
Ферме́нты, или энзи́мы , — обычно сложные белковые соединения, РНК (рибозимы) или их комплексы, ускоряющие химические реакции в живых системах. Каждый фермент, свернутый в определённую структуру, ускоряет соответствующую химическую реакцию: реагенты в такой реакции называются субстратами, а получающиеся вещества — продуктами. Ферменты специфичны к субстратам: АТФ-аза катализирует расщепление только АТФ, а киназа фосфорилазы фосфорилирует только фосфорилазу.
Глико́лиз, или путь Эмбдена — Мейергофа — Парнаса — процесс окисления глюкозы, при котором из одной молекулы глюкозы образуются две молекулы пировиноградной кислоты. Гликолиз состоит из цепи последовательных ферментативных реакций и сопровождается запасанием энергии в форме АТФ и НАДH. Гликолиз является универсальным путём катаболизма глюкозы и одним из трёх путей окисления глюкозы, встречающихся в живых клетках. Реакция гликолиза в суммарном виде выглядит следующим образом:
- Глюкоза + 2НАД+ + 2АДФ + 2Pi → 2 пируват + 2НАД*H + 2Н+ + 2АТФ + 2Н2O.
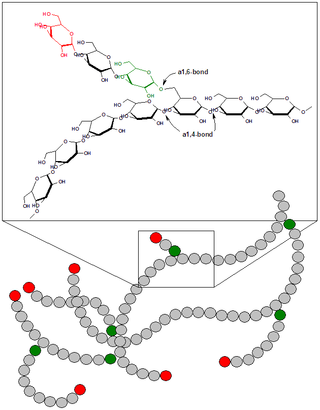
Гликоге́н — гексоза, гомополисахарид состава (C6H10O5)n, образованный остатками глюкозы, соединёнными связями α-1→4 (в местах разветвления — α-1→6). В клетках животных служит основным запасным углеводом и основной формой хранения глюкозы. Откладывается в виде гранул в цитоплазме в клетках многих типов (главным образом в клетках печени и мышц).

Циклический аденозинмонофосфат — органическое соединение, производное АТФ, выполняющее в организме роль вторичного посредника, использующегося для внутриклеточного распространения сигналов некоторых гормонов, которые не могут проходить через клеточную мембрану.

Эдмонд Генри Фи́шер — швейцарский и американский биохимик, лауреат Нобелевской премии по физиологии или медицине 1992 года «За открытия, касающиеся обратимого белкового фосфорилирования как механизма биологической регуляции». Почётный профессор Вашингтонского университета (Сиэтл).

Ге́рти Тере́за Ко́ри, урождённая Ра́дниц — американский биохимик. Член Национальной академии наук США (1948).

Эрл Уилбур Сазерленд-младший — американский физиолог, лауреат Нобелевской премии по физиологии или медицине в 1971 году «за открытия, касающиеся механизмов действия гормонов».
Вторичные посредники, или «вторичные мессенджеры» — это внутриклеточные сигнальные молекулы, высвобождаемые в тех или иных внутриклеточных сигнальных каскадах в ответ на стимуляцию тех или иных рецепторов и вызванную ею активацию первичных эффекторных белков. Вторичные посредники, в свою очередь, приводят к активации вторичных эффекторных белков. Это, в свою очередь, запускает каскад тех или иных физиологических изменений, которые могут быть важны для обеспечения таких важных физиологических процессов, как рост, развитие и дифференцировка клеток, активация деления клетки, транскрипция или, наоборот, угнетение транскрипции тех или иных генов, биосинтез тех или иных белков, выделение ею гормонов, нейромедиаторов или цитокинов соответственно типу клетки, изменение биоэлектрической активности клетки, миграция клеток, обеспечение их выживаемости или, наоборот, индукция апоптоза. Вторичные посредники являются инициирующими элементами во множестве внутриклеточных сигнальных каскадов. Вследствие всего этого вторичные посредники играют очень важную роль в жизни клетки, а грубое нарушение работы любой из систем вторичных посредников оказывает неблагоприятное воздействие на клетку.

Аденилатцикла́за — фермент, который катализирует превращение АТФ в 3',5'-цАМФ с образованием пирофосфата.
Гликогено́лиз — биохимический процесс расщепления гликогена до глюкозо-6-фосфата, и далее, в ряде случаев, до глюкозы. Осуществляется главным образом в печени и скелетных мышцах. Основная задача гликогенолиза — поддержание в организме на постоянном уровне содержания доступных источников энергии: глюкозы в крови и глюкозо-6-фосфата в скелетных мышцах. Регуляция гликогенолиза осуществляется совместно с регуляцией гликогеногенеза, эти два процесса протекают попеременно, в печени — в зависимости от уровня глюкозы в крови, в мышцах — от наличия или отсутстия мышечной активности. Важнейшими гормонами, участвующими в регуляции гликогеногенеза, являются инсулин, глюкагон и адреналин.
Протеинкина́зы — подкласс ферментов киназ (фосфотрансфераз). Протеинкиназы модифицируют другие белки путём фосфорилирования остатков аминокислот, имеющих гидроксильные группы или гетероциклической аминогруппы гистидина.
β2-адренорецепторы — один из подтипов адренорецепторов. Эти рецепторы чувствительны в основном к адреналину, норадреналин действует на них слабо, так как эти рецепторы имеют к нему низкую аффинность.
Углеводный обмен, или метаболизм углеводов в организмах животных и человека. Метаболизм углеводов в организме человека состоит из следующих процессов:
- Расщепление в пищеварительном тракте поступающих с пищей поли- и дисахаридов до моносахаридов, дальнейшее всасывание моносахаридов из кишечника в кровь.
- Синтез и распад гликогена в тканях, прежде всего в печени.
- Гликолиз — распад глюкозы. Первоначально под этим термином обозначали только анаэробное брожение, которое завершается образованием молочной кислоты (лактата) или этанола и углекислого газа. В настоящее время понятие «гликолиз» используется более широко для описания распада глюкозы, проходящего через образование глюкозо-6-фосфата, фруктозо-1,6-дифосфата и пирувата как в отсутствие, так и в присутствии кислорода. В последнем случае употребляется термин «аэробный гликолиз», в отличие от «анаэробного гликолиза», завершающегося образованием молочной кислоты или лактата.
- Анаэробный путь прямого окисления глюкозы или, как его называют, пентозофосфатный путь.
- Взаимопревращение гексоз.
- Анаэробный метаболизм пирувата. Этот процесс выходит за рамки углеводного обмена, однако может рассматриваться как завершающая его стадия: окисление продукта гликолиза — пирувата.
- Глюконеогенез — образование углеводов из неуглеводных продуктов.
Гликогеногене́з — метаболический путь синтеза гликогена из глюкозы, происходящий с расходованием энергии в виде ATP и UTP. Гликогеногенез происходит во всех тканях животных, однако в основном он имеет место в печени и мышцах. Синтез гликогена происходит в период пищеварения (в абсорбтивный период, т. е. 1—2 часа после приёма углеводной пищи.
Аденилатциклазный путь, известный также как цАМФ-зависимый путь — сигнальный каскад, который запускается активацией лигандом-агонистом трансмембранного G-белок-связанного метаботропного рецептора, вследствие чего гетеротримерный G-белок отсоединяет бета-гамма-димер, а альфа-субъединица G-белка активирует либо угнетает, в зависимости от типа альфа-субъединицы, активность аденилатциклазы, что приводит к повышению или снижению концентрации циклического АМФ и к активации либо угнетению вторичных эффекторов, таких, как протеинкиназа A. Это один из основных и самых распространённых способов межклеточной коммуникации и внутриклеточной передачи сигнала в клетках животных, особенно млекопитающих.

Инсулиновый рецептор (IR) — трансмембранный рецептор, который активируется инсулином, IGF-I, IGF-II и принадлежит к большому классу тирозинкиназных рецепторов. Инсулиновый рецептор играет ключевую роль в регуляции гомеостаза глюкозы, функционального процесса, который при дегенеративных условиях может привести к ряду клинических проявлений, в том числе диабету и раку. Биохимически инсулиновый рецептор кодируется одним геном INSR, альтернативный сплайсинг которого в ходе транскрипции производит либо к IR-A-либо к IR-B-изоформам. Последующие посттрансляционные события каждой изоформы приводят к формированию протеолитически расщеплённых α и β-субъединиц, которые при объединении, в конечном счете способны к димеризации с получением дисульфидно связанного трансмембранного инсулинового рецептора массой ≈ 320 кДа.
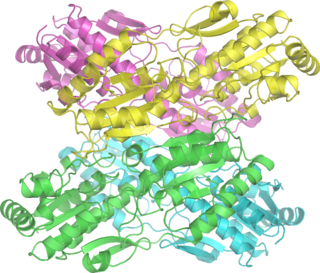
Фосфофруктокиназа-1 — фермент-фосфофруктокиназа, один из наиболее важных регуляторных ферментов. Отвечает за важный этап гликолиза, катализируя перенос фосфатной группы от молекулы АТФ к фруктозо-6-фосфату, в результате чего образуется фруктозо-1,6-бисфосфат и АДФ.

Треониндезаминаза, также треонин аммиак-лиаза или треониндегидратаза — фермент (КФ 4.3.1.19) из семейства дезаминаз (класс лиазы), катализирующий реакцию дезаминирования L-формы треонина (с образованием α-кетобутирата и аммиака). α-Кетобутират затем может быть трансформирован в L-изолейцин. Для функционирования треониндезаминазы необходима активная форма витамина B6 — кофермент пиридоксальфосфат (PLP). Фермент регулируется аллостерически. Треониндезаминаза обнаружена у бактерий, дрожжей и растений, но отсутствует у животных. Именно из-за отсутствия треониндезаминазы изолейцин является незаменимой аминокислотой для человека и других животных.

В биохимии аллостерическая регуляция — это регуляция фермента путем связывания эффекторной молекулы в сайте, отличном от активного сайта фермента.

Протеинфосфатаза 1 принадлежит к классу белковых серин/треониновых фосфатаз. Было обнаружено, что PP1 важна для контроля метаболизма гликогена, сокращения мышц, клеточного развития, нейрональной активности, сплайсинга РНК, митоза, деления клеток, апоптоза, синтеза белка и регуляции мембранных рецепторов и каналов.