
Фотоси́нтез — сложный химический процесс преобразования энергии видимого света в энергию химических связей органических веществ при участии фотосинтетических пигментов.

В биологии активный центр — это область фермента, где молекулы субстрата связываются и подвергаются химической реакции. Активный центр состоит из аминокислотных остатков, которые образуют временные связи с субстратом, и остатков, которые катализируют реакцию этого субстрата. Хотя активный центр занимает только ~ 10-20 % от объёма фермента он является наиболее важной частью, поскольку он непосредственно катализирует химическую реакцию. Обычно активный центр состоит из трех-четырех аминокислот, в то время как другие аминокислоты в белке необходимы для поддержания его третичной структуры.

Малатдегидрогеназа (Дегидрогеназа яблочной кислоты, англ. MDH, EC 1.1.1.37 — фермент, катализирующий окисление S-малата до оксалоацетата.

Никотинамидадениндинуклеотидфосфа́т — широко распространённый в природе кофермент некоторых дегидрогеназ — ферментов, катализирующих окислительно-восстановительные реакции в живых клетках. НАДФ принимает на себя водород и электроны окисляемого соединения и передаёт их на другие вещества. В хлоропластах растительных клеток НАДФ восстанавливается при световых реакциях фотосинтеза и затем обеспечивает водородом синтез углеводов при темновых реакциях. НАДФ, — кофермент, отличающийся от НАД содержанием ещё одного остатка фосфорной кислоты, присоединённого к гидроксилу одного из остатков D-рибозы, обнаружен во всех типах клеток.

Клеточное, или тканевое дыхание — совокупность биохимических реакций, протекающих в клетках живых организмов, в ходе которых происходит окисление углеводов, липидов и аминокислот до углекислого газа и воды, а также образование энергии. Высвобожденная энергия запасается в химических связях макроэргических соединений и может быть использована по мере необходимости. Входит в группу процессов катаболизма. О физиологических процессах транспортировки к клеткам многоклеточных организмов кислорода и удалению от них углекислого газа см. статью Дыхание.
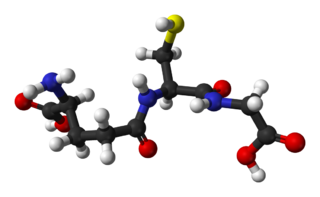
Глутатион — это трипептид γ-глутамилцистеинилглицин. Глутатион содержит необычную пептидную связь между аминогруппой цистеина и карбоксильной группой боковой цепи глутамата. Значение глутатиона в клетке определяется его антиоксидантными свойствами. Фактически глутатион не только защищает клетку от токсичных свободных радикалов, но и в целом определяет окислительно-восстановительные характеристики внутриклеточной среды.

Гемолитическая анемия — групповое название редких заболеваний, общим признаком которых является усиленное разрушение эритроцитов, обусловливающее, с одной стороны, анемию и повышенное образование продуктов распада эритроцитов, с другой стороны — реактивно усиленный эритропоэз.

Флавины — группа органических соединений - производных диметилизоаллоксазина ( ), замещенных обычно по положению 10.

Глутатионпероксидазы (ГП, англ. Glutathione peroxidase, PDB 1GP1, — семейство ферментов, защищающих организм от окислительного повреждения. Глутатионпероксидазы катализируют восстановление перекисей липидов в соответствующие спирты и восстановление пероксида водорода до воды. Известно несколько генов, кодирующих разные формы глутатионпероксидаз, отличающиеся по локализации в организме. Структурно ферменты данного семейства представляют собой селеносодержащие тетрамерные гликопротеины.

НАДФH-оксидаза, или NADPH-оксидаза (NOX) — клеточный мембрано-связанный мультимолекулярный ферментный комплекс, локализующийся на плазматической мембране и в некоторых органеллах. Особенно обогащены этим ферментом фагоцитарные клетки, такие как макрофаги. Эти оксидазы участвуют в клеточной противомикробной защитной системе, а также в клеточной пролиферации, дифференцировке и регуляции экспрессии генов. Существует целая группа НАДФH-оксидаз, которые различаются по составу субъединиц, клеточной специфичности, регуляции и другим параметрам.
Глюкозо-6-фосфатдегидрогеназа — цитозольный фермент, входящий в пентозофосфатный путь, метаболический путь, обеспечивающий образование клеточного НАДФ-H из НАДФ+. НАДФ-H необходим для поддержания уровня восстановленного глутатиона в клетке, синтеза жирных кислот и изопреноидов. У человека наследственное нарушение активности Г6ФД, или недостаточность глюкозо-6-фосфатдегидрогеназы, приводит к гемолитической анемии.

Пентозофосфа́тный путь — альтернативный путь окисления глюкозы, включает в себя окислительный и неокислительный этапы.
Углеводный обмен, или метаболизм углеводов в организмах животных и человека. Метаболизм углеводов в организме человека состоит из следующих процессов:
- Расщепление в пищеварительном тракте поступающих с пищей поли- и дисахаридов до моносахаридов, дальнейшее всасывание моносахаридов из кишечника в кровь.
- Синтез и распад гликогена в тканях, прежде всего в печени.
- Гликолиз — распад глюкозы. Первоначально под этим термином обозначали только анаэробное брожение, которое завершается образованием молочной кислоты (лактата) или этанола и углекислого газа. В настоящее время понятие «гликолиз» используется более широко для описания распада глюкозы, проходящего через образование глюкозо-6-фосфата, фруктозо-1,6-дифосфата и пирувата как в отсутствие, так и в присутствии кислорода. В последнем случае употребляется термин «аэробный гликолиз», в отличие от «анаэробного гликолиза», завершающегося образованием молочной кислоты или лактата.
- Анаэробный путь прямого окисления глюкозы или, как его называют, пентозофосфатный путь.
- Взаимопревращение гексоз.
- Анаэробный метаболизм пирувата. Этот процесс выходит за рамки углеводного обмена, однако может рассматриваться как завершающая его стадия: окисление продукта гликолиза — пирувата.
- Глюконеогенез — образование углеводов из неуглеводных продуктов.

1,3-Бисфосфоглицериновая кислота — смешанный ангидрид фосфорной кислоты и карбоксильной группы. Промежуточный продукт в реакциях гликолиза, глюконеогенеза, а также цикла Кальвина — наиболее распространённого в биосфере пути фиксации углерода при фотосинтезе и хемосинтезе.

Фотосисте́ма I, или пластоциани́н-ферредокси́н-оксидоредукта́за — второй функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он принимает электрон от пластоцианина и, поглощая световую энергию, формирует сильный восстановитель П700, способный через цепь переносчиков электронов осуществить восстановление НАДФ+. Таким образом, при участии ФСI синтезируется источник электронов (НАДФН) для последующих реакций восстановления углерода в хлоропластах в цикле Кальвина. Кроме того, ФСI может осуществлять циклический транспорт электронов, сопряжённый с синтезом АТФ, обеспечивая дополнительный синтез АТФ в хлоропластах.

Реакционный центр — комплекс белков, пигментов и других кофакторов, взаимодействие которых обеспечивает реакцию превращения энергии света в химическую при фотосинтезе. Реакционный центр получает энергию или через непосредственное возбуждение одной из своих молекул или через перенос энергии от светособирающих комплексов, что даёт начало цепочке химических реакций, происходящей на связанных белками кофакторах. Эти кофакторы — светопоглощающие молекулы такие как хлорофилл, феофитин и хиноны. Энергия фотона используется для поднятия электрона на более высокий энергетический уровень. Запасённая таким образом свободная энергия идёт на восстановление цепочки акцепторов электрона с более высоким редокс-потенциалом.

Фотосисте́ма II (втора́я фотосисте́ма, фотосисте́ма два, ФСII), или H2O-пластохиноноксидоредуктаза — первый функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он расположен в мембранах тилакоидов всех растений, водорослей и цианобактерий. Поглощая энергию света в ходе первичных фотохимических реакций, он формирует сильный окислитель — димер хлорофилла a (П680+), который через цепь окислительно-восстановительных реакций способен вызвать окисление воды.

Цитохро́м-b6f-ко́мплекс, или пластохинолпластоцианинредукта́за — мультибелковый комплекс, который осуществляет окисление пластохинолов и восстановление белка пластоцианина, обеспечивая, таким образом, транспорт электронов между реакционными центрами фотосистемы I (ФСI) и фотосистемы II (ФСII). Он восстанавливает маленький водорастворимый белок пластоцианин, который переносит электрон к ФСII. Аналогичную реакцию катализирует цитохром-bc1-комплекс электрон-транспортной цепи митохондрий. Цитохром-b6f-комплекс присутствует в тилакоидной мембране хлоропластов растений, водорослей и цианобактерий. Он функционально объединяет две фотосистемы в единую цепь переноса электронов от воды к НАДФ+, то есть является участником нециклического потока электронов. Кроме того, цитохромный комплекс вовлечён в циклический транспорт электронов, осуществляемый фотосистемой I.
Реакция Хилла, или хлоропластная реакция, была открыта в 1937 году биохимиком Робертом Хиллом из Кембриджского университета. Реакция представляет собой светозависимый перенос электронов от воды на реактив Хилла против градиента химического потенциала. В лабораторной практике эта реакция служит для определения фотохимической активности хлоропластов, обусловленной фотоокислением воды. Реакция Хилла продемонстрировала, что образование кислорода и синтез сахаров из углекислого газа — это два разных, независимых процесса. Эти данные, полученные Хиллом, легли в основу современного понимания фотосинтеза.
Ферредоксин-НАДФ+-редуктаза, сокращенно ФНР, фермент из класса оксидоредуктаз, катализирующий реакцию восстановления НАДФ+, используя в качестве донора электронов ферредоксин.
