
Прокарио́ты, или доя́дерные — одноклеточные живые организмы, не обладающие оформленным клеточным ядром и другими внутренними мембранными органеллами.
Глюконеогене́з — метаболический путь, приводящий к образованию глюкозы из неуглеводных соединений. Наряду с гликогенолизом, этот путь поддерживает в крови уровень глюкозы, необходимый для работы многих тканей и органов, в первую очередь, нервной ткани и эритроцитов. Он служит важным источником глюкозы в условиях недостаточного количества гликогена, например, после длительного голодания или тяжёлой физической работы. Глюконеогенез является обязательной частью цикла Кори, кроме того, этот процесс может быть использован для превращения пирувата, образованного при дезаминировании аминокислот аланина и серина.

Ци́кл трикарбо́новых кисло́т (сокр. ЦТК, цикл Кре́бса, цитра́тный цикл, цикл лимо́нной кислоты́) — центральная часть общего пути катаболизма, циклический биохимический процесс, в ходе которого ацетильные остатки (СН3СО-) окисляются до диоксида углерода (CO2). При этом за один цикл образуется 2 молекулы CO2, 3 НАДН, 1 ФАДH2 и 1 ГТФ (или АТФ). Электроны, находящиеся на НАДН и ФАДH2, в дальнейшем переносятся на дыхательную цепь, где в ходе реакций окислительного фосфорилирования образуется АТФ.

Фотоси́нтез — сложный химический процесс преобразования энергии видимого света в энергию химических связей органических веществ при участии фотосинтетических пигментов.
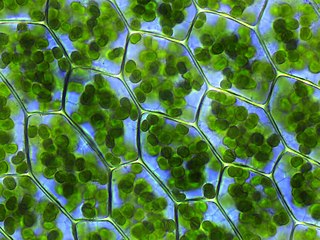
Хлоропла́сты — зелёные пластиды, которые встречаются в клетках фотосинтезирующих эукариот. С их помощью происходит фотосинтез. Хлоропласты содержат хлорофилл. У зелёных растений являются двумембранными органеллами. Под двойной мембраной имеются тилакоиды. Тилакоиды высших растений группируются в граны, которые представляют собой стопки сплюснутых и тесно прижатых друг к другу тилакоидов, имеющих форму дисков. Соединяются граны с помощью ламелл. Пространство между оболочкой хлоропласта и тилакоидами называется стромой. В строме содержатся хлоропластные молекулы РНК, пластидная ДНК, рибосомы, крахмальные зёрна, а также ферменты цикла Кальвина.

Малатдегидрогеназа (Дегидрогеназа яблочной кислоты, англ. MDH, EC 1.1.1.37 — фермент, катализирующий окисление S-малата до оксалоацетата.
Гетероцисты — дифференцированные клетки нитчатых цианобактерий, осуществляющие азотфиксацию. При недостатке соединений азота в среде они появляются регулярно вдоль трихомы из вегетативных клеток и акинет. Цианобактерии — фототрофы, осуществляющие оксигенный фотосинтез, однако кислород, атмосферный и выделяемый при фотосинтезе, ингибирует фермент нитрогеназу, необходимую для азотфиксации, поэтому у нитчатых цианобактерий в процессе эволюции возникли специализированные клетки для азотфиксации.

Динофлагелля́ты, или динофи́товые во́доросли, или динофи́ты, или перидине́и, или па́нцирные жгутиконо́сцы — крупная группа протистов из надтипа альвеолят (Alveolata), которой традиционно присваивают ранг типа. Известно около 4000 ископаемых и более 2500 современных видов, из которых 90 % обитает в морях, остальные — в пресных водах. Около половины представителей — свободноживущие фотосинтезирующие организмы, однако известны и бесцветные гетеротрофные формы, и паразитические динофлагелляты. Некоторые виды являются симбионтами коралловых полипов и двустворчатых моллюсков.

Циане́ллы — пластиды глаукофитовых водорослей. Окрашены в сине-зелёный цвет, так как зелёный хлорофилл a маскируется пигментами фикоцианином и аллофикоцианином, расположенными на поверхности тилакоидов. Особое название используется для них потому, что они обладают уникальным признаком: имеют тонкую клеточную стенку из пептидогликана (муреина) между наружной и внутренней мембраной. По большинству других признаков цианеллы напоминают типичные цианобактерии, хотя содержат сильно редуцированный геном.
Глаукофи́товые во́доросли, или глаукофи́ты, или глаукоцистофи́ты — небольшой древний отдел одноклеточных водорослей, включающий восемь родов и 21 вид. Глаукофиты особенно интересны своими уникальными хлоропластами (цианеллами), довольно примитивно устроенными по сравнению с хлоропластами других водорослей: они содержат слой муреина между мембранами и характеризуются множеством других свойств, присущих цианобактериям. Согласно современным представлениям, глаукофитовые водоросли обособились в отдельную группу у самого основания Archaeplastida.

Сирогем — гемоподобная простетическая группа нитрит- и сульфитредуктаз, катализирующих шестиэлектронное восстановление серы сульфита или азота нитрита до сульфида и аммиака.

Протохлорофиллид, или моновинил протохлорофиллид, это непосредственный предшественник хлорофилла a с отсутствующим фитольным хвостом. В отличие от хлорофилла, протохлорофиллид имеет сильную флюоресценцию; мутанты, накапливающие его в своих тканях, светятся красным, если облучить их синим светом. У цветковых растений, реакция превращения протохлорофиллида в хлорофилл является светозависимой, и такие растения становятся белыми (хлорозными) если выращивать их в темноте. В отличие от них голосеменные, водоросли, и фотосинтезирующие бактерии используют другой, не зависящий от света фермент, и вырастают зелёными даже в темноте.
Электрон-транспортная цепь фотосинтеза — последовательность переносчиков электронов, расположенных на белках фотосинтетических мембран и осуществляющих фотоиндуцированный транспорт электронов, сопряжённый с трансмембранным переносом протонов против электрохимического градиента.
Ферредокси́ны — группа небольших растворимых белков, содержащих железосерные кластеры и являющихся подвижными переносчиками электронов в ряде метаболических процессов. Обычно они переносят один или два электрона за счёт изменения окисленности атомов железа.

Фотосистема представляет собой функциональную и структурную единицу белковых комплексов, которые осуществляют первичные фотохимические реакции фотосинтеза: поглощение света, преобразование энергии и перенос электронов. Фотосистемы находятся в тилакоидной мембраны растений, водорослей и цианобактерий, либо в цитоплазматической мембране фотосинтезирующих бактерий. В общем случае все фотосистемы подразделяют на два типа — подобные фотосистеме II и подобные фотосистеме I.

Седогептулозобисфосфатаза представляет собой фермент, который катализирует удаление фосфатной группы от седогептулозо-1,7-бисфосфата с образованием седогептулозо-7-фосфата. По своей активности — фосфатаза или, говоря более обобщенно, гидролаза. Участвует в цикле Кальвина.
Ферредоксин-НАДФ+-редуктаза, сокращенно ФНР, фермент из класса оксидоредуктаз, катализирующий реакцию восстановления НАДФ+, используя в качестве донора электронов ферредоксин.
Транслоказы — отдельный класс ферментов, катализирующих перенос ионов или молекул через мембраны или их разделение в мембранах. Этот класс ферментов сформировался из ранее принадлежащих другим классам ферментов.

Аденозин-5′-фосфосульфат (АФС, APS) — производное аденозинмонофосфата, сульфатированное по 5′-концу. Промежуточный продукт метаболизма сульфатов (SO42−) и сульфитов (SO32−).
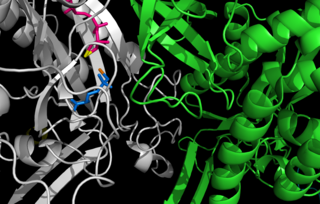
Глутаминсинтетаза, также L-глутамат-аммиак лигаза (сокр. ГС) — фермент (КФ 6.3.1.2) из класса синтетаз (лигаз), принимает участие в обезвреживании свободного аммиака в тканях. Данный фермент катализирует в присутствии двухвалентных ионов (Mg2+ или Mn2+) реакцию образования аминокислоты L-глутамина, посредством присоединения к L-глутамату молекул аммиака NH3 (in vivo свободный аммиак ионизирован и представляет собой катион аммония — NH4+), с использованием энергии гидролиза макроэргических связей АТФ. Реакция имеет следующий вид: