Двусторонне-симметричные
| Двусторонне-симметричные | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
Домен: Эукариоты Царство: Животные Подцарство: Эуметазои Без ранга: Двусторонне-симметричные | |||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Bilateria Hatschek, 1888 | |||||||||||||||||||||||||||||||||||||||||
| Дочерние таксоны | |||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
| Геохронология появился 560 млн лет
◄ Наше время◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание◄ Массовоепермское вымирание ◄ Девонское вымирание◄ Ордовикско-силурийское вымирание◄ Кембрийский взрыв | |||||||||||||||||||||||||||||||||||||||||
Двусторо́нне-симметри́чные[1], или билатера́льные[1], или билате́рии[2] (лат. Bilateria), — таксон, включающий настоящих многоклеточных животных, характеризующихся наличием передне-задней оси тела, систем органов и чётко определённой или истинной эндодермальной мезодермой[3].
Билатеральные — огромная группа, к которой относится почти 99 % описанных видов животных. Известны всего четыре современных типа животных, не относящихся к билатериям: губки (8000 видов), стрекающие (11000 видов), гребневики (150 видов) и пластинчатые (всего 3 вида). Все остальные животные (28 типов, около 1,6 млн описанных видов) относятся к двусторонне-симметричным[4].
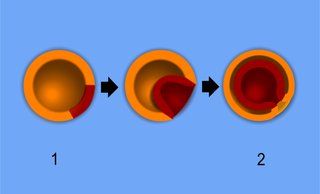
Характерная особенность строения большинства Bilateria — двусторонняя симметрия, при которой тело организма можно разделить на левую и правую половины, зеркально повторяющие друг друга. На основе генетических и биохимических исследований эта группа считается монофилетической, то есть все двусторонне-симметричные организмы имели общего эволюционного предка. В то же время у представителей некоторых групп Bilateria двусторонняя симметрия может нарушаться и даже полностью утрачиваться[5], что наблюдается, например, в некоторой асимметрии внешнего и, особенно, внутреннего строения человека (в неравном развитии мускулатуры, положении сердца, печени и многих других внутренних органов). Один из наиболее известных случаев утраты двусторонней симметрии — поздние личиночные и взрослые стадии иглокожих, расположение систем органов которых глубоко асимметрично или даже приобретает характер радиальной симметрии[5].
Другие особенности билатерий:
- Сложная нервная система, как правило объединённая в цепочку или тяж. У животных, не относящихся к билатеральным, нервной системы или нет вообще (губки и пластинчатые), или она устроена гораздо примитивнее, так называемая диффузная нервная система (стрекающие и гребневики).
- Сквозной кишечник в виде трубки, с однонаправленным движением пищи от рта к анальному отверстию. У небилатерий если кишечник вообще есть, то однонаправленного потока пищи в нем никогда не бывает, непереваренные остатки пищи удаляются через рот.
- Три зародышевых листка: эктодерма, энтодерма и мезодерма. У животных, не относящихся к билатериям, зародышевые листки либо вообще не выделяются (губки, пластинчатые), либо их только два — эктодерма и эндодерма. Поэтому иногда билатерий относят к трехслойным животным, а всех остальных — к двухслойным. Из мезодермы развиваются мышцы, поэтому билатерии способны к гораздо более активным и разнообразным движениям[4].
Механизм формирования двусторонней симметрии
Хотя конкретные механизмы могут существенно отличаться друг от друга, общий принцип формирования двусторонней симметрии в процессе онтогенеза у различных Bilateria не претерпел принципиальных изменений со времени жизни их последнего общего предка, то есть на протяжении уже около 700 млн лет. Как и все процессы дифференцировки клеток, он происходит за счёт экспрессии гомеозисных генов, в результате которой в растущем организме возникает определённый градиент концентрации гомеозисных белков, играющих роль морфогенов. В зависимости от концентрации белков-морфогенов в клетках происходит «включение» или «выключение» определённых генов (набор генов одинаков во всех клетках организма, кроме половых, однако далеко не все гены «включены» в данный момент, что и создаёт возможность для существования в рамках одного организма различных типов клеток), «запускающее» ту или иную «программу» их развития и, соответственно, формирование тех или иных структур. Разница в концентрации белков-морфогенов обуславливает формирование переднего и заднего концов, правой и левой, дорсальной и вентральной сторон тела, а у артропод — ещё и разделение его на сегменты.
Данный процесс хорошо изучен на примере формирования передне-задней оси тела у плодовой мушки-дрозофилы (см. статью Эмбриогенез дрозофилы). Формирование переднего отдела тела этого животного (головы и торакса) происходит благодаря возникающей ещё в организме матери до оплодотворения повышенной концентрации на переднем конце яйца белка-морфогена bicoid, синтезируемого одноимённым геном. Этот белок подавляет синтез белка caudal, активирующего программу развития задней части тела, и одновременно вызывает экспрессию гена hunchback, связанного с сегментацией тела. У мутантов с «выключенным» геном bicoid на месте передних отделов тела формируется вторая задняя часть. При введении же продуктов гена bicoid в среднюю область яиц развивается личинка с передними структурами в центральной области и с задними структурами на полюсах яйца[6].
Система гомеозисных генов и соответствующих им белков у Bilateria исключительно консервативна. Несмотря на то, что общий предок птиц и насекомых жил примерно 670 миллионов лет назад[7], гомеозисные гены курицы сходны с аналогичными генами мухи-дрозофилы до такой степени, что являются функционально взаимозаменяемыми: развитие эмбриона мухи с соответствующими гомеозисными генами курицы, внедрёнными методами генной инженерии, протекает нормально[8].
Эволюция
Существует две полярных точки зрения на время появления билатерий. Первая основана на данных палеонтологии, вторая — на данных молекулярных часов. Согласно первой, появление двусторонне-симметричных животных произошло незадолго до начала кембрия[9] До 2020 года не было никаких убедительных свидетельств существования билатерий ранее 560 млн лет назад[10]. В 2020 году был описан древнейший двусторонне-симметричный организм, Ikaria wariootia, чьи окаменелые остатки найдены в Южной Австралии, в породах, которые датируют 555 миллионами лет[11]. С другой стороны, по данным молекулярных часов, билатерии появились значительно раньше — 600—650 млн лет назад[12][примечание 1]. Отсутствие ископаемых свидетельств существования билатерий сторонники «молекулярной» версии объясняют тем, что ранние представители этой группы были микроскопическими и/или мягкотелыми существами, вероятность фоссилизации которых чрезвычайно низка[13].
Вполне вероятно, что именно развитие разнообразия билатерий в конце эдиакара — начале кембрия привело к кембрийскому взрыву — очень быстрому по эволюционным меркам (несколько десятков миллионов лет) появлению представителей почти всех современных типов животных в раннем кембрии (около 540—510 млн лет назад). Вероятно, первые билатерии имели центральную нервную систему, сквозную кишку и расположенные вокруг неё мышцы (возможно, сегментированные), что делает удобным как плавание, так и ползание или рытьё. Появление роющих и активно плавающих жизненных форм привело к развитию принципиально новых экологических ниш, значительному ускорению транспорта органических веществ и кислорода на большие глубины, что привело к ещё большему развитию биоразнообразия по механизму положительной обратной связи[4].
Билатерии вскоре после своего появления разделились на два эволюционных ствола — Protostomia (первичноротые) и Deuterostomia (вторичноротые). Их основным отличительным признаком считается способ формирования рта: у первичноротых рот развивается из бластопора (эмбрионального рта — отверстия первичной кишки на стадии гаструлы), а у вторичноротых рот не связан с бластопором[2]. Первичноротые, в свою очередь, разделились на две крупные эволюционные линии — спиральные и линяющие[14].
Комментарии
- ↑ Подразумевается, что появление животных, обладающих всеми признаками билатерий, произошло после отделения ветви книдарий, но до разделения билатерий на главные подгруппы (либо Nephrozoa vs Xenacoelomorpha, либо первичноротые vs вторичноротые)
Примечания
- ↑ 1 2 Эуметазои // Биологический энциклопедицеский словарь / Под ред. М. С. Гилярова. — М.: Советская энциклопедия, 1986. — С. 743. — 831 с. Архивировано 26 августа 2014 года.
- ↑ 1 2 Марков А. Эмбриональное развитие брахиопод проливает свет на природу первично- и вторичноротости. Элементы (26 декабря 2016). Дата обращения: 30 декабря 2016. Архивировано 29 декабря 2016 года.
- ↑ Kotpal R. L. Modern Text Book of Zoology: Invertebrates. — Rastogi Publications, 2012. — P. 195. — 908 p. — ISBN 9788171339037. Архивировано 12 марта 2018 года.
- ↑ 1 2 3 Дупликация гомеобоксных генов могла быть одной из причин кембрийского взрыва • Сергей Ястребов • Новости науки на «Элементах» • Эволюция, Палеонтология, Генетика Архивная копия от 30 октября 2018 на Wayback Machine.
- ↑ 1 2 Вестхайде В., Ригер Р. Зоология беспозвоночных. = Spezielle Zoology. Teil 1: Einzeller und Wirbellose Tiere / пер. с нем. О. Н. Бёллинг, С. М. Ляпкова, А. В. Михеев, О. Г. Манылов, А. А. Оскольский, А. В. Филиппова, А. В. Чесунов; под ред. А. В. Чесунова. — М.: Товарищество научных изданий КМК, 2008.
- ↑ Driever W., Siegel V., Nüsslein-Volhard C. Autonomous determination of anterior structures in the early Drosophila embryo by the bicoid morphogen (англ.) // Development. — 1990. — Vol. 109, no. 4. — P. 811—820. — doi:10.1242/dev.109.4.811. — PMID 2226200.
- ↑ Ayala, F.J.; A. Rzhetskydagger. Origin of the metazoan phyla: Molecular clocks confirm paleontological estimates (англ.) // Proceedings of the National Academy of Sciences. — 1998. — 20 January (vol. 95, no. 2). — P. 606—611. — doi:10.1073/pnas.95.2.606. — PMID 9435239. Архивировано 5 сентября 2022 года.
- ↑ Lutz, B.; H.C. Lu, G. Eichele, D. Miller, and T.C. Kaufman. Rescue of Drosophila labial null mutant by the chicken ortholog Hoxb-1 demonstrates that the function of Hox genes is phylogenetically conserved (англ.) // Genes & Development. — 1996. — Vol. 10. — P. 176—184. — doi:10.1101/gad.10.2.176. — PMID 8566751. Архивировано 5 сентября 2022 года.
- ↑ Budd&Jenssen, 2017, fig.6.
- ↑ Cunningham et.al., 2017.
- ↑ Evans S. D., Hughes I. V., Gehling J. G., Droser M. L. Discovery of the oldest bilaterian from the Ediacaran of South Australia (англ.) // Proceedings of the National Academy of Sciences. — 2020. — 23 March (vol. 117, no. 14). — doi:10.1073/pnas.2001045117. — PMID 32205432. Архивировано 5 сентября 2022 года.
- ↑ Reis et.al., 2015.
- ↑ Breandán Anraoi MacGabhann, James D. Schiffbauer, James W.Hagadorn, Peter Van Roy, Edward P.Lynch, Liam Morrison, John Murrayah. Resolution of the earliest metazoan record: Differential taphonomy of Ediacaran and Paleozoic fossil molds and casts (англ.) // Palaeogeography, Palaeoclimatology, Palaeoecology[англ.] : journal. — 2019. — Vol. 513. — P. 146—165.
- ↑ Морские стрелки оказались родственниками коловраток • Сергей Ястребов • Новости науки на «Элементах» • Систематика, Эволюция. Дата обращения: 6 марта 2019. Архивировано 13 августа 2019 года.
Литература
- Budd, Graham. The origin of the animals and a ‘Savannah’ hypothesis for early bilaterian evolution : [англ.] / Graham Budd, Sören Jensen // Biological Reviews. — 2017. — Vol. 92, no. 1. — P. 446–473.
- Don Reis, Mario. Uncertainty in the timing of origin of animals and the limits of precision in molecular timescales : [англ.] / Mario Don Reis, Yuttapong Thawornwattana, Konstantinos Angelis … [et al.] // Current Biology. — 2015. — Vol. 25, no. 22. — P. 2939—2950.
- Cunningham, John A. The origin of animals: can molecular clocks and the fossil record be reconciled? : [англ.] / John A Cunningham, Alexander G Liu, Stefan Bengtson … [et al.] // BioEssays. — 2017. — Vol. 39, no. 1. — P. 1—12.
Ссылки
- Марков А. Сегменты у насекомых развиваются по часам (26 апреля 2012).
- Малахов В. В. (2004). Новые взгляды на происхождение билатерально-симметричных животных. Биология моря 30 (5): 335—346. Текст (рус.) (Дата обращения: 5 июня 2011)
Классификация животных | |
|---|---|
| Губки | |
| Пластинчатые |
|
| Гребневики | |
| Стрекающие | |
| Двусторонне- симметричные | |
| Таксономия | |
|---|---|
| В библиографических каталогах |