Диатомовые водоросли
| Диатомовые водоросли | |
|---|---|
 Fragilariopsis kerguelensis | |
| Научная классификация | |
Домен: Эукариоты Клада: Клада: Клада: Страменопилы Клада: Gyrista Отдел: Охрофитовые водоросли Класс: Диатомовые водоросли | |
| Международное научное название | |
| Bacillariophyceae Haeckel, 1878 | |
| Порядки | |
| |
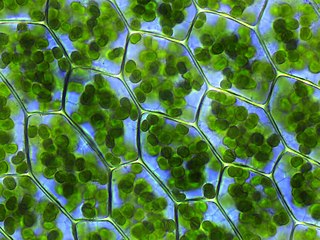
Диатомовые водоросли, или диатомеи (лат. Diatomeae), или бациллариофициевые водоросли (лат. Bacillariophyceae) — группа водорослей, отличающаяся наличием у клеток своеобразного «панциря», состоящего из диоксида кремния. Всегда одноклеточны, но встречаются колониальные формы. Обычно планктонные или перифитонные организмы, морские и пресноводные.
Являясь важнейшей составляющей морского планктона, диатомовые создают до четверти всего органического вещества планеты[1].
Особенности строения
Только коккоиды, форма разнообразна. В основном одиночные, реже — колониальные.
Оболочка клетки не гомогенна. Снаружи панциря, как и внутри него, располагается тонкий слой органического вещества.
Традиционно диатомовые водоросли делят на две группы — пеннатные, обладающие билатеральной симметрией, и центрические, с радиальной симметрией[2][Пр. 1].
Покровы

Для диатомовых характерно наличие особого, состоящего из кремнезёма, покрова — «панциря». Кроме кремнезёма, в состав панциря входит небольшое количество железа, алюминия, магния и органических веществ. У морских планктонных диатомей вещество панциря содержит 95,6 % SiO2 и 1,5 % Al2O3 или Fe2O3[3]. В редких случаях (например, у Phaeodactylum tricornutum) кремнезём отсутствует[4]. Поверхность панциря покрыта тонким пектиновым слоем[5].
Структура и характер орнаментации — важный признак для идентификации видов диатомей; она хорошо различима, когда протопласт удалён[6]. Необходимые для определения панцири, освобождённые от органических частей клетки, получают прокаливанием или промыванием в крепких кислотах[Пр. 2]. Рассматривают препараты панцирей, заключая их в среду с высоким коэффициентом преломления — в монобромнафталин, стиракс, среду Кольбе[2].
Панцирь состоит из двух половинок, большей и меньшей, входящих друг в друга как части чашки Петри. При делении половинки панциря расходятся, и новые половинки формируются в борозде деления. У обеих дочерних клеток старая половинка панциря становится большей (эпитекой, см. ниже), а меньшая достраивается заново. При этом размеры клеток в ряду делений постепенно уменьшаются. Восстановление размера происходит при половом размножении[7] или через стадию образования спор.
По типу симметрии клетка диатомовой водоросли, если рассматривать её со створки, может быть[8][9]:
- Радиальной (актиноморфной), такой тип симметрии характерен для центрических диатомей,
- Билатеральной (зигоморфной), у пеннатных диатомей. Чаще концы створок одинаковые (изопольные створки), иногда концы створок различаются по форме (гетеропольные створки).
Прежде группы центрических и пеннатных диатомей рассматривались в ранге классов, выделяемых на основе чисто морфологических признаков[2].
Выделяют также два дополнительных типа симметрии[8]:
- Триллиссоидный — при этом структуры створки располагаются по дугам и радиусам окружности, центр которой расположен вне клетки (например, у Eunotia) и
- Гоноидный, с угловатой створкой (у Triceratium).
Терминология
При описания панциря используется следующая терминология:
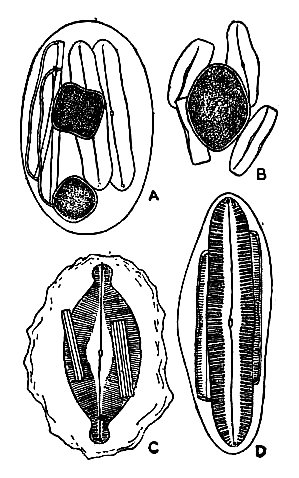
Эпитека — бо́льшая половинка панциря, его «крышечка», гипотека — меньшая его половинка. Поверхность створки эпитеки называют эпивальва, гипотеки — гиповальва. Поясковый ободок эпитеки — эпицингулюм, гипотеки — гипоцингулюм. Оба поясковых ободка, вложенные друг в друга, образуют поясок. На изображении различают вид панциря со створки и вид панциря с пояска[8][9].
Створка обычно плоская, её край называют загибом створки. Между поясковым ободком и загибом створки могут развиваться дополнительные один или несколько вставочных ободков. Количество вставочных ободков может увеличиваться при росте клетки, самый молодой из них располагается возле загиба створки. Вставочные ободки могут быть кольцевыми, воротничковыми или состоять из нескольких частей — полукольцевые, ромбовидные, чешуйчатые. На вставочных ободках могут развиваться направленные внутрь клетки неполные перегородки — септы. Септы всегда имеют одно или несколько отверстий[10].
Многие пеннатные диатомеи имеют шов — центральную щель, проходящую вдоль створки. Шов может быть S-образным. В области шва могут иметься утолщения панциря: центральный узелок и полярные узелки. Некоторые пеннатные диатомеи на месте шва имеют участок, лишённый орнаментации — осевое поле. Здесь может формироваться ложный шов — продольное ребро панциря. Диатомеи, лишенные шва, называют бесшовными[11].
Перфорация
Связь протопласта со внешней средой обеспечивается за счёт перфораций панциря. Перфорация может отсутствовать только на отдельных участках панциря и занимает от 10 до 75 % его площади[12].
Многослойная структура оболочки содержит большое количество крошечных отверстий, которые в свою очередь ведут к более крошечным отверстиям — такие структуры фокусируют свет на хлоропластах[13].
Формирование панциря
При делении каждая дочерняя клетка получает половину панциря от родительской. Полученная половинка становится эпитекой, гипотеку клетка достраивает заново. В результате деления одна из клеток сохраняет размер материнской, а вторая становится меньше. Энергия, необходимая для формирования панциря, получается за счёт аэробного дыхания; энергия, полученная в результате фотосинтеза, непосредственно не используется[14].
Наличие растворённого кремнезёма в окружающей среде — абсолютно необходимое условие для деления диатомей.
В воде кремнезём присутствует в виде кремниевой кислоты:
, либо


При увеличении концентрации раствора при pH менее 9 или при уменьшении pH насыщенного раствора, кремниевая кислота выпадает в осадок в виде аморфного кремнезёма. Хотя кремний — один из наиболее распространённых элементов земной коры, его доступность для диатомей ограничена растворимостью. Среднее содержание кремния в морской воде — около 6 ppm. Морские диатомовые быстро исчерпывают запасы растворённого кремнезёма в поверхностном слое воды, и это ограничивает их дальнейшее размножение.
Кремний поступает в клетки диатомовых в форме Si(OH)4 через белки транспорта кремнёвой кислоты (SIT). Каким образом происходит транспорт внутрь клетки, до сих пор не известно, и нет явных доказательств, активный он или пассивный (Curnow et al., 2012); предположительно, происходит у морских диатомовых симпортно с ионами натрия, у пресноводных — возможно, что также и с ионами калия. У морских видов Si(OH)4 и Na+ переносятся в соотношении 1:1. Обнаружено по несколько генов у разных видов диатомовых, имеющих отношение к транспорту кремниевой кислоты (GenBank). Германий нарушает транспорт кремниевой кислоты у диатомовых[15].
В процессе цитокинеза клеточная мембрана формирует перетяжку, а комплекс Гольджи начинает производить полупрозрачные везикулы, собирающиеся у вновь сформированной мембраны. Везикулы сливаются, образуя силикалемму пузыря, в котором формируется створка. Пузырь увеличивается в размерах и приобретает форму будущей створки. Комплекс Гольджи продолжает производить везикулы, которые транспортируют вещество будущей створки к пузырю и объединяются с ним. Кремний в пузыре откладывается в виде аморфных сфер диаметром 30—50 нм. Низкий pH внутри пузыря способствует агрегации частиц кремнезёма, и препятствует растворению сформированных частиц.
Вероятно, в процессе формирования створки участвует центр организации микротрубочек, который в этот момент перемещается в положение между ядром и силикалеммой[16]. Детали этого процесса пока неясны.
В осаждении кремния участвуют силаффины — пептиды, катализирующие поликонденсацию кремниевой кислоты[17]. Силаффины разных видов диатомовых водорослей различаются, соответственно, различается и характер кремнезёмных гранул, осаждаемых с их участием. Предполагается, что с этим могут быть связаны различия в орнаментации панциря у различных видов диатомей[15][18].
После формирования створки, подобным же образом, в собственной силикалемме формируется поясок[15] и вставочные ободки.
Хлоропласты
У диатомовых хлоропласты разнообразной формы, обычно пристенные. У центрических диатомей они обычно многочисленные, мелкие, у пеннатных крупные, часто лопастные. Хлоропласты имеют типичное для охрофитов строение. Пиреноидов может быть несколько, они выступают за пределы хлоропласта и иногда пронизаны тилакоидами.
Окраска хлоропластов бурая, желтоватая или золотистая. Она обусловлена тем, что зелёные хлорофиллы маскируются добавочными каротиноидами (бурый пигмент диатомин; β, ε — каротины; ксантофиллы: фукоксантин, неофукоксантин, диадиноксантин, диатоксантин). У большинства диатомей содержатся две формы хлорофилла c: c1 и c2. У ряда форм хлорофилл c1 может замещаться хлорофиллом c3 (найден также у примнезиофитовых и пелагофициевых). У некоторых видов могут присутствовать все три формы хлорофилла c, в то время как у других форма только одна.
Другие структуры
Большая часть клетки диатомей приходится на вакуоль с клеточным соком, цитоплазма занимает постеночное положение. Кроме того, цитоплазма скапливается в центре клетки в виде цитоплазматического мостика, соединённого с периферическим слоем цитоплазмы. В мостике расположено ядро. В цитоплазме множество капель масла. В виде крупных капель с характерным голубым блеском в ней встречается волютин. Присутствует хризоламинарин.
Митохондрии у диатомей разнообразной формы (шаровидные, овальные, палочковидные, нитчатые). Аппарат Гольджи расположен рядом с ядром, он состоит из нескольких диктиосом (до 20), которые содержат от 4 до 12 цистерн.
Ядро крупное, содержит 1—8 ядрышек, которые исчезают во время митоза. Центриоли отсутствуют. Центром организации микротрубочек являются пластинки, расположенные на полюсах веретена. Микротрубочки веретена формируются вне ядра, затем проходят в ядро через разрушенные участки его оболочки; ядерная оболочка постепенно исчезает. Таким образом, у диатомей митоз открытый. На ранних этапах микротрубочки идут от полюса к полюсу. Кинетохоры хромосом, по-видимому, прикрепляются к полюсным микротрубочкам. В анафазе хромосомы двигаются к полюсам, в поздней анафазе веретено удлиняется.
Колониальные формы

У некоторых диатомей клетки после деления не расходятся, образуя колонии. Клетки в колониях не связаны между собой, плазмодесмы отсутствуют.
Клетки могут соединяться всей поверхностью створок, образуя нитевидные (у Melosira) или лентовидные (у Fragilaria) колонии.
Если клетки имеют клиновидную форму, формируется веерообразная колония (у Meridion).
Клетки, соединённые только уголками, формируют колонии в виде цепочек (Tabellaria) или звёздочек (Asterionella).
Колонии могут иметь вид слизистых трубок, внутри которых содержатся отдельные подвижные (Navicula, Cymbella) или неподвижные клетки.
Колониальные диатомеи могут быть планктонными и прикреплёнными к субстрату слизистой ножкой[19].
Жизненный цикл
Вегетативное размножение
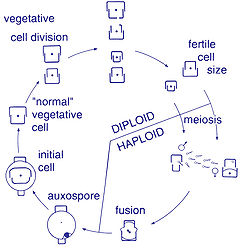

Вегетативное размножение диатомей происходит путём простого митотического деления. Цитокинез имеет ряд особенностей, связанных с наличием панциря (см. формирование панциря)[20]. Поскольку получаемая от родительской клетки половинка панциря у дочерней становится эпитекой, а гипотека достраивается заново, размеры одной из клеток остаются равными родительской, а второй становятся меньше. В ряде последовательных делений размеры клеток в популяции уменьшаются, а исходные максимальные размеры восстанавливаются в процессе полового воспроизведения, связанного с формированием ауксоспор. Ауксоспоры могут возникать автогамно за счёт слияния двух гаплоидных ядер одной клетки или апогамно (из вегетативных клеток)[21]. В редких случаях возможен выход цитоплазмы из панциря и формирование его заново — вегетативное укрупнение[22].
Споры и покоящиеся клетки
При наступлении неблагоприятных условий некоторые диатомовые могут формировать споры и покоящиеся клетки. Эти структуры богаты запасными веществами, которые потребуются при прорастании. Покоящиеся клетки морфологически близки к вегетативным клеткам, в то время как панцирь спор становится более толстым, округляется, изменяется его орнаментация. Покоящиеся клетки могут возникать в условиях с низким содержанием растворённого кремния, а споры, наоборот, требуют наличия достаточного количества кремния для построения собственной толстой оболочки. Покоящиеся клетки формируются чаще пресноводными центрическими и пеннатными диатомеями, в то время как споры — центрическими морскими диатомовыми. И покоящиеся клетки, и споры могут выживать десятилетиями. При их прорастании для формирования нормального панциря требуется два митоза с дегенерацией ядер. Споры морских диатомей играют важную роль в транспорте органического углерода и кремния в осадочные отложения.
При образовании спор клетка теряет вакуоли, и размер споры оказывается меньше исходной клетки[23].
Половой процесс
Половым путем размножаются крайне редко.
Движение
Многие шовные пеннатные и некоторые центрические диатомеи способны ползать по субстрату.
Экология

Диатомеи широко распространены во всевозможных биотопах. Они живут в океанах, морях, солоноватых и различных пресных водоёмах: стоячих (озёрах, прудах, болотах и т. д.) и текучих (реках, ручьях, оросительных каналах и др.). Они распространены в почве, их выделяют из образцов воздуха, они образуют богатые сообщества во льдах Арктики и Антарктики. Такое широкое распространение диатомовых обусловлено их пластичностью по отношению к различным экологическим факторам и в то же время существованием видов, узко приспособленных к экстремальным значениям этих факторов.
Диатомовые в водных экосистемах доминируют над другими микроскопическими водорослями круглый год. Они обильны как в планктоне, так и в перифитоне и бентосе. В планктоне морей и океанов преобладают центрические диатомеи, хотя к ним примешиваются и некоторые пеннатные. В планктоне пресных водоёмов, наоборот, преобладают пеннатные. Бентосные ценозы также отличаются большим разнообразием и количеством диатомовых, которые обычно обитают на глубине не более 50 м. Жизнь бентосных диатомей обязательно связана с субстратом: они ползают по субстрату или прикрепляются к нему с помощью слизистых ножек, трубок, подушечек.
Наиболее богаты по качественному и количественному составу диатомей ценозы обрастаний. Диатомеи занимают господствующее положение среди обрастаний высших растений и макроскопических водорослей в пресных водоёмах и морях. Обрастанию могут подвергаться многие животные (такие водоросли называются эпизоонтами) от ракообразных до китов. Среди диатомей встречаются и эндобионты, которые обитают в других организмах, например в бурых водорослях, фораминиферах.
Видовой состав диатомей в водоёмах определяется комплексом абиотических факторов, из которых большое значение в первую очередь имеет солёность воды. Не менее важным фактором для развития диатомей являются температура, степень освещённости и качество света. Диатомовые вегетируют в диапазоне 0—70 °C, но в состоянии покоя способны переносить как более низкие, так и более высокие температуры.
Диатомовые — фототрофные организмы, но среди них встречаются миксотрофы, симбиотрофы, а также бесцветные гетеротрофные формы.
Геном
Недавно при расшифровке полного генома диатомовой водоросли Phaeodactylum tricornutum выяснилось, что он содержит рекордное для эукариот число генов, полученных путём горизонтального переноса от бактерий и архей[24].
Филогения
Створки диатомовых водорослей не растворяются в большинстве природных вод, поэтому они осаждаются на протяжении последних 150 млн лет, начиная с раннего мелового периода. Таким образом, есть основания полагать, что диатомеи появились до наступления мелового периода. Наиболее древние ископаемые диатомеи были центрическими, в то время как самые древние пеннатные были бесшовными из позднего мелового периода (около 70 млн лет назад). Ископаемые остатки шовных диатомей имеют более поздний возраст. Согласно остаткам пресноводные диатомеи появились около 60 млн лет назад и достигли расцвета в миоцене (24 млн лет назад). Палеонтологические данные подтверждают наличие более примитивных признаков в организации центрических диатомей, как древней группы, в то время как снабжённые швом пеннатные представляют вершину эволюции этой группы. Методами молекулярной биологии было показано, что диатомовые — монофилетичная группа, но внутри этой группы центрические диатомеи не формируют, как ранее считали, монофилетичную группу.
Наличие трёхчастных мастигонем на жгутике, строение хлоропластов, пигментные системы, трубчатые митохондрии, запасные продукты — всё это подтверждает несомненную принадлежность диатомовых водорослей к группе охрофитовых. Чаще всего дискутируется вопрос об их близости к другим классам этого отдела, так как наличие таких особенностей, как кремнезёмный панцирь, диплобионтный жизненный цикл, редукция жгутикового аппарата, особенности карио- и цитокинеза, значительно отличают диатомей от других представителей охрофитовых. Предполагали, что предками диатомей могли быть какие-то древние синуровые. Некоторые авторы даже рассматривали синуровых как «жгутиковых диатомей». Однако данные молекулярной биологии показывают, что среди страминопил диатомовые образуют достаточно обособленную группу, которая отстоит от других охрофитовых водорослей дальше, чем сами они отделены друг от друга, но всё-таки ближе к охрофитовым, чем к грибоподобным протистам. Анализ последовательности нуклеотидов генов SSU rDNA rbcL и пигментного состава показали, что внутри охрофитовых сестринской линией диатомовым является недавно открытая группа болидофициевых — окрашенных двужгутиковых монад, обитающих в океанах и морях.
Диатомовые относятся к группе гетероконтных водорослей, имеющих вторичные пластиды. По молекулярным данным установлено, что предком их пластид была красная водоросль. С другой стороны, молекулярно-генетический анализ ядерных генов, продукты которых обеспечивают работу пластид, восходят как к красным, так и к зелёным водорослям, в примерно равном количестве. Считается, что это указывает на существование у их предков хлоропласта, чьим предком был хлоропласт зелёной водоросли, в течение более или менее продолжительного периода[25].
Значение
Силаффины диатомовых перспективны для использования в области нанотехнологий, для получения материалов на основе диоксида кремния с заранее заданными свойствами[26][27]. В криминалистике разнообразие диатомей позволяет с высокой точностью идентифицировать водоемы.
Систематика
Считается, что к классу диатомей относится около 300 родов, включающих 20—25 тысяч видов, но некоторые авторы убеждены, что истинное количество видов диатомей может достигать 200 тысяч. Самый большой род, состоящий более чем из 10 тысяч видов — Навикула (Navicula).
В настоящее время нет устоявшейся системы диатомовых. В большинстве работ, которые касаются изучения флор диатомей, систематики и классификации, класс диатомей рассматривается в ранге отдела (Bacillariophyta) с тремя классами (Coscinophyceae, Fragilariophyceae, Bacillariophyceae). Вместе с тем использование методов молекулярной биологии показало, что Coscinophyceae и Fragilariophyceae — парафилетичные группы и требуется дальнейший пересмотр системы диатомовых водорослей, однако, с точки зрения молекулярной биологии и филогенетики наиболее верной является классификация диатомей в составе отдела Охрофитовые (Ochrophyta)[28].
Галерея
 Диатомовые водоросли
Диатомовые водоросли Haeckel Diatomea
Haeckel Diatomea Различные диатомовые водоросли
Различные диатомовые водоросли Nitzschia-kerguelensis
Nitzschia-kerguelensis- Веерообразная колония Meridion circulare
 Жизненный цикл пеннатных диатомовых
Жизненный цикл пеннатных диатомовых Жизненный цикл центрических диатомовых
Жизненный цикл центрических диатомовых Цикл океанического кремния
Цикл океанического кремния Диатомовая водоросль
Диатомовая водоросль Несколько видов пресноводных диатомовых водорослей
Несколько видов пресноводных диатомовых водорослей
См. также
- Atthea
- Hannaea arcus
- Константин Сергеевич Мережковский
- Диатомовый ил
- Биология
- Водоросли
Примечания
Комментарии
- ↑ Прежде эти группы рассматривались в ранге классов Centricae и Pennatae. Выделялся также класс Mediatae. К центрическим диатомовым относили порядки:
- Thalassiosirales
- Coscinodiscales
- Melosirales
- Chaetocerotales;
- Fragilariales
- Tabellariales
- Achnanthales
- Cymbellales
- Naviculales
- Bacillariales
- Surirellales.
- ↑ Обычно химическая обработка собранного материала имеет следующий порядок: проба очищается от грубых примесей, промывается соляной кислотой для освобождения от извести, затем кипятится 10—20 минут в крепкой серной кислоте. После кипячения к горячему раствору прибавляют немного селитры, выделяющаяся азотная кислота заканчивает окисление. Проба тщательно промывается водой, концентрируется центрифугированием, материал наносится на покровное стекло и высушивается (Курсанов, 1953, с. 10).
Источники
- ↑ Белякова, 2006, с. 89.
- ↑ 1 2 3 Курсанов, 1953, с. 10.
- ↑ Lee, 2008, с. 373.
- ↑ Саут, 1990, с. 94.
- ↑ Белякова, 2006, с. 79.
- ↑ Саут, 1990, с. 91.
- ↑ Саут, 1990, с. 200.
- ↑ 1 2 3 Саут, 1990, с. 93.
- ↑ 1 2 Белякова, 2006, с. 79—80.
- ↑ Белякова, 2006, с. 79—82.
- ↑ Курсанов, 1953, с. 8.
- ↑ Белякова, 2006, с. 81.
- ↑ Водоросли оказались удивительными оптическими системами — «Мембрана.ру». Дата обращения: 6 января 2018. Архивировано из оригинала 3 июля 2017 года.
- ↑ Lee, 2008, с. 374.
- ↑ 1 2 3 Lee, 2008, с. 376.
- ↑ Белякова, 2006, с. 85.
- ↑ Kroger, 1999.
- ↑ Poulsen, 2003, с. 12075.
- ↑ Курсанов, 1953, с. 9.
- ↑ Саут, 1990, с. 199.
- ↑ Белякова, 2006, с. 88.
- ↑ Stosch, 1965, с. 21—44.
- ↑ Lee, 2008, с. 382.
- ↑ Чужие гены помогли диатомовым водорослям добиться успеха — «Элементы.ру». Дата обращения: 6 января 2018. Архивировано 29 ноября 2016 года.
- ↑ Diatom genes originating from red and green algae: Implications for the secondary endosymbiosis models (англ.) // Marine Genomics. — 2019-06-01. — Vol. 45. — P. 72–78. — ISSN 1874-7787. — doi:10.1016/j.margen.2019.02.003. Архивировано 10 апреля 2021 года.
- ↑ Marner, 2008, с. 1—5.
- ↑ Микросхемы из микроорганизмов — на сайте «Элементы.ру», 12.10.05. Дата обращения: 6 января 2018. Архивировано 4 марта 2016 года.
- ↑ Белякова, Г. А., Ю. Т. Дьяков, К. Л. Тарасов. Ботаника : учебник для студ. высш. учеб. заведений : в 4 т. // Т. 2: Водоросли и грибы // Отдел охрофитовые водоросли (охрофиты) — Ochrophyta / Г. А. Белякова. — Москва : Академия, 2006. — С. 50—138. — 320 с. — ISBN — 5-7695-2730-7. — ISBN 5-7695-2750-1 (т. 2).
Литература
- Диатомовые водоросли // Энциклопедический словарь Брокгауза и Ефрона : в 86 т. (82 т. и 4 доп.). — СПб., 1890—1907.
- Белякова Г. А. Водоросли и грибы // Ботаника: в 4 тт. / Белякова Г. А., Дьяков Ю. Т., Тарасов К. Л. — М.: Издательский центр «Академия», 2006. — Т. 2. — 320 с. — 3000 экз. — ISBN 5-7695-2750-1.
- Киселёв И. А., Зинова А. Д., Курсанов Л. И. Водоросли // Определитель низших растений (в пяти томах) / под ред. Курсанов Л. И. — М.: Государственное издательство «Советская Наука», 1953. — Т. 2. — 15 000 экз.
- Саут Р., Уиттик А. Основы альгологии = Introduction to Phycology / пер. Тарасов К. Л. — М.: «Мир», 1990. — 597 с. — 3000 экз. — ISBN 5-03-001522-1.
- Скабичевский А.П. Планктонные диатомовые водоросли пресных вод СССР. — М., 1960. Архивная копия от 1 апреля 2016 на Wayback Machine
- Lee, R. E. Phycology, 4th edition. — Cambridge: Cambridge University Press, 2008. — 547 p. — ISBN 9780521682770.
- Kröger N., Deutzmann R., Sumper M. Polycationic peptides from diatom biosilica that direct silica nanosphere formation (англ.) // Science. — 1999. — Vol. 286, no. 5. — P. 1129—1132. — PMID 10550045.
- Poulsen N., Sumper M., Kröger N. Biosilica formation in diatoms: Characterization of native silaffin-2 and its role in silica morphogenesis (англ.) // PNAS. — 2003. — Vol. 100, no. 21. — P. 12075—12080. — doi:10.1073/pnas.2035131100. — PMID 14507995.
- Stosch H. A. von. Manipulierung der Zellgrösse von Diatomeen in Experiment (нем.) // Phycologia. — 1965. — Nr. 5. — P. 21—44.
- Marner W. D. II, Shaikh A. S., Muller S. J., Keasling J. D. Morphology of Artificial Silica Matrices Formed via Autosilification of a Silaffin/Protein Polymer Chimera (англ.) // Biomacromolecules. — 2008. — Vol. 9, no. 1. — P. 1—5. — doi:10.1021/bm701131.
Ссылки
- База данных диатомовых водорослей (англ.).
- Геном диатомовых водорослей (англ.).
- Чужие гены помогли диатомовым водорослям добиться успеха — «Элементы.ру»
- Plankton*Net, таксонемическая база данных
- Edlund M., Kociolek P., Lowe R., Potapova M., Rushforth S., Spaulding S., Stoermer G.. Diatoms of the United States - хорошо иллюстрированный ресурс, посвящённый диатомовым США (англ.). Дата обращения: 13 июля 2014.
Классификация Sar | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Страменопилы |
| ||||||||||||||||
| Альвеоляты |
| ||||||||||||||||
| Ризарии |
| ||||||||||||||||