбур, Оберкассель, саамы), определили у образца DCP1 из Денисовой пещеры (от 19 000 до 25 000 л. н.)[4].
Гаплогруппа U1
В основном встречается на Ближнем Востоке, хотя с более низкой частотой может быть также обнаружена во всём Средиземноморье. Например, U1a распространена на большом пространстве от Индии до Атлантики, но очень редко встречается на западных и северных берегах Европы, в том числе на Британских островах и в Скандинавии. Несколько носителей обнаружено в Италии (в Тоскане). В Индии U1a найдена в западной части. U1b распространена в тех же регионах, но с ещё меньшей частотой. Немногочисленные носители обнаружены в еврейской диаспоре. В Восточной Европе U1a и U1b распространены с одинаковой частотой[5]. U1a1a определена у образцов из римско-христианского кладбища Келлис 2 (K2) в египетском оазисе Дахла[6]. U1b2 определена у образца из карельской Хийтолы (1200—1500 гг.) в Лахденпохском районе[7].
Гаплогруппа U5
Наряду с U8a, U5 — старейшая гаплогруппа в Европе. Возраст — от 50 до 60 тыс. лет[8]. Чаще всего U5 встречается на севере, среди саамов, финнов и эстонцев (преимущественно U5b), но с низкой частотой может быть обнаружена и в остальных частях Европы, на Ближнем Востоке и в Африке[1][9]. Возраст финской линии U5b1b2 составляет около 6000 лет[10].
U5 определили у палеолитических обитателей Дольни-Вестонице (граветтская культура), живших 31 тыс. лет назад[11].
U5b2b определили у двух поздних эпиграветтских образцов из сицилийской пещеры Сан-Теодоро[итал.] (San Teodoro 3 и San Teodoro 5), живших после последнего ледникового максимума (LGM) 15 322—14 432 лет назад[12].
U5b2b определили у образца Villabruna 1 (ок. 14 тыс. л. н.) из локации Рипари Виллабруна в Северной Италии (коммуна Соврамонте)[13].
U5b1h определили у палеолитического обитателя Бишонского грота в Швейцарии, жившего 13 560—13 770 лет назад[14].
U5b1 определили у представителя мадленской культуры из Оберкасселя (Двойное захоронение в Оберкасселе[нем.]), жившего 13,4 тыс. лет назад[11].
U5a2 определили у образца Sidelkino (11,55 тыс. л. н.) со стоянки Гора Маяк у села Сиделькино Самарской области[15].
U5b2 определили у образца HG1 из погребения 37 (Vasilyevka 3, 11 143—10 591 л. н.) в Васильевке-на-Днепре[16].
U5b1 определили у образца Arene Candide 16 (10 810±65 л. н.[17]) из пещеры Арене Кандиде[англ.] в Италии[18].
U5a2 определили у образца I1737 (9000—7500 лет до н. э.) из Василевки-на-Днепре[19].
U5b2b у определили мезолитического образца AVH-1 (10,211—9,91 тыс. л. н.) из пещеры Арма Вейрана (Arma Veirana) в Лигурийских Альпах (Италия)[20].
U5a2d определили у трёх человек с мезолитической стоянки Гусиный Клюв (Huseby Klev) на западе Швеции (о. Уруст), живших около 9880—9540 лет назад. ДНК выделена из слюны, извлечённой из фрагментов дёгтя[21].

Эта гаплогруппа обнаружена у темнокожего и голубоглазого Чеддарского человека (U5b1) — мумии эпохи мезолита из Англии, а также у ископаемых скелетов охотников-собирателей из Германии, Польши, Литвы и Испании[22].
U5b2 определили у трёх мезолитических образцов из Василевки-на-Днепре: I1819 (8825—8561 лет до н. э.), I1763 (8280—7967 лет до н. э.), I1734 (7446—7058 лет до н. э.)[19].
U5a2 определили у мезолитического образца Lec2 (10,39 тыс. л. н.) из крымской пещеры Лесника (Lesnik Cave), известной также как Медвежья пещера (Bear Cave)[23][24].
U5 определили у темнокожего мезолитического «человека из Лошбура»[англ.] (5998 лет до н. э.) гаплогруппу[25][26].
U5b2c1 определили у темнокожего и голубоглазого мезолитического охотника-собирателя La Braña 1[исп.] из пещеры Ла Брана-Аринтеро, жившего около 7 тыс. лет назад на северо-западе Иберийского полуострова, и у его брата La Braña 2[27][28].
U5a1 определили у неолитического образца I1958 (4723—4558 гг. до н. э.) со стоянки со стоянки Мергень 6 на озере Мергень в Тюменской области[29].
U5b1b1 (U5b1b-a*[30]) определили у неолитического образца I4893 (4446—4347 лет до н. э.) из пражского района Кобылисы[чеш.] (Чехия)[31].
U5a1i определили у представителя хвалынской культуры[32].
U5 определили у представителя культуры кардиальной керамики[33].
U5b определили у трёх представителей неолитической сернийской культуры (север Франции)[34].
U5a2+16362 определили у представителя трипольской культуры (3789—3650 лет до н. э.) из пещеры Вертеба[35].
U5a1 (U5a1i[36]) обнаружена у представителя катакомбной культуры RK4001 (2451—2203 лет до н. э.)[37].
U5 определили у представителей ямной культуры[37].
U5a1a обнаружили у представителей днепро-донецкой культуры[38].
U5a1d1 определили у образцов RV 1852 и 2018/1 со стоянки Риннюкалнс (Riņņukalns) на реке Салаца в Буртниекском крае на севере Латвии, U5a2b2 определили у образца RV 2039 (5300—5050 л. н.). Кроме того, у RV 2039 удалось зафиксировать следы возбудителя чумы грамотрицательной бактерии Yersinia pestis[39].
U5 определили у представителей межовской культуры[37].
U5a1a2a, U5a1b1 и U5a2c определили представителей унетицкой культуры из Чехии (Czech_EBA_Unetice 3850—4100 л. н.)[40].
U5b2b5 обнаружили у 4000-летней головы египетской мумии из гробницы в коптской деревне Дейр эль-Берша[41].
U5b2b5 обнаружили у 4000-летней мумии Kadruka 1 с севера Судана (зуб KDR001.A)[42].
U5a1a2a определили у образца I4773 (1618—1513 гг. до н. э.) из среднего-позднего бронзового века Казахстана (Aktogai_MLBA)[29].
Редкий субклад U5b2c1 обнаружен у образца «Ариш»[фр.] из пунического погребального склепа на холме Бирса недалеко от входа в Национальный музей Карфагена в Тунисе возрастом около 2500 лет назад (конец VI века до н. э.)[43]. Предполагается, что эта линия унаследована из Испании, колонизированной финикийцами[44].
U5a1b1 определили у мальчика из двойного захоронения в селе Чулице в окрестностях Кракова (395—418 гг., Y-хромосомная гаплогруппа N)[45].
В финском Леванлухта[англ.] были определены U5a, U5b и характерная для саамов U5b1b1a, у двух образцов из Холлолы (955—1390 гг.) определена гаплогруппа U5a2a1e[7].
U5a1a определили у одного из викингов с языческого захоронения Галгедил (Galgedil) на датском острове Фюн (700—1100 гг. н. э.)[46].
U5b1g определили у средневекового (X век) жителя Исландии с озера Миватн[47].
U5a1g выявлена у крестоносца из Сидона (Ливан), жившего примерно в XI—XIII веках[48].
U5a1b1, U5a2, U5a2a1c определили у трёх женщин из Рубленого города в Ярославле (массовое захоронение № 76, 1238 год)[49].
U5a2a1 определили у человека, жившего 700 л. н. и найденного на восточном Памире (Ташкурган), что указывает на происхождение от доисторического скотоводческого населения Волго-Уральского региона[50]. U5a2a1 сохранился у некоторых представителей памирских народов[51].
U5a1a1 определили у пятимесячного эмбриона win002_all (Y-хромосомная гаплогруппа R1b-DF17>R1b1a1b1a1a2a1a2~-BY1806[52]), обнаруженного в гробу епископа Лундского Педера Винструпа, похороненного в 1679 году[53].
Подгруппа U5a считается особо восприимчивой к синдрому приобретённого иммунного дефицита[54].
Гаплогруппа U6
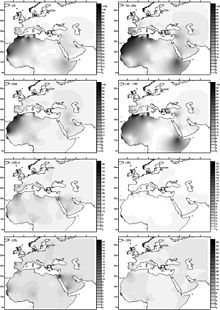
На уровне 10 % распространена в Северной Африке, особенно среди алжирских берберов (29 %)[1][55][56], а также на Пиренейском полуострове, Канарских островах[56] и в Восточной Африке. Хотя на Пиренеях распространение гаплогруппы U6 достигает максимума, считается, что возникла она не здесь, а либо в Леванте 40 тыс. лет назад[57], либо в Восточной Африке между 25 и 66 тыс. лет назад[56].
Примерно 45—40 тыс. л. н. преобладающие ныне североафриканские гаплогруппы M1 и U6 возникли в Юго-Западной Азии и вместе двинулись в Африку[58][59].
U6 начинает расширяться ~ 26 000 л. н.[60]
Среди трёх основных подгрупп U6[56]:
- U6a распространена от Канарских островов и Пиренейского полуострова до Сирии, Эфиопии и Кении с наивысшей частотой в Восточной Африке. Возраст: 24 тыс. л. н. Выделяется также U6a1, распространённая в тех же регионах, но выделившаяся около 15 — 20 тыс. лет назад.
- U6b встречается чаще в западных регионах, в Испании — на севере страны, кроме неё — в Марокко, Алжире, Сенегале и Нигерии. Возраст: 8,5 — 12 тыс. лет. Подгруппа U6b1, выделившаяся около 6 тыс. лет назад, встречается только на Пиренейском полуострове и Канарских островах.
- U6c имеется только в Марокко и на Канарских островах. Возраст: 6 — 17,5 тыс. лет. Не имеет мутации (16219), общей для U6a и U6b.
Базальная гаплогруппа U6* была обнаружена у образца PM1 возрастом 35 тыс. л. н. из румынской «пещеры Женщин» (Мьюрилор[англ.]), что свидетельствует об обратной верхнепалеолитической миграции из Западной Евразии в Северную Африку[61].
U6 определена у обитателя грузинской пещеры Дзудзуана (Dzudzuana Cave), жившего 26 тыс. лет назад[62].
Субклады U6d3, U6a1b, U6a6b, U6a7b определены у образцов иберо-мавританской культуры из марокканского Тафоральта (ок. 14 тыс. л. н.)[63][64].
U6d определена у энеолитического (4500—3900/3800 гг. до н. э.) образца I1187 из израильской пещеры Пкиин (Peqi’in Cave)[65].
U6a1a1 определили у образца UE2298/MS060 из исламского некрополя XI века в городе Сегорбе (Кастельон, Валенсия, Испания)[66].
U6a3 определили у образца VK116 (XII—XIII века) из Тронхейма[47].
U2’3’4’7’8’9
Субклады U2, U3, U4, U7, U8 и U9 теперь считаются монофилетическими, их общий предок «U2’3’4’7’8’9» определяется мутацией A1811G, возникшей примерно между 42 000 и 48 000 годами назад (Behar et al., 2012). В пределах U2’3’4’7’8’9, субклады U4 и U9 могут быть монофилетическими, как «U4’9» (мутации T195C!, G499A, T5999C), возникшие между 31 000 и 43 000 лет назад (Behar et al., 2012).
U2’3’4’7’8’9 был найден в останках двух древних северных сибиряков (Ancient North Siberians, ANS) возрастом 31 600 л. н. с Янской стоянки, палеолитическая стоянка человека современного типа в Арктике (Якутия)[67].
U2’3’4’7’8’9 был найден у образца Paglicci108 (28 тыс. л. н.) из итальянской пещеры Пальиччи. Граветтская культура археологическая культура кроманьонцев позднего палеолита.[13].
U2’3’4’7’8’9 был найден у образца Rigney1 (15,690-15,240 тыс. л. н.), Кластер Эль-Мирон (El Mirón Cluster), Мадленская культура во Франции[13]
U2’3’4’7’8’9 был найден у образцов эпиграветтской культуры Ориенте С (14200-13800 кал. лет назад) с острова Фавиньяна[итал.][68], у позднего эпиграветтского индивида San Teodoro 2 из пещеры Сан-Теодоро[итал.] на Сицилии[18], мезолитического индивида Oriente B (9 377 ± 25 л. н.) с острова Фавиньяна (Эгадские острова), у раннемезолитических особей из сицилийской пещеры Гротта-дель-Уззо[итал.] (провинция Трапани)[69].
U2’3’4’7’8’9 был найден у образца Balma Guilanya (~13 тыс. л. н., Испания). Географическое распространение U2’3’4’7’8’9 соответствует раннему распространению человеческих групп в Западную Европу и, как предполагается, пережило LGM в Иберийских и Апеннинских рефугиумах.[70].
U4’9 определили у образца NEO283 (23,685 л. н. до н. э.) из карстового грота Котиас Клде,Грузия[71].
U4’9 был найден у итальянского образца Tagliente 2 (коммуна Греццана) возрастом 16 980 — 16 510 л. н. (эпиграветтская культура)[72].
Гаплогруппа U2
С низкой частотой встречается по всей Европе и на Ближнем Востоке[73].
Субклады U2a, U2b и U2c являются автохтонными гаплогруппами в Индии. В наборе данных по Центральной Азии (Comas et al. 2004) из 232 линий только 2 принадлежат к U2a, 1 — к U2c[74].
Гаплогруппа U2 (U2*[75]) определена у образцов Kostenki 12(33—31,8 тыс. л. н.), Kostenki 14(38,7—36,3 тыс. л. н.), найденного на стоянке Маркина Гора Костёнковского комплекса стоянок в России[76].
U2 (субклад U2f2[77]) определили у трёх образцов со стоянки Сунгирь Sunghir 2(35,3—32,2 тыс. л. н.), Sunghir 3(35,1—32,0 тыс. л. н.), Sunghir 4(34,5—33,5 тыс. л. н.) в России[78].
U2 определили у образцов Ostuni 2(29,3 — 28,6 тыс. лет) из Италии, Goyet Q53-1(28,2 — 27,7 тыс. лет), Goyet Q375-19(27,7 — 27,3 тыс. лет), Goyet Q56-16(26,6 — 26 тыс. лет) из пещеры Гойе[фр.] (фр. Grottes de Goyet) в Бельгии. Граветтская культура (Граветт, Ла-Граветт, Лаграветтская, Граветянская) — археологическая культура кроманьонцев позднего палеолита.
U2e3 определили у охотницы-собирательницы I1960 (Tyumen_HG, 8166 л. н.) со стоянки Мергень 6 на озере Мергень (озеро) в Тюменской области (Россия)[29].
U2 обнаружена у представителей синташтинской[37] археологической культуры.
Гаплогруппа U2b2, характерная для Южной Азии, определена у представительницы хараппской (индской) культуры из Ракхигархи (2800—2300 гг. до н. э.)[79].
U2e3a определили у представителя катакомбной культуры SA6003 (2474—2211 лет до н. э.)[36].
U2e2 и U2e2a1a2 определили представителей унетицкой культуры из Чехии (Czech_EBA_Unetice 3850—4100 л. н.)[40].
U2e1a1 и U2e2a1a2 определили у участников битвы на реке Толлензе, произошедшей более 3200 лет назад[80][81].
U2b2 определили у образцов I12142 и I12446 (1000—800 лет до н. э., Katelai) из долины Сват (Пакистан)[82][83].
Один мужчина хунну из Duurlig Nars в Северо-Восточной Монголии, живший 2000 лет назад, оказался обладателем митохондриальной гаплогруппы U2e1[84].
Гаплогруппа U3
В Европе и Средней Азии распространение зафиксировано на уровне 1 % населения, на Ближнем Востоке — 2,5 %. Носителями U3 является около 6 % населения Кавказа (у сванов наличие U3 составляет 4,2 %) и от 36 % до 56 % у цыган Польши, Литвы и Испании[85][86][87].
U3a2a1 определили у неолитического образца cay011 (ок. 10 тыс. л. н.) из Чайоню (Анатолия)[88].
U3a определили у образца Ash033 (7870—7595 лет до н. э.) из Ашиклы-Хююка (Анатолия)[89].
U3a1 определили у одного из 9 убитых около 5300 года до н. э. неолитических фермеров, найденных пещере Эльс Трокс (Бисаурри, Арагон, Пиренеи)[90].
U3 определили у представителей днепро-донецкой культуры[91].
U1a’c, U3b, U3b1a, U3b3, U4a1 определили у образцов из пещеры B3 (Cave B3) в долине Бакаа (Baq҅ah Valley) в 20 км к северо-западу от Аммана (бронзовый век Иордании, 1424—1288 лет до н. э.)[92].
U3b1b определили у образца VK555 (VIII век) из корабля «Салме-2»[англ.] (волость Сальме, Эстония)[47].
U3a1 определили у женского образца VK193 (X—XII века) из Гренландии[47].
U3a1c определили у образца VK125 (XII—XIII века) из Тронхейма[47].
U3a* определили у образца KRA004 из Кракауэр-Берга (Krakauer Berg) в Восточной Германии, датирующегося возрастом примерно 612 л. н. (666—558 л. н.)[93].
Гаплогруппа U4
Субклады U4: U4a[94], U4b[95], U4c[96] и U4d[97].
Широко распространилась в Европе примерно 25 тыс. лет назад. Предполагается ближневосточное (анатолийское) происхождение субгаплогруппы U4[98][99]. Гаплогруппы U4a1, HV3, HV4, возможно, могут быть прослежены до неолита в Восточной Европе и на Кавказе. Эти линии мтДНК, вероятно, были вовлечены в позднеледниковую экспансию из восточноевропейских рефугиумов (мест выживания) после LGM (ледниковых периодов), (примерно за 12 000-19 000 лет до настоящего времени)[100]. U4a2a, U4a2* определенно моложе и датируются между 6400 и 8200 YBP, что позволяет предположить, что их расширение может быть связано с более поздними историческими событиями. Наиболее важным компонентом с точки зрения этнической истории славян является митохондриальный субкластер U4a2, скорее всего, центрально-восточноевропейского происхождения. Расширение этого подкластера можно объяснить рассредоточением культуры шнуровой керамики, которая процветала 5200-4300 л. н. в Восточной и Центральной Европе. Эта культура (также известная как культура боевых топоров) охватывала большую часть континентальной Северной Европы от реки Волги на востоке до реки Рейн на западе[100].
U4c1 определена у одного из образцов мехтоидов из Тафоральта, Марокко (19 800 лет до н. э.)[101].
U4a1 определена у образца PES001 (10 785—10 626 лет до н. э., Y-хромосомная гаплогруппа R1a5-YP1301 (под R1a1b~-YP1272)) с могильника Песчаница 1 на озере Лача[102].
U4a2b и U4c1 выявлены у представителей иберо-мавританской культуры на территории Северной Африки (10 120—8550 лет до н. э.)[103].
U4b1b1 определили у образцов BDB001 (7060-6611 calBCE), BDB002 Бад-Дюрренберг в Саксонии-Анхальт, Германия[104].
U4b определили у образца I1733 (9000—7500 лет до н. э.), Y — R1a из Василевки-на-Днепре[19].
U4 определили у образца NEO497 (10081, no 7, grave 14) (8705 лет до н. э.), Y- R1a из Василевки-на-Днепре[71].
U4a NEO537, Y-R-Y13202, U4a1, U4a2, U4d NEO536, Y-R-P297 (R1b) определили у образцов (от 8671±48 до 4600 лет до н. э.) с мезолитической стоянки культуры Веретье у озера Кубенское в деревне Минино (Вологодская область)[71][105].
U4a1 определили у образцов MN2002(ок. 8595 лет до н. э.) Y-хромосомная гаплогруппа R1a1 M459, MNN005 (ок. 8563 лет до н. э.), MNN003 (ок. 5610 лет до н. э.), Y-хромосомная гаплогруппа CT у озера Кубенское в деревне Минино, Вологодская область, Россия[106]
U4 определили у образцов BVG1 (8112 лет до н. э.) и BVG2 (8988 лет до н. э.), найденным в многослойном торфяном участке Береговая 2, расположенном примерно в 150 км к северо-западу от Екатеринбурга[107][108].
U4a1 определили у образца SF12(9033-8757 лет до н. э.), U4a2 у образца SF9 (9300-8988 лет до н. э.), могильник Stora Förvar, Stora Karlsö , у образца SBj, могильник Stora Bjers, Stenkyrka parish, остров в Балтийском море Готланд, Швеция (8963-8579 лет до н. э.)[109].
U4a2 определили у образца Lepe51(7,940-7,571 лет до н. э.) Лепенски-Вир, Сербия[110].
U4 определили у образца NEO17 (X82) (ок.7197 лет до н. э.), Y-I-PF6915 (I2) в образце, найденном в затопленной лодке, Hummervikholmen, Vest-Agder, Норвегия[71].
U4 определили у представителей эпохи неолита в Украине у образца Ukraine_N1=I1378 (6,469-6,293 лет до н. э.) днепро-донецкой культуры[111].
U4a2 определили у образца Spiginas4 (6440-6230 лет до н. э) из Литвы, относят к кундской культуре[112][113].
U4, U4a1 определили у представителей эпохи неолита в Латвии у образца Latvia_MN2 (6179-5750 лет до н. э.) и у образца Latvia_MN1 (6201-5926 лет до н. э.). Относят к культуре ямочно-гребенчатой керамики[111].
U4 определили у представителей эпохи мезолита или неолита в Карелии Оленеостровский могильник (Онежское озеро) (ок.7500 лет до н. э.)[114].
U4a определили у образца NEO671(F1586) (6709 лет до н. э.), Y-хромосомная гаплогруппа R-V88 (R1b), Железные Ворота, Schela Cladovei, Румыния[71].
U4a2 или U4d определили у представителя хвалынской культуры, жившего 6700 лет назад[115].
U4d определили у образца NEO557(No 4, 8625, burial 11) (5900 лет до н. э.), Y-хромосомная гаплогруппа R-Y13202 (R1b), U4a1 у образца NEO556(No 3, 8623, burial 7) (5900 лет до н. э.) Каргапольская культура, Караваиха,Россия[71].
U5b1d1, U4a, U2e1 и архаичная Y-хромосомная гаплогруппа R1a5 (R1a1b~-YP1272) у представителей культуры гребенчатой керамики из местонахождения Кудрукюла(эст. Kudruküla, устье реки Нарва 5600 лет назад)[116].
U4a2a определили у образца NEO212(ок. 5,443 лет до н. э.), Y-хромосомная гаплогруппа I2a, NEO204(ок. 5,278 лет до н. э.), Y-хромосомная гаплогруппа I-P220 (I2a), Днепро-донецкая культура, Голубая Криница, Россия[71].
U4a определили у образца I2074 (5,602-5,376 лет до н. э.) и I0992 (5,002-4,730 лет до н. э.) могильник Корчуган 1, погр. 1, Васьково 4, Межгорная котловина между отрогами Алтая и Саян;Новосибирская область, Россия[117]
U4d определили у образца NEO270(ок. 5,343 лет до н. э.), Y-хромосомная гаплогруппа I-Y24472 (I2a) эпохи неолита, Мамай гора, Украина[71].
U4a1 определили у образца 2017/01(5,300—5,050 л. н.) со стоянки Riņņukalns на реке Салаца на севере Латвии[118].
U4a2 определили у образца NEO814(ок. 5,165 лет до н. э.), эпохи мезолита, Bodal K, Зеландия, Дания[71].
U4a1 определили у образца NEO185 (ок. 5,078 лет до н. э.), Y-Q-YP761 представителя льяловской культуры; U4b1b1 у образца NEO194(ок. 3,647 лет до н. э.), Y-R-M479 (R2) со стоянки Сахтыш IIA, Ивановская область, Россия[71].
U4a2 определили у образца NEO746 (ок. 5,036 лет до н. э.) Henriksholm-Bøgebakken (Vedbæk), Y — I-L596Зеландия,Дания[71].
U4a1 определили у образца NEO78 (ок. 5,003 лет до н. э.), Y-Q-YP1669 (Q1a) эпохи неолита Омская стоянка 2, Омская область, Россия[71].
U4a образец NEO302(N134, Ukraine; n7)(ок. 4,889 лет до н. э.), I-L699 (I2a) могильник Волненский и NEO305(N133, Ukraine; n15)(ок. 4,661 лет до н. э.), I-L1443 могильник Васильевский, Днепро-донецкая культура, Украина[71].
U4a1 определили у представителя эпохи неолита (GE83 (KOP002)(5,200 — 4,850 лет до н. э.) на острове Эланд в Швеции[119].
U4 определили у представителей эпохи неолита (Ajv4, Fri24, Ire5, Ire4) на острове Готланд в Швеции (5000 — 4400 лет до н. э.)[119].
U4a1 определили у образца NEO498(ок. 5,106 лет до н. э.), Y-R1b, эпохи мезолита и у NEO508(ок. 4,354 лет до н. э.) эпохи неолита, Вовниги2, Украина[71].
U4a1 определили у образцов I5875 (5,291-5,060 лет до н. э.) и I5892 (5301-4982 лет до н. э.) могильник Дереивка, Украина[120].
U4a2 определили у образцa C3349 (5,043-4,019 лет до н. э.), U4b1a1a1 определили у образцa C3350 (2722—2426 лет до н. э.) могильник Songshugou уезда Зимунай в округе Алтай Синьцзян-Уйгурского автономного района Китая.[121].
U4d3 определили у представителя чемурчекской культуры, образец C1707 (4,814-4,450 лет до н. э.) могильник Bolati уезда Бурчун в округе Алтай Или-Казахского автономного округа Синьцзян-Уйгурского автономного района Китая.[121].
U4a1 определили у представителя чемурчекской культуры, образец C1703 (4,200 лет до н. э.) могильник Tuoganbai уезда Каба в округе Алтай Синьцзян-Уйгурского автономного района КНР[121].
U4a1 определили у образцов Zv162 (4470 ± 72 лет до н. э.) и Zv317 (3,890 ± 67 лет до н. э.) у представителя кундской культуры из могильника Звейниеки, на севере Латвии, на северном берегу озера Буртниекс в устье реки Руя.[105].
U4b1b2 определили у представительницы трипольской культуры этапа BI-II KYT-SSX (4,049-3,820 лет до н. э.) из урочища Коломийцев Яр близ Копачова (Обуховский район Киевской области)[122].
U4d1 определили у представителя унетицкой культуры из Чехии (Czech_EBA_Unetice 3,850—4,100 л. н.)[40].
U4a1 определили у образцов могильника Большой в Карелии (Мурманская область) (ок.3,500 лет до н. э.)[114].
U4a определили у образца NEO83(ок. 3,716 лет до н. э.), R-L1432(R1b) эпохи неолита Боровянка 17, Омская область, Россия[71].
U4a1 определили у женщины с позднетрипольской стоянки Gordinești в Молдавии (3,500—3,100 лет до н. э.)[123].
U4c1 определили у образцов SIJ003(BZNK-279/1)(3,349-3,033 лет до н. э.), SIJ002(BZNK-278/1)(3,349-3,033 лет до н. э.) Y-L, SIJ001(BZNK-278/1)(3,329-3,022 лет до н. э.), Синюха могильник поздней майкопской культуры в Адыгее, Россия[36].
U4a2 определили у образцов VEK006, VEK007 Y-J2, VEK008 (3,000-2,800 лет до н. э.) Курганы «Харман-тепе», Куро-араксская культура, село Великент в Дербентском районе Дагестана[36].
U4a2 определили у образца MK3003(BZNK-051/1)(2,577-2,476 лет до н. э.) из курганного могильника «Маринская-3», курган 1, м. 7, U4d3, Y DNA — R1b1a2 у образца RK4002(BZNK-304/2)(2,831-2,489 лет до н. э.) из курганного могильника «Расшеватская-4», Катакомбная культурно-историческая общность, Россия[36]
U4a2f определили у образца NEO121(MAN 15, A, 101)(ок. 2,577 лет до н. э.), Y- I-Z27038 (I2a) эпохи палеолита, пещера Grotte Mandrin, Rhone-Alpes, Франция[71].
U4a1 определили у образца IVA001 (2,864-2,496 лет до н. э.) с могильника Ивановогорский фатьяновской культуры, Россия[102].
U4d3 определили у представителя катакомбной культуры RK4002 (2,831—2,489 лет до н. э.)[36].

- 3000 г. до н. э.: первоначальная миграция на восток, положившая начало афанасьевской культуре, возможно, прототохарской.
- 2900 г. до н. э.: миграции на северо-запад, несущие культуру шнуровой керамики, превращаясь в культуру колоколовидных кубков, возможно, предок итало-кельтов (оспаривается).
- 2700 г. до н. э.: миграция шнуровиков на восток, переходящая в Фатьяново-Баланово (2800 г. до н. э.) → Абашево (2200 г. до н. э.) → Синташта (2100—1900 гг. до н. э.) → Андроново (1900—1700 гг. до н. э.) .
U4a2 определили у образцa C4255 (2,678-2,356 лет до н. э.) могильник Eminhuojierte уезда Дурбульджин в округе Чугучак Синьцзян-Уйгурского автономного района КНР[121].
U4 определили у представителей ямной культуры, андроновской культуры[37].
U4d1 определили у образца NEO51(MHM 4555, Grave 105)(ок. 2,391 лет до н. э.), Баальбергская культура, Kastanjegården, Skåne Швеция[71].
U4a2 определили у образцa C3319 (-), U4b1a4 определили у образцa C3313 (2,345-2,159 лет до н. э.) могильник Caishichang уезда Нилки в составе Или-Казахского автономного округа Синьцзян-Уйгурского автономного района КНР[121].
U4a1 определили у образца I0231= SVP3 в могильнике Екатериновка, Самарская область Ямная культура, (2,310 год до н. э.)[124].
U4b1a1a1 определили у образца NEO737(individ X, PMD 206)(ок. 2,161 лет до н. э.), Y- R-P310 (R1b) и U4c1 определили у образца NEO735(Individ III, PMD 200)(ок. 2,045 лет до н. э.), Y-R-P310 (R1b), эпоха среднего неолита B/позднего неолита, Borreby, Зеландия, Дания[71].
U4a2 определили у образца NEO76(IPDN 48-1, pit 3)(ок. 2,117 лет до н. э.) эпохи Неолита — Халколита, Остров2, Тюменская область, Россия[71].
U4a2 определили у образца NEO878(Kyndeløse XIII, not in PMD)(ок. 2,088 лет до н. э.) эпоха среднего неолита/позднего неолита, Kyndeløse, Зеландия, Дания[71].
U4a1a определили у образца NEO946(NM 271/45, AS 30/45, Reol: 33:12)(ок. 1,195 лет до н. э.), Q-F1213 эпоха среднего неолита A Hove Å, Зеландия, Дания[71].
U4b1a4 определили у образцa C387 (1,727-1,575* лет до н. э.), U4a2 определили у образцa C401 (1,727-1,575* лет до н. э.) могильник Hetian уезда Лоп в округе Хотан Синьцзян-Уйгурского автономного района КНР.[121].
U4a1a определили у образцов ARZ-T25, ARZ-T29 (700 ± 25 н. э.) в курганах Аржан-1 и Аржан-2 у представителей алды-бельской[англ.] (части уюкской культуры) скифо-сибиряков железного века[125][126].
U4d2 определили представителя неволинской культуры Bartym3/Bartim16/B (428—591 гг., Phase II) из Бартыма (Пермский край) и у образца с озера Уелги (Челябинская область)[127]. Носителей считают представителями древних венгров, которые произошли из Уральского региона России и мигрировали через Среднее Поволжье и восточноевропейские степи в Карпатский бассейн в девятом веке нашей эры.[128].
U4a1 определили представителя пазырыкской культуры Ak-Alakha-5, mound 4, burial 2 (400-300лет до н. э.), Алтай, плоскогорье Укок, курганы в долине реки Акалаха[129].
U4, U4a3 определили у представителей тагарской культуры[130].
U4a1 и U4a2 встречается в современной популяции Центральной и Восточной Европы (русские, белорусы, украинцы, поляки и словаки) — в среднем 4,6 %; Северо-Восточная Европы (финны, карелы, эстонцы, латыши, литовцы) — в среднем 2,5 %; Волго-Уральского региона(марийцы, коми-зыряне, коми-пермяки, мордва, удмурты, чуваши, татары, башкиры) — в среднем 6,7 %. Центральной и Западной Европы (британцы, немцы, чехи, австрийцы, швейцарцы, боснийцы, словенцы, итальянцы, французы, испанцы и португальцы)- в среднем 1,1 %[100].
U4 более 3 % встречается в некоторых современных популяциях Испании: Кантабрия — 3,8 %; Каталония — 3,8 %; Франции: Бирон (Атлантические Пиренеи) — 6,2 %; Сарта и Мен и Луара — 4,4 %; Атлантическая Луара — 4 %; Великобритания: Корнуолл — 4,3 %; в некоторых частях Германии до 6,3 %; в Австрии — 4 %; в Швейцарии — 3,9 %[131]
Гаплогруппа U4a1 характеризуется также западно-сибирским распространением, встречающимся с самой высокой частотой (7-21 %) в таких популяциях, как кеты[100][117] и U4a, U4c у народа нганасаны[132]. В другом исследовании указано на распространение гаплогруппы U4 с беспрецедентно высокой частотой — 28,9 % у кетов, 20,8 % у энцев/нганасан и 16,3 % у манси.[133]
На Кавказе и Турции U4 встречается в Грузии (18 %), в Армении (6,8 %), в Турции (1 %)[99].
Из современных народов за пределами Европы и Кавказа U4 встречается особенно в Иране (3 %) и по всей Центральной Азии, особенно в Кыргызстане (3 %), Туркменистане (3 %), Узбекистане (2,5 %) и Казахстане (2 %), а также в части Сибири, особенно в Республике Алтай (5 %) и среди носителей языков ханты и манси (12 %). U4 также встречается с высокой частотой у некоторых этнических групп в Пакистане и Афганистане, в том числе у белуджей (2,5 %), хунза бурушо (4,5 %), хазарейцев (8 %), парсов (13,5 %) и особенно у калашей Гиндукуша (34 %)[134][135][136].
Описание Ульрике (Ulrike)(U4) от Брайана Сайкса: Клан Ульрике по-немецки «Хозяйка всего») не входит в число первоначальных кланов «Семи дочерей Евы» (семь главных митохондриальных европейских родословных), но, имея в своем составе чуть менее 2 % европейцев, он претендует на то, чтобы быть включенным в число численно важных кланов. Ульрике жила около 18 000 лет назад в холодных убежищах Украины на северных границах человеческого обитания. Хотя потомки Ульрике редко встречаются, клан сегодня встречается в основном на востоке и севере Европы с особенно высокой концентрацией в Скандинавии и странах Балтии.[137]
Гаплогруппа U7
Во многих европейских популяциях гаплогруппа U7 отсутствует, но на Ближнем Востоке частота её распространения превышает 4 %, достигая 10 % среди жителей Ирана и вновь опускаясь до 5 % в Пакистане. На западе Индии к носителям этой гаплогруппы относится 12 % населения, но в остальных регионах страны — только 2 %. Родиной U7 считают Иран или западную Индию[138].
U7 определили у неолитического образца Tutkaul 1 (8419—8026 лет до настоящего времени, Y-хромосомная гаплогруппа Q1b2a, ANE) со стоянки Туткаул в Таджикистане[139].
Одна из женщин, похороненных в Осебергском корабле, имела гаплогруппу U7.
Гаплогруппа U8
- U8 определена у позднепалеолитического образца BK-1653 возрастом ок. 34,5 тыс. л. н. из пещеры Бачо Киро в Болгарии[140]
- U8c обнаружена у палеолитического образца Vestonice 13 из Дольни-Вестонице (Чехия), жившего 31 тыс. лет назад[11]
- U8c обнаружена у образца Sunghir 1 (33,9—31,8 тыс. л. н.) со стоянки Сунгирь[78]
- U8c определили у представителя граветтской культуры Paglicci 12 из Италии[18]
- U8a определили у представителей мадленской культуры[13]
- U8a специфична для басков. Поскольку в Северной Африке отсутствует, считается, что она занесена из Западной Азии[2]
- U8b1a2 и U8b1b определили у представителей трипольской культуры (3700—3500 лет до н. э.)[141]
- U8a1 обнаружен у двух представителей унетицкой культуры[142]
- U8b найдена в Италии и Иордании[2]
- U8b1a2 обнаружен у двух представителей майкопской культуры[143]
Гаплогруппа UK
Считается, что носители гаплогруппы UK мало восприимчивы к синдрому приобретённого иммунного дефицита[54].
См. также
| Митохондриальная Ева | ||||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||
| L0 | L1 | L2 | L3 | L4 | L5 | L6 | L7 | |||||||||||||||||||
| | | ||||||||||||||||||||||||||
| M | N | |||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||
| CZ | D | E | G | Q | R | O | A | S | X | Y | N1 | N2 | ||||||||||||||
| | | | | | | | | |||||||||||||||||||||||
| C | Z | B | F | R0 | R2'JT | P | UK | I | N1a | W | ||||||||||||||||
| | | | | | | ||||||||||||||||||||||||
| HV | JT | U | K | |||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||
| H | V | J | T | Устаревшие кластеры IWX | ||||||||||||||||||||||
Примечания
- ↑ 1 2 3 The Genographic Project at Архивная копия от 5 апреля 2008 на Wayback Machine National Geographic
- ↑ 1 2 3 González A. et al. The mitochondrial lineage U8a reveals a Paleolithic settlement in the Basque country. BMC Genomics, 2006. Дата обращения: 5 января 2009. Архивировано 18 февраля 2009 года.
- ↑ Raghavan M. et al. Upper Palaeolithic Siberian genome reveals dual ancestry of Native Americans Архивная копия от 29 октября 2018 на Wayback Machine, 2014
- ↑ Elena Essel et al. Ancient human DNA recovered from a Palaeolithic pendant Архивная копия от 7 мая 2023 на Wayback Machine // Nature, 03 May 2023 (Fig. 3: Ancient human mtDNA and nuclear DNA isolated from DCP1. Архивная копия от 7 мая 2023 на Wayback Machine)
- ↑ mtDNA Haplogroup U1a page at cagetti.com. Дата обращения: 5 января 2009. Архивировано 22 января 2010 года.
- ↑ J. Eldon Molto et al. Complete Mitochondrial Genome Sequencing of a Burial from a Romano-Christian Cemetery in the Dakhleh Oasis, Egypt: Preliminary Indications Архивная копия от 3 ноября 2018 на Wayback Machine, 2017
- ↑ 1 2 Sanni Översti et al. Human mitochondrial DNA lineages in Iron-Age Fennoscandia suggest incipient admixture and eastern introduction of farming-related maternal ancestry Архивная копия от 17 ноября 2019 на Wayback Machine, 15 November 2019
- ↑ Barbujani G, Bertorelle G. «Genetics and the population history of Europe» // Proceedings of the National Academy of Sciences, 2001. [1]
- ↑ Alessandro Achilli et al. (2005) Saami and Berbers—An Unexpected Mitochondrial DNA Link Архивная копия от 19 января 2020 на Wayback Machine
- ↑ Sanni Översti, Päivi Onkamo, Monika Stoljarova, Bruce Budowle, Antti Sajantila, Jukka U. Palo. Identification and analysis of mtDNA genomes attributed to Finns reveal long-stagnant demographic trends obscured in the total diversity, 2017
- ↑ 1 2 3 Fu Q. et al. A revised timescale for human evolution based on ancient mitochondrial genomes, Current Biology, 21 March 2013.
- ↑ Gabriele Scorrano et al. Genomic ancestry, diet and microbiomes of Upper Palaeolithic hunter-gatherers from San Teodoro cave (Sicily, Italy) Архивная копия от 11 декабря 2021 на Wayback Machine, December 08, 2021
- ↑ 1 2 3 4 Qiaomei Fu et al. The genetic history of Ice Age Europe, 2016
- ↑ Jones, E. R. et al. Upper Palaeolithic genomes reveal deep roots of modern Eurasians Архивная копия от 21 августа 2016 на Wayback Machine, 2015
- ↑ Peter de Barros Damgaard et al. Table S15. Mitochondrial DNA lineages identified in 74 ancient samples sequenced in the present study with Haplogrep Архивная копия от 24 июня 2018 на Wayback Machine // Supplementary Materials for: The first horse herders and the impact of early Bronze Age steppe expansions into Asia, 29 Jun 2018
- ↑ Eppie R. Jones et al. The Neolithic Transition in the Baltic Was Not Driven by Admixture with Early European Farmers // Current Biology, February 02, 2017
- ↑ New insights on Final Epigravettian funerary behavior at Arene Candide Cave (Western Liguria, Italy) Архивная копия от 5 сентября 2021 на Wayback Machine, June 2018
- ↑ 1 2 3 Alessandra Modi et al. More data on ancient human mitogenome variability in Italy: new mitochondrial genome sequences from three Upper Palaeolithic burials Архивная копия от 5 сентября 2021 на Wayback Machine // Annals of Human Biology. Volume 48, 2021 — Issue 3
- ↑ 1 2 3 Iain Mathieson et al. The Genomic History Of Southeastern Europe Архивная копия от 6 июня 2020 на Wayback Machine, 2017
- ↑ Jamie Hodgkins et al. An infant burial from Arma Veirana in northwestern Italy provides insights into funerary practices and female personhood in early Mesolithic Europe Архивная копия от 20 декабря 2021 на Wayback Machine // Scientific Reports volume 11, Article number: 23735, 14 December 2021
- ↑ Natalija Kashuba et al. Ancient DNA from mastics solidifies connection between material culture and genetics of mesolithic hunter-gatherers in Scandinavia Архивная копия от 9 января 2020 на Wayback Machine, 2019
- ↑ Lenta.ru: Прогресс: ДНК рассказала о связях между древними европейцами. Дата обращения: 30 июня 2012. Архивировано 2 июля 2012 года.
- ↑ Jens Blöcher. Genetic variation related to the adaptation of humans to an agriculturalist lifestyle // Dissertation, Mainz, Johannes Gutenberg-Universität, 2019
- ↑ Zuzana Hofmanová et al. Between fishing and farming: palaeogenomic analyses reveal cross-cultural interactions triggered by the arrival of the Neolithic in the Danube Gorges, June 28, 2022
- ↑ Iosif Lazaridis et al. «Ancient human genomes suggest three ancestral populations for present-day Europeans» Архивная копия от 12 февраля 2021 на Wayback Machine, 2013 (PDF Архивная копия от 16 марта 2021 на Wayback Machine)
- ↑ Новые генетические данные говорят о трояком происхождении европейцев. Дата обращения: 9 февраля 2020. Архивировано 4 марта 2016 года.
- ↑ Brown-skinned, blue-eyed, Y-haplogroup C-bearing European hunter-gatherer from Spain (Olalde et al. 2014). Дата обращения: 24 октября 2014. Архивировано 25 мая 2018 года.
- ↑ Federico Sánchez-Quinto et al. Genomic affinities of two 7,000-year-old Iberian hunter-gatherers Архивная копия от 16 мая 2019 на Wayback Machine, 2012 Aug 21
- ↑ 1 2 3 Vagheesh M. Narasimhan et al. The formation of human populations in South and Central Asia (англ.) // Science. — 2019-09-06. — Vol. 365, iss. 6457. — ISSN 1095-9203 0036-8075, 1095-9203. — doi:10.1126/science.aat7487. Архивировано 4 апреля 2021 года. (bioRxiv Архивная копия от 17 марта 2021 на Wayback Machine)
- ↑ U5b1b-a MTree
- ↑ Olalde, I., Brace, S., Allentoft, M. et al. The Beaker phenomenon and the genomic transformation of northwest Europe. Nature (21 февраля 2018). Дата обращения: 18 марта 2021. Архивировано 14 февраля 2021 года.
- ↑ Iain Mathieson et al. Eight thousand years of natural selection in Europe Архивная копия от 3 марта 2016 на Wayback Machine, 2015
- ↑ Inigo Olalde et al. A common genetic origin for early farmers from Mediterranean Cardial and Central European LBK cultures Архивная копия от 5 сентября 2015 на Wayback Machine, 2015
- ↑ Maïté Rivollat et al. Ancient DNA gives new insights into a Norman Neolithic monumental cemetery dedicated to male elites, April 21, 2022
- ↑ Pere Gelabert et al. Genomes From Verteba Cave Suggest Diversity Within The Trypillians In Ukraine Архивная копия от 5 января 2022 на Wayback Machine, November 2021
- ↑ 1 2 3 4 5 6 Chuan-Chao Wang et al. Ancient human genome-wide data from a 3000-year interval in the Caucasus corresponds with eco-geographic regions Архивная копия от 15 июля 2021 на Wayback Machine, 04 February 2019 (Supplementary Data 1)
- ↑ 1 2 3 4 5 Allentoft ME et al. Population genomics of Bronze Age Eurasia. (англ.) // Europe PMC. — 2015. — doi:10.1038/nature14507. Архивировано 9 мая 2020 года.
- ↑ Nikitin, A. G. Mitochondrial haplogroup C in ancient mitochondrial DNA from Ukraine extends the presence of East Eurasian genetic lineages in Neolithic Central and Eastern Europe, 2012
- ↑ Julian Susat et al. A 5,000-year-old hunter-gatherer already plagued by Yersinia pestis Архивная копия от 2 июля 2021 на Wayback Machine // Cell. Volume 35, ISSUE 13, 109278, June 29, 2021
- ↑ 1 2 3 Nick Patterson et al. Large-scale migration into Britain during the Middle to Late Bronze Age Архивная копия от 1 января 2022 на Wayback Machine // Nature, 22 December 2021
- ↑ Odile Loreille et al. Biological Sexing of a 4000-Year-Old Egyptian Mummy Head to Assess the Potential of Nuclear DNA Recovery from the Most Damaged and Limited Forensic Specimens Архивная копия от 21 апреля 2021 на Wayback Machine, Genes 2018, 9(3), 135; doi:10.3390/genes9030135
- ↑ 4000-year-old hair from the Middle Nile highlights unusual ancient DNA degradation pattern and a potential source of early eastern Africa pastoralists, 03 December 2022
- ↑ Elizabeth A. Matisoo-Smith et al. A European Mitochondrial Haplotype Identified in Ancient Phoenician Remains from Carthage, North Africa Архивная копия от 8 марта 2020 на Wayback Machine, 2016
- ↑ У карфагенян нашли первобытные европейские корни: История: Наука и техника: Lenta.ru. Дата обращения: 26 мая 2016. Архивировано 3 июня 2016 года.
- ↑ Jakub M. Niebylski et al. Unveiling Hunnic legacy: Decoding elite presence in Poland through a unique child’s burial with modified cranium, 2024
- ↑ Linea Melchior, Toomas Kivisild, Niels Lynnerup, Jørgen Dissing. Evidence of Authentic DNA from Danish Viking Age Skeletons Untouched by Humans for 1,000 Years Архивная копия от 10 апреля 2022 на Wayback Machine, May 28, 2008
- ↑ 1 2 3 4 5 Ashot Margaryan et al. Population genomics of the Viking world Архивная копия от 26 марта 2021 на Wayback Machine, 2020 (bioRxiv Архивная копия от 12 февраля 2020 на Wayback Machine)
- ↑ Marc Haber et al. A Transient Pulse of Genetic Admixture from the Crusaders in the Near East Identified from Ancient Genome Sequences Архивная копия от 31 мая 2019 на Wayback Machine, 2019
- ↑ Мустафин Х. Х., Энговатова А. В., Альборова И. Э., Тарасова А. А. Палеогенетическая экспертиза останков из одного массового захоронения 1238 года в Ярославле Архивная копия от 7 марта 2022 на Wayback Machine // Археология Подмосковья. Материалы научного семинара. Выпуск 18. М.: Институт археологии РАН, 2022. С. 111
- ↑ Chao Ning, Shizhu Gao, Boping Deng, Hongxiang Zheng, Dong Wei. Ancient mitochondrial genome reveals trace of prehistoric migration in the east Pamir by pastoralists (англ.) // Journal of Human Genetics. — 2015. — Vol. 61, iss. 2. — P. 103–108. — ISSN 1434-5161. — doi:10.1038/jhg.2015.128. Архивировано 12 мая 2020 года.
- ↑ Min-Sheng Peng, Weifang Xu, Jiao-Jiao Song, Xing Chen, Xierzhatijiang Sulaiman. Mitochondrial genomes uncover the maternal history of the Pamir populations // European Journal of Human Genetics. — 2018-1. — Т. 26, вып. 1. — С. 124–136. — ISSN 1018-4813. — doi:10.1038/s41431-017-0028-8. Архивировано 8 мая 2020 года.
- ↑ Maja Krzewińska et al. Related in Death? Further Insights on the Curious Case of Bishop Peder Winstrup and His Grandchild’s Burial Архивная копия от 4 февраля 2024 на Wayback Machine, 25 January 2024
- ↑ Maja Krzewińska et al. Related in death? A curious case of a foetus hidden in bishop Peder Winstrup’s coffin in Lund, Sweden Архивная копия от 31 марта 2022 на Wayback Machine, 2021
- ↑ 1 2 Mitochondrial DNA haplogroups influence AIDS progression. Дата обращения: 3 октября 2017. Архивировано 5 марта 2016 года.
- ↑ Erwan Pennarun et al. Divorcing the Late Upper Palaeolithic demographic histories of mtDNA haplogroups M1 and U6 in Africa Архивная копия от 20 сентября 2015 на Wayback Machine, 2012
- ↑ 1 2 3 4 Maca-Mayer N. Mitochondrial DNA transit between West Asia and North Africa inferred from U6 phylogeography // BMC Genetics, 2003
- ↑ Александр Марков. Эволюция человека: обезьяны, кости и гены. 2011
- ↑ Olivieri A., Achilli A., Pala M., Battaglia V., Fornarino S.. et al. (2006) The mtDNA Legacy of the Levantine Early Upper Paleolithic in Africa. Science 314: 1767
- ↑ Adimoolam Chandrasekar et al. Updating Phylogeny of Mitochondrial DNA Macrohaplogroup M in India: Dispersal of Modern Human in South Asian Corridor Архивная копия от 31 августа 2021 на Wayback Machine, October 20
- ↑ Supplementary Materials for. Table S16. Y-haplogroup assignment for six Taforalt males. All individuals could be assigned to haplogroup E1b1b, and five of them more specifically to E1b1b1a1 (M-78) Архивная копия от 18 марта 2018 на Wayback Machine // Loosdrecht el al. Pleistocene North African genomes link Near Eastern and sub-Saharan African human populations, 2018
- ↑ Обратная миграция в Африку произошла ещё в верхнем палеолите Архивная копия от 15 мая 2021 на Wayback Machine, 02.06.2016
- ↑ Iosif Lazaridis et al. Paleolithic DNA from the Caucasus reveals core of West Eurasian ancestry Архивная копия от 22 сентября 2018 на Wayback Machine, 2018
- ↑ M. van de Loosdrecht el al. Pleistocene North African genomes link Near Eastern and sub-Saharan African human populations, Science (2018)
- ↑ Ancient Epigenomics. Дата обращения: 18 марта 2018. Архивировано 17 марта 2018 года.
- ↑ Éadaoin Harney et al. Ancient DNA from Chalcolithic Israel reveals the role of population mixture in cultural transformation Архивная копия от 20 августа 2018 на Wayback Machine, 2018
- ↑ Marina Silva et al. Biomolecular insights into North African-related ancestry, mobility and diet in eleventh-century Al-Andalus Архивная копия от 25 сентября 2021 на Wayback Machine, 13 September 2021
- ↑ Martin Sikora et al. The population history of northeastern Siberia since the Pleistocene Архивная копия от 24 октября 2018 на Wayback Machine
- ↑ Giulio Catalano Late Upper Palaeolithic hunter-gatherers in the Central Mediterranean: new archaeological and genetic data from the Late Epigravettian burial Oriente C (Favignana, Sicily) Архивная копия от 5 марта 2021 на Wayback Machine, 2019
- ↑ Alessandra Modi et al. Paleogenetic and morphometric analysis of a Mesolithic individual from Grotta d’Oriente: An oldest genetic legacy for the first modern humans in Sicily Архивная копия от 1 ноября 2020 на Wayback Machine, 2020
- ↑ Villalba-Mouco, V., van de Loosdrecht, M.S., Rohrlach, A.B. et al. 23,000-year-old southern Iberian individual links human groups that lived in Western Europe before and after the Last Glacial Maximum. Архивная копия от 3 февраля 2024 на Wayback Machine, 2022
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 Morten E. Allentoft et al. Population Genomics of Stone Age Eurasia Архивная копия от 26 мая 2022 на Wayback Machine, 2022
- ↑ Eugenio Bortolini et al. Early Alpine human occupation backdates westward human migration in Late Glacial Europe, April 21, 2021 (bioRxiv Архивная копия от 22 апреля 2021 на Wayback Machine, August 10, 2020)
- ↑ FTDNA mtDNA U2 Haplogroup project.
- ↑ Malliya gounder Palanichamy et al. Phylogeny of Mitochondrial DNA Macrohaplogroup N in India, Based on Complete Sequencing: Implications for the Peopling of South Asia Архивная копия от 5 мая 2021 на Wayback Machine, December 2004
- ↑ U2 MTree
- ↑ Молекулярная Генеалогия > Статьи > A Complete mtDNA Genome of an Early Modern Human from Kostenki, Russia Архивная копия от 1 февраля 2010 на Wayback Machine
- ↑ U2f2 Архивная копия от 1 октября 2022 на Wayback Machine // YFull MTree 1.02.16767.
- ↑ 1 2 Sikora M. et al. Ancient genomes show social and reproductive behavior of early Upper Paleolithic foragers Архивная копия от 22 декабря 2018 на Wayback Machine, Science 10.1126/science.aao1807 (2017).
- ↑ Vasant Shinde et al. An Ancient Harappan Genome Lacks Ancestry from Steppe Pastoralists or Iranian Farmers Архивная копия от 17 сентября 2019 на Wayback Machine, 2019
- ↑ Joachim Burger, Vivian Link. Genomic Data from an Ancient European Battlefield Indicates On-Going Strong Selection on a Genomic Region Associated with Lactase Persistence Over the Last 3,000 Years Архивная копия от 30 октября 2020 на Wayback Machine // Current Biology, 9 Apr 2020
- ↑ Supplemental Information, Table S1: List of samples sequenced for this study. PDF. Дата обращения: 13 сентября 2020. Архивировано 7 июня 2022 года.
- ↑ Vagheesh M Narasimhan et al. The Genomic Formation of South and Central Asia Архивная копия от 1 апреля 2018 на Wayback Machine, March 31, 2018
- ↑ Vagheesh M. Narasimhan et al. The formation of human populations in South and Central Asia Архивная копия от 4 апреля 2021 на Wayback Machine, 06 Sep 2019
- ↑ A western Eurasian male is found in 2000-year-old elite Xiongnu cemetery in Northeast Mongolia. Дата обращения: 19 января 2015. Архивировано 27 декабря 2014 года.
- ↑ Архивированная копия. Дата обращения: 5 января 2009. Архивировано 4 марта 2009 года.
- ↑ JSTOR: An Error Occurred Setting Your User Cookie
- ↑ Blackwell Synergy — Cookie Absent (недоступная ссылка)
- ↑ N. Ezgi Altınışık et al. A genomic snapshot of demographic and cultural dynamism in Upper Mesopotamia during the Neolithic Transition Архивная копия от 15 ноября 2022 на Wayback Machine, 9 Nov 2022 (bioRxiv, February 01, 2022)
- ↑ Reyhan Yaka et al. Variable kinship patterns in Neolithic Anatolia revealed by ancient genomes, April 14, 2021
- ↑ Kurt W. Alt et al. A massacre of early Neolithic farmers in the high Pyrenees at Els Trocs, Spain Архивная копия от 7 февраля 2020 на Wayback Machine, 07 February 2020
- ↑ Newton J. (2011) Ancient Mitochondrial DNA From Pre-historic Southeastern Europe Архивная копия от 31 декабря 2019 на Wayback Machine: The Presence of East Eurasian Haplogroups Provides Evidence of Interactions with South Siberians Across the Central Asian Steppe Belt
- ↑ Lily Agranat-Tamir et al. The Genomic History of the Bronze Age Southern Levant Архивная копия от 8 августа 2021 на Wayback Machine, May 28, 2020 (Table S1. Overview of the 73 Individuals Newly Reported in this Study, Related to Figure 1)
- ↑ Cody Parker et al. A systematic investigation of human DNA preservation in medieval skeletons Архивная копия от 22 декабря 2021 на Wayback Machine, 26 October 2020 (bioRxiv Архивная копия от 22 апреля 2021 на Wayback Machine, May 22, 2020)
- ↑ Phylogeny of mt-hg U4a (2011).
- ↑ Phylogeny of mt-hg U4b (2011). Дата обращения: 6 декабря 2022. Архивировано 20 июля 2013 года.
- ↑ Phylogeny of mt-hg U4c (2011). Дата обращения: 6 декабря 2022. Архивировано 17 августа 2015 года.
- ↑ Phylogeny of mt-hg U4d (2010). Дата обращения: 6 декабря 2022. Архивировано 17 августа 2015 года.
- ↑ Boris Malyarchuk. Differentiation of the Mitochondrial Subhaplogroup U4 in the Populations of Eastern Europe, Ural, and Western Siberia: Implication to the Genetic History of the Uralic Populations // Russian Journal of Genetics 40(11):1281-1287. — 2004-01. Архивировано 2 декабря 2022 года.
- ↑ 1 2 Kristiina Tambets et al. [https://www.researchgate.net/publication/251416857_The_Topology_of_the_Maternal_Lineages_of_the_Anatolian_and_Trans-Caucasus_Populations_and_the_Peopling_of_Europe_Some_Preliminary_Considerations The Topology of the Maternal Lineages of the Anatolian and Trans-Caucasus Populations and the Peopling of Europe: Some Preliminary Considerations]. — 2000-01. Архивировано 2 декабря 2022 года.
- ↑ 1 2 3 4 B. Malyarchuk et al. Mitochondrial DNA Phylogeny in Eastern and Western Slavs // Molecular Biology and Evolution, Volume 25, Issue 8, August 2008, Pages 1651–1658,. — 2008-08. Архивировано 10 ноября 2022 года.
- ↑ On the origin of Iberomaurusians: new data based on ancient mitochondrial DNA and phylogenetic analysis of Afalou and Taforalt populations, 2016. Дата обращения: 30 декабря 2016. Архивировано 20 октября 2019 года.
- ↑ 1 2 Lehti Saag, Sergey V. Vasilyev, Svetlana V. Oshibkina et al. Genetic ancestry changes in Stone to Bronze Age transition in the East European plain Архивная копия от 23 января 2021 на Wayback Machine (Table 1), July 03, 2020 (bioRxiv Архивная копия от 30 января 2021 на Wayback Machine)
- ↑ Rym Kefi et al. On the origin of Iberomaurusians: new data based on ancient mitochondrial DNA and phylogenetic analysis of Afalou and Taforalt populations Архивная копия от 20 октября 2019 на Wayback Machine, 2016
- ↑ Orschiedt, Jörg & Haak, Wolfgang & Dietl, Holger & Siegl, Andreas & Meller, Harald. The Shaman and the Infant: The Mesolithic Double Burial from Bad Dürrenberg, 2023. Дата обращения: 10 марта 2024.
- ↑ 1 2 Jens Blöcher. Genetic variation related to the adaptation of humans to an agriculturalist lifesty Архивная копия от 3 января 2021 на Wayback Machine, 2019
- ↑ Posth, C., Yu, H., Ghalichi, A; et al. Palaeogenomics of Upper Palaeolithic to Neolithic European hunter-gatherers Архивная копия от 24 марта 2023 на Wayback Machine, Nature, Published Online: March 22, 2023
- ↑ Ancient DNA: https://ancientdna.info. Map based on public dataset on www.haplogroup.info. Дата обращения: 28 января 2023. Архивировано 28 января 2023 года.
- ↑ Mikhail Zhilin al. Early art in the Urals: new research on the wooden sculpture from Shigir // Cambridge University Press. — 2018-24-04. Архивировано 14 сентября 2019 года.
- ↑ Günther T, Malmström H, Svensson EM, Omrak A, Sánchez-Quinto F, Kılınç GM et al. [2] Архивная копия от 11 июня 2023 на Wayback Machine, PLOS Biology, Published Online: January 16, 2018
- ↑ Hofmanová, Z. et al. Palaeogenomic and biostatistical analysis of ancient DNA data from Mesolithic and Neolithic skeletal remains Архивная копия от 21 августа 2023 на Wayback Machine
- ↑ 1 2 Eppie R. Jones et al. The Neolithic Transition in the Baltic Was Not Driven by Admixture with Early European Farmers, Current Biology, Published Online: February 02, 2017
- ↑ Alissa Mittnik et al. The Genetic History of Northern Europe Архивная копия от 25 апреля 2017 на Wayback Machine, 03.03.2017
- ↑ Alissa Mittniket al. The genetic prehistory of the Baltic Sea region // Nat Commun.. — 2018-30-01. Архивировано 31 мая 2022 года.
- ↑ 1 2 Clio Der Sarkissian et al. «Ancient DNA Reveals Prehistoric Gene-Flow From Siberia in the Complex Human Population History of North East Europe» Архивная копия от 15 июня 2015 на Wayback Machine, 2013
- ↑ Iain Mathieson et al. Eight thousand years of natural selection in Europe, 2015. Дата обращения: 11 октября 2015. Архивировано 3 марта 2016 года.
- ↑ Lehti Saag et al. Extensive farming in Estonia started through a sex-biased migration from the Steppe Архивная копия от 15 апреля 2017 на Wayback Machine, March 2, 2017
- ↑ 1 2 Stanislav V. Dryomov. Stanislav V. Dryomov. Mitochondrial genome diversity on the Central Siberian Plateau with particular reference to the prehistory of northernmost Eurasia // PLoS One. — 2021-01-28.
- ↑ Susat; et al. (2021). "A 5,000-year-old hunter-gatherer already plagued by Yersinia pestis". Cell Report. 35 (13). doi:10.1016/j.celrep.2021.109278. Архивировано 20 августа 2023. Дата обращения: 25 января 2024.
- ↑ 1 2 Helena Malmström et al. Ancient mitochondrial DNA from the northern fringe of the Neolithic farming expansion in Europe sheds light on the dispersion process // Philos Trans R Soc Lond B Biol Sci.. — 2015-19-017. Архивировано 9 ноября 2022 года.
- ↑ Mathieson et al. 2018The Genomic History of Southeastern Europe // Nature. — 2018-03-08. Архивировано 24 июля 2023 года.
- ↑ 1 2 3 4 5 6 Vikas Kumar et al. Bronze and Iron Age population movements underlie Xinjiang population history Архивная копия от 18 апреля 2022 на Wayback Machine, SCIENCE, Published Online: March 31, 2022
- ↑ Alexey G. Nikitin et al. Interactions between Trypillian farmers and North Pontic forager-pastoralists in Eneolithic central Ukraine, 2022
- ↑ Alexander Immel et al. Gene-flow from steppe individuals into Cucuteni-Trypillia associated populations indicates long-standing contacts and gradual admixture Архивная копия от 19 марта 2022 на Wayback Machine, 2020 (препринт на bioRxiv Архивная копия от 8 декабря 2019 на Wayback Machine)
- ↑ Архивная копия на Wayback Machine
- ↑ Choongwon Jeong et al. A dynamic 6,000-year genetic history of Eurasia’s Eastern Steppe Архивная копия от 3 ноября 2020 на Wayback Machine, 2020
- ↑ Mary, Laura (March 28, 2019). "Genetic kinship and admixture in Iron Age Scytho-Siberians". Human Genetics. 138 (4): 411—423. doi:10.1007/s00439-019-02002-y. PMID 30923892. S2CID 85542410.
The absence of R1b lineages in the Scytho-Siberian individuals tested so far and their presence in the North Pontic Scythians suggest that these 2 groups had a completely different paternal lineage makeup with nearly no gene flow from male carriers between them
- ↑ Veronika Csáky et al. Genetic insights into the social organisation of the Avar period elite in the 7th century AD Carpathian Basin Архивная копия от 12 мая 2021 на Wayback Machine // Scientific Report, 22 January 2020
- ↑ Veronika Csáky al. Early medieval genetic data from Ural region evaluated in the light of archaeological evidence of ancient Hungarians // nature.com. — 2020-05-11. Архивировано 12 апреля 2021 года.
- ↑ Gubina et al. Gubina, M.A., Kulikov, I.V., Babenko, V.N. et al. The dynamics of the composition of mtDNA haplotypes of the ancient population of the Altai Mountains from the early bronze age (3rd millennium BC) to the iron age (2nd–1st centuries BC) // Russian Journal of Genetics. — 2016-02-02. Архивировано 12 апреля 2021 года.
- ↑ Pilipenko et al. Pilipenko AS, Trapezov RO, Cherdantsev SV, Babenko VN, Nesterova MS, Pozdnyakov DV, et al. Maternal genetic features of the Iron Age Tagar population from Southern Siberia (1st millennium BC) // PLOS ONE. — 2018-09-20. Архивировано 24 мая 2023 года.
- ↑ O García and R Fregel et al. Using mitochondrial DNA to test the hypothesis of a European post-glacial human recolonization from the Franco-Cantabrian refuge // Heredity volume 106, pages37–45 (2011). — 2010-04-21. Архивировано 6 января 2023 года.
- ↑ Natalia V Volodko, Elena B Starikovskaya, Ilya Mazunin, N. P. Eltsov. Mitochondrial Genome Diversity in Arctic Siberians, with Particular Reference to the Evolutionary History of Beringia and Pleistocenic Peopling of the Americas // The American Journal of Human Genetics 82, 1084–1100, May 2008. — 2008-05. Архивировано 2 декабря 2022 года.
- ↑ Дербенева О. А. Анализ изменчивости мтДНК коренных жителей Нижней Оби и Енисея: Манси, кеты и энцы/нганасаны Архивная копия от 18 мая 2019 на Wayback Machine, 2002
- ↑ Lluís Quintana-Murci et al. Where West Meets East: The Complex mtDNA Landscape of the Southwest and Central Asian Corridor. — 2004-05. Архивировано 9 ноября 2022 года.
- ↑ Qasim Ayub, Massimo Mezzavilla, Luca Pagani, Marc Haber, Aisha Mohyuddin. The Kalash Genetic Isolate: Ancient Divergence, Drift, and Selection // American Journal of Human Genetics. — 2015-05-07. — Т. 96, вып. 5. — С. 775–783. — ISSN 0002-9297. — doi:10.1016/j.ajhg.2015.03.012. Архивировано 8 марта 2021 года.
- ↑ Olga A. Derbeneva, Elena B. Starikovskaya, Douglas C. Wallace, and Rem I. Sukernik. Traces of Early Eurasians in the Mansi of Northwest Siberia Revealed by Mitochondrial DNA Analysis. — 2002-04. Архивировано 9 ноября 2022 года.
- ↑ Брайан Сайкс, «Кровь Островов» (Bantam, 2006) страницы 118—119
- ↑ Архивированная копия. Дата обращения: 5 января 2009. Архивировано 11 июня 2011 года.
- ↑ Cosimo Posth et al. Palaeogenomics of Upper Palaeolithic to Neolithic European hunter-gatherers Архивная копия от 4 марта 2023 на Wayback Machine // Nature, 01 March 2023
- ↑ Jean-Jacques Hublin et al. Initial Upper Palaeolithic Homo sapiens from Bacho Kiro Cave, Bulgaria Архивная копия от 19 июня 2020 на Wayback Machine, 11 May 2020
- ↑ Alexey G. Nikitin, Inna Potekhina, Nadin Rohland, Swapan Mallick, David Reich, Malcolm Lillie. Mitochondrial DNA analysis of eneolithic trypillians from Ukraine reveals neolithic farming genetic roots (англ.) // PLoS ONE. — 2017. — 24 February (no. 12(2)). — doi:10.1371/journal.pone.0172952.
- ↑ Brandt G. et al. (2013), Ancient DNA Reveals Key Stages in the Formation of Central European Mitochondrial Genetic Diversity, Science, vol. 342, no. 6155 (2013), pp. 257—261.
- ↑ Sokolov A. S. et al. Six complete mitochondrial genomes from Early Bronze Age humans in the North Caucasus Архивная копия от 11 мая 2017 на Wayback Machine
| Ключевые понятия |  | |
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||
Формирование славянских народов — генофонд, археология | |||||||
|---|---|---|---|---|---|---|---|
| Основное | |||||||
| Генофонд славян |
| ||||||
| Древнеславянские археологические культуры | |||||||
Ссылки
- U YFull MTree 1.02.00 (under construction)
- PhyloTree.org — mtDNA tree Build 17 (18 Feb 2016): subtree U
- Spread of Haplogroup U, from National Geographic
- Ursula (U5)
- U6b
- Haplogroup U4 (Ulrike)
- [www.familytreedna.com/public/india The India(subcontinent)Genealogical DNA Project]
- Danish Demes Regional DNA Project: mtDNA Haplogroup U (недоступная ссылка)
- Most of the extant mtDNA boundaries in South and Southwest Asia were likely shaped during the initial settlement of Eurasia by anatomically modern humans, Mait Metspalu et al., 2004
- [www.familytreedna.com/public/mtDNA%20U7 The U7 Haplogroup Mitochondrial DNA Project]
- Mitochondrial DNA and human evolution: In search of Eve (недоступная ссылка)
- Brian Hamman’s Clan Ursula (U5 sub-group) Website
- MtDNA Haplogroup U5 Webpage at WorldFamilies.net
- Мт-ДНК гаплогруппы U в северной Финляндии (Американский журнал генетики человека за март 2000 г.) на англ.
| Ключевые понятия | | |
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||










