Ген
Ген (др.-греч. γένος — род) — в классической генетике — наследственный фактор, который несёт информацию об определённом признаке или функции организма, и который является структурной и функциональной единицей наследственности. В таком качестве термин «ген» был введён в 1907 году датским ботаником, физиологом растений и генетиком Вильгельмом Йоханнсеном[1].
После открытия нуклеиновых кислот в качестве носителя наследственной информации определение гена изменилось, и ген стали определять как участок ДНК (у некоторых вирусов — участок РНК), задающий последовательность мономеров в полипептиде либо в функциональной РНК[2].
По мере накопления сведений о строении и работе генов определение понятия «ген» продолжало изменяться, однако в настоящее время не существует универсального определения гена, которое удовлетворило бы всех исследователей[3][4][4][5]. Одно из современных определений гена звучит следующим образом: ген представляет собой последовательность ДНК, составляющие сегменты которой не обязательно должны быть физически смежными. Эта последовательность ДНК содержит информацию об одном или нескольких продуктах в виде белка или РНК. Продукты гена функционируют в составе генетических регуляторных сетей, результат работы которых реализуется на уровне фенотипа[6].
Совокупность генов организма составляют генотип. Генотип наряду с факторами окружающей среды и развитием определяют, каким будет фенотип. Передача генов потомству является основой наследования фенотипических признаков. Большинство биологических признаков является полигенным, то есть находится под влиянием многих генов. Гены могут изменяться в результате мутаций, изменяющих последовательность ДНК. Вследствие мутаций в популяции гены существуют в различных вариантах, называемых аллелями. Разные аллели гена могут кодировать различающиеся версии белка, что может проявляться фенотипически. Гены наряду с участками ДНК, не содержащими генов, входят в состав генома, представляющего собой весь наследственный материал организма.
История
Обнаружение генов как дискретных носителей наследственности

Экспериментальные доказательства наличия дискретных факторов наследственности впервые были представлены в 1865 году Грегором Менделем в докладе на заседании Общества естествоиспытателей в Брно. В 1866 году доклад был опубликован в печатном виде[7]. Грегор Мендель изучал наследование признаков у гороха, количественно отслеживая частоту признаков у родительских растений и у потомства. В скрещиваниях растений с различными признаками он проанализировал более 8000 растений. В этих экспериментах Мендель продемонстрировал независимое наследование признаков, различие между доминантными и рецессивными признаками, различие между гетерозиготами и гомозиготами, а также явление прерывистого наследования. Результаты своих экспериментов он описал математически и интерпретировал их, предположив, что существуют дискретные, несмешиваемые в потомстве, факторы наследственности.
До работы Менделя доминирующей концепцией в объяснении закономерностей наследования была концепция, которая предполагала, что признаки родителей у потомков смешиваются аналогично смешиванию жидкостей. Этой концепции следует теория пангенезиса, разработанная Чарльзом Дарвином в 1868 году, два года спустя после публикации результатов Менделя[8]. В этой теории Дарвин предположил существование очень мелких частиц, названных им «геммулами», которые смешиваются во время зачатия.
Статья Менделя осталась практически незамеченной после её публикации в 1866 году, но она получила второе «рождение» в конце XIX века, благодаря Хуго де Фризу, Карлу Корренсу и Эриху фон Чермаку, которые пришли к аналогичным выводам в своих собственных исследованиях[9]. В частности, в 1889 году Хуго де Фриз опубликовал свою книгу «Intracellular Pangenesis»[10], в которой он постулировал, что разные признаки имеют собственные наследственные носители, и что наследование специфических черт у организмов происходит при помощи частиц. Де Фрис назвал эти единицы «пангенами» (Pangens на немецком языке), использовав часть названия теории пангенеза Дарвина.
В 1909 году Вильгельм Йоханнсен ввел термин «ген»[1], а Уильям Бейтсон — термин «генетика»[11], в то время как Эдуард Страсбургер все ещё использовал термин «панген» для обозначения основной физической и функциональной единицы наследственности[12].
Открытие ДНК в качестве носителя генетической информации
Эксперименты, проведённые в 40-е годы американскими бактериологами из Рокфеллеровского института под руководством О. Эвери, показали, что молекулярным хранилищем генетической информации является ДНК. В работах по генетической трансформации пневмококков было показано, что передача признаков от одних бактерий к другим происходит при помощи только одного вещества — ДНК. Ни белок, ни другие химические компоненты клетки этим свойством не обладали [13][14][15]. В 1953 году при помощи рентгеновской кристаллографии Розалинд Франклин и Морис Уилкинс получили высококачественные снимки структуры ДНК. Эти снимки помогли Джеймсу Д. Уотсону и Фрэнсису Крику создать модель молекулы двуцепочечной спирали ДНК и сформулировать гипотезу механизма генетической репликации[16][17].
В начале 1950-х годов преобладало мнение, что гены в хромосоме действуют как отдельные объекты, неразделимые путем рекомбинации и расположенные как бусы на веревочке. Эксперименты Сеймура Бензера с использованием мутантов, дефектных бактериофагов в области rII T4[англ.] (1955—1959), показали, что отдельные гены имеют простую линейную структуру и, вероятно, эквивалентны линейному сечению ДНК[18][19].
В совокупности этот объём исследований установил центральную догму молекулярной биологии, которая утверждает, что белки транслируются с РНК, которая транскрибируется с ДНК. Эта догма с тех пор, как было показано, имеет исключения, такие как обратная транскрипция в ретровирусах. Современное исследование генетики на уровне ДНК известно как молекулярная генетика.
В 1972 году Уолтер Файерс и его команда первыми определили последовательность гена: последовательность белка оболочки Bacteriophage MS2[англ.][20]. Последующее развитие секвенирования ДНК с методом Сэнгера в 1977 году Фредериком Сангером улучшило эффективность секвенирования и превратило его в рутинный лабораторный инструмент[21]. Автоматизированная версия метода Сангера использовалась на ранних этапах проекта «Геном человека»[22].
Современный синтез и его преемники
Теории, разработанные в начале XX века для интеграции менделевской генетики с дарвиновской эволюцией, называются современным синтезом, термином, введенным Джулианом Хаксли[23].
Эволюционные биологи впоследствии модифицировали эту концепцию, такую как геноцентричный взгляд[англ.] Джорджа Уильямса на эволюцию. Он предложил эволюционную концепцию гена как единицы естественного отбора с определением: «то, что разделяет и рекомбинирует с заметной частотой»[24]:24. С этой точки зрения, молекулярный ген транскрибируется как единое целое, а эволюционный ген наследуется как единое целое. Связанные идеи, подчеркивающие центральную роль генов в эволюции, были популяризированы Ричардом Докинзом[25][26].
Молекулярная основа
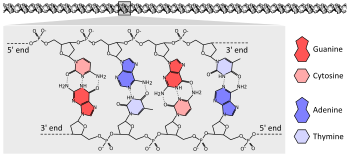
ДНК
Генетическая информация у подавляющего большинства организмов закодирована в длинных молекулах ДНК. ДНК состоит из двух спирально закрученных полимерных цепей, мономерами которых служат четыре нуклеотида: dАМФ, dГМФ, dЦМФ и dТМФ. Нуклеотиды в ДНК состоят из пятиуглеродного сахара (2-дезоксирибозы), фосфатной группы и одного из четырёх азотистых оснований: аденина, цитозина, гуанина и тимина[27]:2.1. Азотистое основание связано гликозидной связью с пятиуглеродным (пентозного) сахаром в 1'-положении. Остовом цепей ДНК служит чередующаяся последовательность пентозных сахаров и фосфатов, фосфатные группы присоединяются к сахару в 5'- и 3'-положениях. Номера позиций пентозного кольца отмечены штрихом для того, чтобы различать нумерацию колец в сахаре и азотистом основании[28].
Из-за химического состава пентозных остатков цепи ДНК имеют направленность. Один конец полимера ДНК содержит открытую гидроксильную группу на дезоксирибозе в 3'-положении; этот конец называется 3'-конец. Другой конец содержит открытую фосфатную группу, это 5'-конец. Две цепи (нити) двойной спирали ДНК ориентированы в противоположных направлениях. Синтез ДНК, в том числе при репликации ДНК, происходит в направлении 5 '→ 3', потому что новые нуклеотиды добавляются посредством реакции дегидратации, которая использует открытый 3’-гидроксил в качестве нуклеофила[29]:27.2.
Экспрессия генов, закодированных в ДНК, начинается с транскрипции нуклеотидной последовательности ДНК в последовательность нуклеотидов другого типа нуклеиновых кислот — РНК. РНК очень похожа на ДНК, но её мономеры содержат рибозу, а не дезоксирибозу. Кроме того, вместо тимина в РНК используется урацил. Молекулы РНК являются одноцепочечными и менее стабильны, чем ДНК. Гены в ДНК, а после транскрипции и в тРНК, представляют собой природный шифр (код) для синтеза белка. Каждые три нуклеотида (триплет) кодируют одну аминокислоту. К примеру если в гене 300 нуклеотидов, то в белке должно быть 100 аминокислот. Поэтому код называется триплетный. Правило, по которому определяется, какому триплету соответствует какая аминокислота, называется генетическим кодом. Считывание генетического кода происходит в рибосоме во время трансляции РНК в белок. Генетический код почти одинаков для всех известных организмов[27]:4.1.
Хромосома

Наследственный материал организма, или геном, хранится в одной или нескольких хромосомах, число которых специфично для вида. Хромосома состоит из одной очень длинной молекулы ДНК, которая может содержать тысячи генов[27]:4.2. Область хромосомы, где находится ген, называется локусом. Каждый локус содержит определённый аллель гена. Представители популяции могут отличаться по аллелям гена, находящимся в одинаковых локусах хромосом.
Большинство эукариотических генов хранятся в нескольких линейных хромосомах. Хромосомы упакованы в ядре в комплексе с белками хроматина. Наиболее многочисленными белками хроматина являются гистоны, которые формируют белковую глобулу, называемую нуклеосомой. ДНК обвивается вокруг нуклеосом, что представляет собой первый уровень упаковки ДНК в хромосоме[27]:4.2. Распределение нуклеосом вдоль ДНК, а также химические модификации самих гистонов регулируют доступность ДНК для регуляторных факторов, участвующих в репликации, транскрипции, трансляции и репарации. Помимо генов эукариотические хромосомы содержат также служебные последовательности, обеспечивающие стабильность и воспроизведение хромосом, а также их распределение между дочерними клетками в митозе. Это теломеры, сайты инициации репликации и центромера, соответственно[27]:4.2.
Функциональные определения
Трудно точно определить, в какую часть последовательности ДНК входит ген[5].
Основные характеристики гена
В молекулярной биологии установлено, что гены — это участки ДНК, несущие какую-либо целостную информацию — о строении одной молекулы белка или одной молекулы РНК, которые определяют развитие, рост и функционирование организма.
Каждый ген характеризуется рядом специфических регуляторных последовательностей ДНК[англ.], таких как промоторы, которые принимают участие в регулировании функции гена. Регуляторные последовательности могут находиться как в непосредственной близости от открытой рамки считывания, кодирующей белок, или начала последовательности РНК, как в случае с промоторами (так называемые cis-регуляторные элементы, англ. cis-regulatory elements), так и на расстоянии многих миллионов пар оснований (нуклеотидов), как в случае с энхансерами, инсуляторами и супрессорами (иногда классифицируемые как trans-регуляторные элементы, англ. trans-regulatory elements). То есть, понятие гена не ограничено только кодирующим участком ДНК. Ген представляет собой более широкую концепцию, включающую в себя и регуляторные последовательности.
Изначально термин «ген» появился как теоретическая единица передачи дискретной наследственной информации. История биологии помнит споры о том, какие молекулы могут являться носителями наследственной информации. Считалось, что такими носителями могут быть только белки, так как их строение (20 аминокислот) позволяет создать больше вариантов, чем ДНК, которая состоит всего из четырёх видов нуклеотидов. Однако экспериментально доказано, что именно ДНК содержит в себе наследственную информацию, что было выражено в виде центральной догмы молекулярной биологии: ДНК — РНК — белок.
Гены могут подвергаться мутациям — случайным или целенаправленным изменениям последовательности нуклеотидов в цепи ДНК. Мутации могут приводить к изменению последовательности, а следовательно изменению биологических характеристик белка или РНК. Результатом может стать изменённое или даже анормальное функционирование организма. Такие мутации в ряде случаев являются патогенными, так как их результатом является заболевание, или летальными на эмбриональном уровне. Далеко не все изменения последовательности нуклеотидов приводят к изменению структуры белка (благодаря эффекту вырожденности генетического кода) или к существенному изменению последовательности и не являются патогенными. В частности, геном человека характеризуется однонуклеотидными полиморфизмами и вариациями числа копий (англ. copy number variations), такими как делеции и дупликации, которые составляют около 1 % всей нуклеотидной последовательности человека[31]. Однонуклеотидные полиморфизмы, в частности, определяют различные аллели одного гена.
Молекулярная эволюция
Мутация
Репликация ДНК по большей части чрезвычайно точна, однако ошибки (мутации) случаются[27]:7.6. Частота ошибок в эукариотических клетках может составлять всего 10−8 в нуклеотиде на репликацию[32][33], тогда как для некоторых РНК-вирусов она может достигать 10−3[34]. Это означает, что в каждое поколение, каждый человек в геноме накапливает около 30 новых мутаций[35]. Небольшие мутации могут быть вызваны репликацией ДНК и последствиями повреждения ДНК и включают точечные мутации, в которых изменяется одно основание, и мутации со сдвигом рамки, в которых одно основание вставляется или удаляется. Любая из этих мутаций может изменить ген по миссенс (изменить код для кодирования другой аминокислоты) или по нонсенс (преждевременный стоп-кодон)[36]. Большие мутации могут быть вызваны ошибками в рекомбинации, чтобы вызвать хромосомные аномалии, включая дублирование, делецию, перегруппировку или инверсию больших участков хромосомы. Кроме того, механизмы восстановления ДНК могут вносить мутационные ошибки при восстановлении физического повреждения молекулы. Восстановление, даже с мутацией, является более важным для выживания, чем восстановление точной копии, например, при восстановлении двухцепочечных разрывов[27]:5.4.
Когда в популяции вида присутствует несколько различных аллелей гена, это называется полиморфизм. Большинство различных аллелей функционально эквивалентны, однако некоторые аллели могут вызывать различные фенотипические признаки. Самый распространенный аллель гена называется диким типом, а редкие аллели — мутантами. Генетические различия в относительных частотах различных аллелей в популяции обусловлены как естественным отбором, так и генетическим дрейфом[37]. Аллель дикого типа не обязательно является предком менее распространенных аллелей и не обязательно более приспособлена.
Количество генов
Размер генома и количество генов, которые он содержит, значительно варьируют у таксономических групп. Наименьший геном встречаются у вирусов[38], и вироидов (которые действуют как один некодирующий ген РНК)[39]. И наоборот, растения могут иметь очень большие геномы[40], в рисе содержатся более 46 000 генов, кодирующих белок[41]. Общее количество кодирующих белок генов (протеома Земли), которое оценивалось в 2007 году в 5 млн. последовательностей[42], к 2017 году было снижено до 3,75 млн[43].
Генная инженерия
Генная инженерия — это методы модификации генетического материала для изменения свойств живого организма. С 1970-х годов было разработано множество методов, специально предназначенных для добавления, удаления и редактирования генов в вирусах, бактериях, растениях, грибах и животных, включая человека[44]. Недавно разработанные методы геномной инженерии используют инженерные нуклеазные ферменты для создания целевой репарации ДНК в хромосоме, чтобы либо разрушить, либо отредактировать ген в процессе репарации искусственно внесённого разрыва ДНК[45][46][47][48]. Связанный термин синтетическая биология иногда используется для обозначения обширной дисциплины генной инженерии организма[49].
Генная инженерия в настоящее время является рутинным инструментом при работе с модельными организмами. Например, гены легко добавляются к бактериям[50], а линии «Knockout mouse[англ.]» мышей с нарушенной функцией определённого гена используются для исследования функции этого гена[51][52]. Многие организмы были генетически модифицированы для применения в сельском хозяйстве, промышленной биотехнологии, клеточной биологии и медицине.
У многоклеточных организмов обычно модифицируется эмбрион, который вырастает во взрослый генетически модифицированный организм[53]. Однако геномы клеток взрослого организма можно редактировать с использованием методов генной терапии для лечения генетических заболеваний.
Свойства гена
- дискретность — несмешиваемость генов;
- стабильность — способность сохранять структуру;
- лабильность — способность многократно мутировать;
- множественный аллелизм — многие гены существуют в популяции во множестве молекулярных форм;
- аллельность — в генотипе диплоидных организмов только две формы гена;
- специфичность — каждый ген кодирует свой признак;
- плейотропия — множественный эффект гена;
- экспрессивность — степень выраженности гена в признаке;
- пенетрантность — частота проявления гена в фенотипе;
- амплификация — увеличение количества копий гена[].
Классификация
- Структурные гены — гены, кодирующие информацию о первичной структуре белка. Расположение нуклеотидных триплетов в структурных генах коллинеарно последовательности аминокислот в полипептидной цепи, кодируемой данным геном (См. также статью гены домашнего хозяйства).
- Функциональные гены — гены, которые контролируют и направляют деятельность структурных генов[54].
См. также
- Непосредственно ранние гены
- Аллели
- Геном
- Генотерапия
- Реализация генетической информации
- Эпигенетика
- Репликация
- ДНК
- РНК
- Генетическая экспертиза
- Генетическая карта
- Библиотека генов
Примечания
- ↑ 1 2 Johannsen, W. (1905). Arvelighedslærens elementer («The Elements of Heredity». Copenhagen). Rewritten, enlarged and translated into German as Elemente der exakten Erblichkeitslehre (Jena: Gustav Fischer, 1909; Scanned full text. Архивная копия от 30 мая 2009 на Wayback Machine
- ↑ Тарантул В. З. Толковый словарь по молекулярной и клеточной биотехнологиию Русско-английский. — М.: Языки славянской литературы, 2015. — Т. 1. — С. 370—371. — 984 с. — ISBN 978-5-94457-249-3.
- ↑ Pearson H. Genetics: what is a gene? (англ.) // Nature. — 2006. — May (vol. 441, no. 7092). — P. 398—401. — doi:10.1038/441398a. — . — PMID 16724031.
- ↑ 1 2 Pennisi E. Genomics. DNA study forces rethink of what it means to be a gene (англ.) // Science : journal. — 2007. — June (vol. 316, no. 5831). — P. 1556—1557. — doi:10.1126/science.316.5831.1556. — PMID 17569836.
- ↑ 1 2 Gericke, Niklas Markus; Hagberg, Mariana. Definition of historical models of gene function and their relation to students' understanding of genetics (англ.) // Science & Education[англ.] : journal. — 2006. — 5 December (vol. 16, no. 7—8). — P. 849—881. — doi:10.1007/s11191-006-9064-4. — .
- ↑ Portin P., Wilkins A. The evolving definition of the term “gene” (англ.) // Genetics. — 2017. — Vol. 205, no. 4. — P. 1353—1364. Архивировано 7 ноября 2019 года.
- ↑ Noble D. Genes and causation (англ.) // Philosophical Transactions of the Royal Society of London. Series A, Mathematical and Physical Sciences[англ.] : journal. — 2008. — September (vol. 366, no. 1878). — P. 3001—3015. — doi:10.1098/rsta.2008.0086. — . — PMID 18559318. Архивировано 27 марта 2020 года.
- ↑ Magner, Lois N. A History of the Life Sciences (неопр.). — Third. — Marcel Dekker, CRC Press, 2002. — С. 371. — ISBN 978-0-203-91100-6.
- ↑ Henig, Robin Marantz. The Monk in the Garden: The Lost and Found Genius of Gregor Mendel, the Father of Genetics (англ.). — Boston: Houghton Mifflin[англ.], 2000. — P. 1—9. — ISBN 978-0395-97765-1.
- ↑ Де Фриз, Хуго, Intracellulare Pangenese, Verlag von Gustav Fischer, Йена (город), 1889. Translated in 1908 from German to English by C. Stuart Gager as Intracellular Pangenesis Архивная копия от 8 ноября 2017 на Wayback Machine, Open Court Publishing Co., Chicago, 1910
- ↑ Либацкая Т. Е. Уильям Бэтсон: у истоков генетики // Вестник Российской академии наук. — 2003. — Т. 73, № 9. — С. 830—837.
- ↑ C. Stuart Gager, Translator’s preface to Intracellular Pangenesis Архивная копия от 8 ноября 2017 на Wayback Machine, p. viii.
- ↑ Франк-Каменецкий, 2004, с. 18.
- ↑ Avery, OT; MacLeod, CM; McCarty, M. Studies on the Chemical Nature of the Substance Inducing Transformation of Pneumococcal Types: Induction of Transformation by a Desoxyribonucleic Acid Fraction Isolated from Pneumococcus Type III (англ.) // The Journal of Experimental Medicine[англ.] : journal. — Rockefeller University Press[англ.], 1944. — Vol. 79, no. 2. — P. 137—158. — doi:10.1084/jem.79.2.137. — PMID 19871359. — PMC 2135445.
- ↑ Hershey, AD; Chase, M. Independent functions of viral protein and nucleic acid in growth of bacteriophage (англ.) // The Journal of General Physiology[англ.] : journal. — Rockefeller University Press[англ.], 1952. — Vol. 36, no. 1. — P. 39—56. — doi:10.1085/jgp.36.1.39. — PMID 12981234. — PMC 2147348.
- ↑ Judson, Horace[англ.]. The Eighth Day of Creation: Makers of the Revolution in Biology (англ.). — Cold Spring Harbor Laboratory Press[англ.], 1979. — P. 51—169. — ISBN 978-0-87969-477-7.
- ↑ Watson, J.D.; Crick, F. H. Molecular Structure of Nucleic Acids: A Structure for Deoxyribose Nucleic Acid (рум.) // Nature. — 1953. — Т. 171, nr. 4356. — P. 737—738. — doi:10.1038/171737a0. — . — PMID 13054692. Архивировано 24 октября 2017 года.
- ↑ Benzer S. Fine Structure of a Genetic Region in Bacteriophage (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1955. — Vol. 41, no. 6. — P. 344—354. — doi:10.1073/pnas.41.6.344. — . — PMID 16589677. — PMC 528093.
- ↑ Benzer S. On the Topology of the Genetic Fine Structure (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1959. — Vol. 45, no. 11. — P. 1607—1620. — doi:10.1073/pnas.45.11.1607. — . — PMID 16590553. — PMC 222769.
- ↑ Min Jou W., Haegeman G., Ysebaert M., Fiers W. Nucleotide sequence of the gene coding for the bacteriophage MS2 coat protein (англ.) // Nature : journal. — 1972. — May (vol. 237, no. 5350). — P. 82—88. — doi:10.1038/237082a0. — . — PMID 4555447.
- ↑ Sanger, F; Nicklen, S; Coulson, A. R. DNA sequencing with chain-terminating inhibitors (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1977. — Vol. 74, no. 12. — P. 5463—5467. — doi:10.1073/pnas.74.12.5463. — . — PMID 271968. — PMC 431765.
- ↑ Adams, Jill U. DNA Sequencing Technologies (неопр.) // Nature Education Knowledge. — Nature Publishing Group, 2008. — Т. 1, № 1. — С. 193. Архивировано 30 октября 2019 года.
- ↑ Huxley, Julian. Evolution: the Modern Synthesis (неопр.). — Cambridge, Massachusetts: MIT Press, 1942. — ISBN 978-0262513661.
- ↑ Williams, George C. Adaptation and Natural Selection a Critique of Some Current Evolutionary Thought (англ.). — Online. — Princeton: Princeton University Press, 2001. — ISBN 9781400820108.
- ↑ Dawkins, Richard. The selfish gene (англ.). — Repr. (with corr.). — London: Oxford University Press, 1977. — ISBN 978-0-19-857519-1.
- ↑ Dawkins, Richard. The extended phenotype (англ.). — Paperback. — Oxford: Oxford University Press, 1989. — ISBN 978-0-19-286088-0.
- ↑ 1 2 3 4 5 6 7 Alberts, Bruce; Johnson, Alexander; Lewis, Julian[англ.]; Raff, Martin[англ.]; Roberts, Keith; Walter, Peter. Molecular Biology of the Cell (неопр.). — Fourth. — New York: Garland Science[англ.], 2002. — ISBN 978-0-8153-3218-3. Архивировано 27 сентября 2017 года.
- ↑ Кребс, 2017, с. 21.
- ↑ Stryer L., Berg J. M., Tymoczko J. L. Biochemistry (неопр.). — 5th. — San Francisco: W.H. Freeman[англ.], 2002. — ISBN 978-0-7167-4955-4. Архивировано 11 марта 2014 года.
- ↑ Bolzer, Andreas; Kreth, Gregor; Solovei, Irina; Koehler, Daniela; Saracoglu, Kaan; Fauth, Christine; Müller, Stefan; Eils, Roland; Cremer, Christoph; Speicher, Michael R.; Cremer, Thomas. Three-Dimensional Maps of All Chromosomes in Human Male Fibroblast Nuclei and Prometaphase Rosettes (англ.) // PLoS Biology : journal. — 2005. — Vol. 3, no. 5. — P. e157. — doi:10.1371/journal.pbio.0030157. — PMID 15839726. — PMC 1084335.

- ↑ Levy S., Sutton G., Ng P. C., Feuk L., Halpern A. L., Walenz B. P., Axelrod N., Huang J., Kirkness E. F., Denisov G., Lin Y., Macdonald J. R., Pang A. W., Shago M., Stockwell T. B., Tsiamouri A., Bafna V., Bansal V., Kravitz S. A., Busam D. A., Beeson K. Y., McIntosh T. C., Remington K. A., Abril J. F., Gill J., Borman J., Rogers Y. H., Frazier M. E., Scherer S. W., Strausberg R. L., Venter J. C. The Diploid Genome Sequence of an Individual Human (англ.) // PLoS Biol : journal. — 2007. — Vol. 5, no. 10. — P. e254. Архивировано 4 ноября 2007 года.
- ↑ Nachman M. W., Crowell S. L. Estimate of the mutation rate per nucleotide in humans (англ.) // Genetics : journal. — 2000. — September (vol. 156, no. 1). — P. 297—304. — PMID 10978293. — PMC 1461236. Архивировано 8 апреля 2011 года.
- ↑ Roach J. C., Glusman G., Smit A. F., etal. Analysis of genetic inheritance in a family quartet by whole-genome sequencing (англ.) // Science : journal. — 2010. — April (vol. 328, no. 5978). — P. 636—639. — doi:10.1126/science.1186802. — . — PMID 20220176. — PMC 3037280.
- ↑ Drake J. W., Charlesworth B., Charlesworth D., Crow J. F. Rates of spontaneous mutation (англ.) // Genetics. — 1998. — April (vol. 148, no. 4). — P. 1667—1686. — PMID 9560386. — PMC 1460098. Архивировано 21 августа 2010 года.
- ↑ Reed E. Pyeritz, Bruce R. Korf, Wayne W. Grody. Emery and Rimoin's principles and practice of medical genetics and genomics. — 7th ed. — London: Academic press, 2019. — ISBN 978-0-12-812537-3.
- ↑ What kinds of gene mutations are possible? Genetics Home Reference. United States National Library of Medicine (11 мая 2015). Дата обращения: 19 мая 2015. Архивировано 15 марта 2016 года.
- ↑ Andrews, Christine A. Natural Selection, Genetic Drift, and Gene Flow Do Not Act in Isolation in Natural Populations (англ.) // Nature Education Knowledge : journal. — Nature Publishing Group, 2010. — Vol. SciTable, no. 10. — P. 5. Архивировано 31 марта 2019 года.
- ↑ Belyi, V.A.; Levine, A.J.; Skalka, A.M. Sequences from Ancestral Single-Stranded DNA Viruses in Vertebrate Genomes: the Parvoviridae and Circoviridae Are More than 40 to 50 Million Years Old (англ.) // Journal of Virology[англ.] : journal. — 2010. — 22 September (vol. 84, no. 23). — P. 12458—12462. — doi:10.1128/JVI.01789-10. — PMID 20861255. — PMC 2976387.
- ↑ Flores, Ricardo; Di Serio, Francesco; Hernández, Carmen. Viroids: The Noncoding Genomes (неопр.) // Seminars in Virology. — 1997. — February (т. 8, № 1). — С. 65—73. — doi:10.1006/smvy.1997.0107.
- ↑ Zonneveld, B.J.M. New Record Holders for Maximum Genome Size in Eudicots and Monocots (англ.) // Journal of Botany : journal. — 2010. — Vol. 2010. — P. 1—4. — doi:10.1155/2010/527357.
- ↑ Yu J., Hu S., Wang J., Wong G. K., Li S., Liu B., Deng Y., Dai L., Zhou Y., Zhang X., Cao M., Liu J., Sun J., Tang J., Chen Y., Huang X., Lin W., Ye C., Tong W., Cong L., Geng J., Han Y., Li L., Li W., Hu G., Huang X., Li W., Li J., Liu Z., Li L., Liu J., Qi Q., Liu J., Li L., Li T., Wang X., Lu H., Wu T., Zhu M., Ni P., Han H., Dong W., Ren X., Feng X., Cui P., Li X., Wang H., Xu X., Zhai W., Xu Z., Zhang J., He S., Zhang J., Xu J., Zhang K., Zheng X., Dong J., Zeng W., Tao L., Ye J., Tan J., Ren X., Chen X., He J., Liu D., Tian W., Tian C., Xia H., Bao Q., Li G., Gao H., Cao T., Wang J., Zhao W., Li P., Chen W., Wang X., Zhang Y., Hu J., Wang J., Liu S., Yang J., Zhang G., Xiong Y., Li Z., Mao L., Zhou C., Zhu Z., Chen R., Hao B., Zheng W., Chen S., Guo W., Li G., Liu S., Tao M., Wang J., Zhu L., Yuan L., Yang H. A draft sequence of the rice genome (Oryza sativa L. ssp. indica) (англ.) // Science : journal. — 2002. — April (vol. 296, no. 5565). — P. 79—92. — doi:10.1126/science.1068037. — . — PMID 11935017.
- ↑ Perez-Iratxeta C., Palidwor G., Andrade-Navarro M. A. Towards completion of the Earth's proteome (англ.) // EMBO Reports[англ.] : journal. — 2007. — December (vol. 8, no. 12). — P. 1135—1141. — doi:10.1038/sj.embor.7401117. — PMID 18059312. — PMC 2267224.
- ↑ Mier P, Andrade-Navarro MA (2019). "Toward completion of the Earth's proteome: an update a decade later". Brief Bioinform. 20 (2): 463–470. doi:10.1093/bib/bbx127. PMID 29040399.
- ↑ Stanley N. Cohen; Annie C.Y. Chang. Recircularization and Autonomous Replication of a Sheared R-Factor DNA Segment in Escherichia coli Transformants (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1973. — 1 May (vol. 70, no. 5). — P. 1293—1297. — doi:10.1073/pnas.70.5.1293. — . — PMID 4576014. — PMC 433482.
- ↑ Esvelt, KM.; Wang, HH. Genome-scale engineering for systems and synthetic biology (англ.) // Mol Syst Biol[англ.] : journal. — 2013. — Vol. 9, no. 1. — P. 641. — doi:10.1038/msb.2012.66. — PMID 23340847. — PMC 3564264.
- ↑ Tan, WS.; Carlson, DF.; Walton, MW.; Fahrenkrug, SC.; Hackett, PB. Precision editing of large animal genomes (неопр.) // Adv Genet. — 2012. — Т. Advances in Genetics. — С. 37—97. — ISBN 9780124047426. — doi:10.1016/B978-0-12-404742-6.00002-8. — PMID 23084873. — PMC 3683964.
- ↑ Puchta, H.; Fauser, F. Gene targeting in plants: 25 years later (англ.) // Int. J. Dev. Biol.[англ.] : journal. — 2013. — Vol. 57, no. 6—7—8. — P. 629—637. — doi:10.1387/ijdb.130194hp. — PMID 24166445.
- ↑ Ran F. A., Hsu P. D., Wright J., Agarwala V., Scott D. A., Zhang F. Genome engineering using the CRISPR-Cas9 system (англ.) // Nat Protoc[англ.] : journal. — 2013. — Vol. 8, no. 11. — P. 2281—2308. — doi:10.1038/nprot.2013.143. — PMID 24157548. — PMC 3969860.
- ↑ Kittleson, Joshua. Successes and failures in modular genetic engineering (англ.) // Current Opinion in Chemical Biology : journal. — Elsevier, 2012. — Vol. 16, no. 3—4. — P. 329—336. — doi:10.1016/j.cbpa.2012.06.009. — PMID 22818777.
- ↑ Berg, P.; Mertz, J.E. Personal Reflections on the Origins and Emergence of Recombinant DNA Technology (англ.) // Genetics : journal. — 2010. — Vol. 184, no. 1. — P. 9—17. — doi:10.1534/genetics.109.112144. — PMID 20061565. — PMC 2815933.
- ↑ Austin, Christopher P.; Battey, James F.; Bradley, Allan; Bucan, Maja; Capecchi, Mario; Collins, Francis S.; Dove, William F.; Duyk, Geoffrey; Dymecki, Susan. The Knockout Mouse Project (англ.) // Nature Genetics : journal. — 2004. — September (vol. 36, no. 9). — P. 921—924. — ISSN 1061-4036. — doi:10.1038/ng0904-921. — PMID 15340423. — PMC 2716027.
- ↑ Guan, Chunmei; Ye, Chao; Yang, Xiaomei; Gao, Jiangang. A review of current large-scale mouse knockout efforts (англ.) // Genesis : journal. — 2010. — Vol. 48, no. 2. — P. 73—85. — doi:10.1002/dvg.20594. — PMID 20095055.
- ↑ Deng C. In celebration of Dr. Mario R. Capecchi's Nobel Prize (англ.) // International Journal of Biological Sciences[англ.] : journal. — 2007. — Vol. 3, no. 7. — P. 417—419. — doi:10.7150/ijbs.3.417. — PMID 17998949. — PMC 2043165.
- ↑ О.-Я.Л.Бекиш. Медицинская биология. — Минск: Ураджай, 2000. — С. 114. — 518 с.
Литература
- Инге-Вечтомов С. Г. Генетика с основами селекции: учебник для студентов высших учебных заведений. — СПб.: Изд-во Н-Л, 2010. — 720 с. — ISBN 978-5-94869-105-3..
- Молекулярная биология клетки: в 3-х томах / Б. Альбертс, А. Джонсон, Д. Льюис и др. — М.-Ижевск: НИЦ «Регулярная и хаотическая динамика», Институт компьютерных исследований, 2013. — Т. I. — 808 с. — ISBN 978-5-4344-0112-8..
- Кребс Дж. Гены по Льюину / Дж. Кребс, Э. Голдштейн, С. Килпатрик, пер. 10-го англ. изд. — М.: Лаборатория знаний, 2017. — 919 с. — ISBN 978-5-906828-24-8.
- Франк-Каменецкий М. Д. Век ДНК. — М.: Изд-во «КДУ», 2004. — 240 с. — ISBN 5-98227-017-2.
Ссылки
- Материалы по генетике от Российской Академии Наук Архивная копия от 4 ноября 2005 на Wayback Machine
- ГЕН Архивная копия от 7 апреля 2019 на Wayback Machine Большая российская энциклопедия
| Ключевые понятия |  | |
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||










