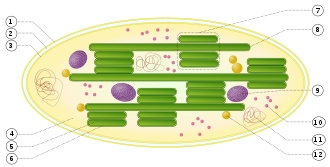
Органе́ллы, раннее называвшиеся также органо́идами, — постоянные компоненты клетки. Располагаются во внутренней части клетки — цитоплазме, в которой, наряду с органеллами, могут находиться различные включения.

Пласти́ды — полуавтономные органеллы высших растений, водорослей и некоторых фотосинтезирующих простейших. Пластиды имеют от двух до четырёх мембран, собственный геном и белоксинтезирующий аппарат.

Фотоси́нтез — сложный химический процесс преобразования энергии видимого света в энергию химических связей органических веществ при участии фотосинтетических пигментов.
Хлоропла́сты — зелёные пластиды, которые встречаются в клетках фотосинтезирующих эукариот. С их помощью происходит фотосинтез. Хлоропласты содержат хлорофилл. У зелёных растений являются двумембранными органеллами. Под двойной мембраной имеются тилакоиды. Тилакоиды высших растений группируются в граны, которые представляют собой стопки сплюснутых и тесно прижатых друг к другу тилакоидов, имеющих форму дисков. Соединяются граны с помощью ламелл. Пространство между оболочкой хлоропласта и тилакоидами называется стромой. В строме содержатся хлоропластные молекулы РНК, пластидная ДНК, рибосомы, крахмальные зёрна, а также ферменты цикла Кальвина.

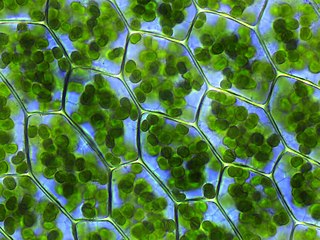
Зелёные во́доросли — группа низших растений. В современной систематике эта группа имеет ранг отдела, включающего одноклеточные и колониальные планктонные водоросли, одноклеточные и многоклеточные формы бентосных водорослей. Здесь встречаются все морфологические типы слоевища, кроме ризоподиальных одноклеточных и крупных многоклеточных форм со сложным строением. Многие нитчатые зелёные водоросли крепятся к субстрату только на ранних стадиях развития, затем они становятся свободноживущими, формируя маты или шары.
Лейкопла́сты — бесцветные сферические пластиды в клетках растений. Основная функция лейкопластов — накопление питательных веществ.

Кра́сные во́доросли, или багря́нки — отдел водорослей, обитатели прежде всего морских водоёмов, пресноводных представителей известно немного. Обычно это довольно крупные растения, но встречаются и микроскопические. Среди красных водорослей имеются одноклеточные, нитчатые и псевдопаренхимные формы; истинно паренхимные формы отсутствуют. Ископаемые остатки свидетельствуют, что это очень древняя группа растений. Красные водоросли неоднократно переходили к паразитированию на других красных водорослях: как близкородственных (адельфопаразитизм), так и более удалённых.

Стро́ма — основа (остов) паренхиматозного органа, состоящая из ретикулярной соединительной ткани (интерстиция), представляет собой мелкопетлистую трёхмерную сеть, в петлях которой расположены паренхима органа, имеются способные к размножению клетки, а также волокнистые структуры, обусловливающие её опорное значение. В строме проходят кровеносные и лимфатические сосуды; элементы стромы играют и защитную роль, так как способны к фагоцитозу. Из клеток стромы кроветворных органов развиваются красные и белые кровяные тельца, где строма выполняет функцию микроокружения для развивающихся форменных элементов крови.
Тилакоиды — ограниченные мембраной компартменты внутри хлоропластов и цианобактерий. В тилакоидах происходят светозависимые реакции фотосинтеза. Слово «тилакоид» происходит от греческого слова θύλακος, означающего «мешочек». Тилакоиды состоят из мембраны, окружающей просвет тилакоида. Тилакоиды хлоропластов часто имеют структуру, напоминающую стопку дисков. Эти стопки называют гранами. Граны соединены межграновыми или строматическими тилакоидами (ламеллами) в единое функциональное пространство.
У термина компартментализация существует другое значение, см. Раздельное мышление.
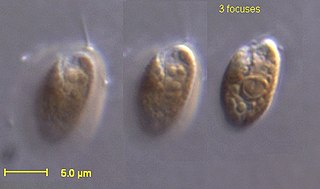
Криптофи́товые во́доросли, или криптомона́ды, или криптофи́ты , — группа одноклеточных эукариотических фотосинтезирующих организмов, включающая около 165 видов, которой традиционно присваивают ранг типа. Почти все криптофитовые имеют монадную форму с дорсовентральным строением, несут два неравных жгутика. Покровы клетки представлены перипластом, имеются стрекательные структуры (эжектосомы). Хлоропласты окружены четырьмя мембранами и содержат редуцированное ядро — нуклеоморф. Основные фотосинтетические пигменты — хлорофиллы a, c2, а также каротиноиды и фикобилины. Митоз обычно открытый, без центриолей, размножение в основном — вегетативное.

Циане́ллы — пластиды глаукофитовых водорослей. Окрашены в сине-зелёный цвет, так как зелёный хлорофилл a маскируется пигментами фикоцианином и аллофикоцианином, расположенными на поверхности тилакоидов. Особое название используется для них потому, что они обладают уникальным признаком: имеют тонкую клеточную стенку из пептидогликана (муреина) между наружной и внутренней мембраной. По большинству других признаков цианеллы напоминают типичные цианобактерии, хотя содержат сильно редуцированный геном.
Глаукофи́товые во́доросли, или глаукофи́ты, или глаукоцистофи́ты — небольшой древний отдел одноклеточных водорослей, включающий восемь родов и 21 вид. Глаукофиты особенно интересны своими уникальными хлоропластами (цианеллами), довольно примитивно устроенными по сравнению с хлоропластами других водорослей: они содержат слой муреина между мембранами и характеризуются множеством других свойств, присущих цианобактериям. Согласно современным представлениям, глаукофитовые водоросли обособились в отдельную группу у самого основания Archaeplastida.

Расти́тельные кле́тки — эукариотические клетки, однако несколькими своими свойствами они отличаются от клеток остальных эукариот. К их отличительным чертам относят:
- Крупная центральная вакуоль, пространство, заполненное клеточным соком и ограниченное мембраной — тонопластом. Вакуоль играет ключевую роль в поддержании клеточного тургора, контролирует перемещение молекул из цитозоля в выделения клетки, хранит полезные вещества и расщепляет отслужившие старые белки и органеллы.
- Есть клеточная стенка, состоящая главным образом из целлюлозы, а также гемицеллюлозы, пектина и во многих случаях лигнина. Она образуется протопластом поверх клеточной мембраны. Она отличается от клеточной стенки грибов, состоящей из хитина, и бактерий, построенной из пептидогликана (муреина).
- Специализированные пути связи между клетками — плазмодесмы, цитоплазматические мостики: цитоплазма и эндоплазматический ретикулум (ЭПР) соседних клеток сообщаются через поры в клеточных стенках.
- Пластиды, из которых наиболее важны хлоропласты. Хлоропласты содержат хлорофилл, зелёный пигмент, поглощающий солнечный свет. В них осуществляется фотосинтез, в ходе которого клетка синтезирует органические вещества из неорганических. Другими пластидами являются лейкопласты: амилопласты, запасающие крахмал, элайопласты, хранящие жиры и др., а также хромопласты, специализирующиеся на синтезе и хранении пигментов. Как и митохондрии, чей геном у растений содержит 37 генов, пластиды имеют собственные геномы (пластомы), состоящие из около 100—120 уникальных генов. Как предполагается, пластиды и митохондрии возникли как прокариотические эндосимбионты, поселившиеся в эукариотических клетках.
- Деление клеток (митоз) наземных растений и некоторых водорослей, особенно харовых (Charophyta) и порядка Trentepohliales характеризуется наличием дополнительной стадии — препрофазы. Помимо этого цитокинез у них осуществляется при помощи фрагмопласта — «формы» для строящейся клеточной пластинки.
- Мужские половые клетки мхов и папоротниковидных имеют жгутик, схожий со жгутиком сперматозоидов животных, но у семенных растений — голосеменных и цветковых — они лишены жгутика и называются спермиями.
- Из присущих животной клетке органелл у растительной отсутствуют только центриоли.
Электрон-транспортная цепь фотосинтеза — последовательность переносчиков электронов, расположенных на белках фотосинтетических мембран и осуществляющих фотоиндуцированный транспорт электронов, сопряжённый с трансмембранным переносом протонов против электрохимического градиента.

Фотосисте́ма I, или пластоциани́н-ферредокси́н-оксидоредукта́за — второй функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он принимает электрон от пластоцианина и, поглощая световую энергию, формирует сильный восстановитель П700, способный через цепь переносчиков электронов осуществить восстановление НАДФ+. Таким образом, при участии ФСI синтезируется источник электронов (НАДФН) для последующих реакций восстановления углерода в хлоропластах в цикле Кальвина. Кроме того, ФСI может осуществлять циклический транспорт электронов, сопряжённый с синтезом АТФ, обеспечивая дополнительный синтез АТФ в хлоропластах.
Танносомы — органеллы, участвующие в транспорте танинов из хлоропластов в вакуоли. Обнаружены в клетках сосудистых растений.

Цитохро́м-b6f-ко́мплекс, или пластохинолпластоцианинредукта́за — мультибелковый комплекс, который осуществляет окисление пластохинолов и восстановление белка пластоцианина, обеспечивая, таким образом, транспорт электронов между реакционными центрами фотосистемы I (ФСI) и фотосистемы II (ФСII). Он восстанавливает маленький водорастворимый белок пластоцианин, который переносит электрон к ФСII. Аналогичную реакцию катализирует цитохром-bc1-комплекс электрон-транспортной цепи митохондрий. Цитохром-b6f-комплекс присутствует в тилакоидной мембране хлоропластов растений, водорослей и цианобактерий. Он функционально объединяет две фотосистемы в единую цепь переноса электронов от воды к НАДФ+, то есть является участником нециклического потока электронов. Кроме того, цитохромный комплекс вовлечён в циклический транспорт электронов, осуществляемый фотосистемой I.

Хлородыхание — процесс в хлоропластах растений, который, как предполагается, включает цепь переноса электронов в мембране тилакоида. Перенос протона осуществляется двумя ферментами: НАД(Ф)Н-дегидрогеназным комплексом и альтернативной оксидазой, подобной митохондриальной альтернативной оксидазе. Оба фермента способны регулировать окислительно-восстановительное состояние пула пластохинонов в хлоропласте. Этот процесс может происходить для того, чтобы сбалансировать уровни АТФ в хлоропласте ночью, когда циклическая передача электрона в Фотосистеме I не происходит.