Диминуция хроматина
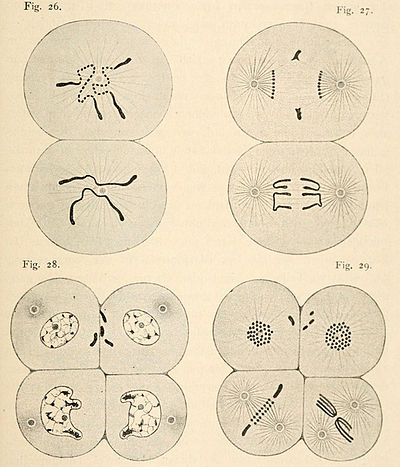

Диминуция хроматина (от лат. diminutio — уменьшение) — общее название клеточных генетических процессов, в ходе которых при эмбриональном развитии некоторых многоклеточных животных (в основном, беспозвоночных) соматические клетки запрограммировано теряют часть генетического материала, присутствовавшего в зиготе и остающегося нетронутым в клетках зародышевой линии. Диминуция хроматина наблюдается у некоторых представителей двукрылых насекомых, паразитических круглых червей (нематод), веслоногих ракообразных, а также миксиновых[1]. Механизмы диминуции у различных организмов отличаются, но объединяет их то, что теряется в основном повторяющаяся и некодирующая ДНК и происходит это только в зачатках соматических тканей[2]. Аналогичный процесс существует также у простейших, а именно у инфузорий, у некоторых из которых в ходе реорганизации вегетативного ядра (макронуклеуса) теряется значительная часть генетического материала, имеющегося в генеративном ядре (микронуклеусе) — аналоге клеток зародышевой линии многоклеточных животных.
Явление диминуции хроматина было открыто и описано при помощи цитологических методов у аскарид в 80-х годах XIX века немецким биологом Теодором Бовери[3].
Диминуция хроматина у нематод
Диминуция хроматина наблюдается у 12 видов нематод, ведущих паразитический образ жизни, и не обнаружена у свободноживущих нематод. Как было указано выше, диминуцию хроматина у круглых червей открыл в XIX веке Теодор Бовери, изучая эмбриональное развитие аскариды Ascaris megalocephala[3]. Значительный вклад в изучение механизмов диминуции хроматина у нематод внесли в XX веке швейцарские зоологи Хайнц Тоблер (Heinz Tobler) и Фриц Мюллер (Fritz Müller), основными объектами изучения которых были аксариды Parascaris univalens и свиная аскарида Ascaris suum[4].
В ходе диминуции хроматина у нематод происходит необратимая дифференцировка клеток соматической и зародышевых линий, заключающаяся во фрагментации хромосом в клетках соматической линии, добавлении новых теломерных последовательностей к полученным фрагментам и в последующей элиминации гетерохроматиновых сегментов хромосом. В результате этого процесса клетки зародышевой и соматической линий различаются по диплоидному числу хромосом, количеству ДНК и структуре ядерного хроматина[5].
Диминуция хроматина у P.univalens происходит последовательно со 2-го по 6-е деление дробления. Цитологический анализ голоцентрических хромосом аскарид Parascaris показал, что в анафазе диминуционного деления к полюсам веретена деления мигрируют только эухроматиновые сегменты хромосом, тогда как теломерные блоки гетерохроматина остаются в экваториальной зоне и в дальнейшем мигрируют в цитоплазму, где вскоре деградируют. Диплоидное число хромосом в клетках соматической линии увеличивается с двух до шестидесяти. По различным подсчётам у P.univalens при диминуции хроматина элиминируется около 80-90 % тотальной ДНК[1].
Диминуция хроматина у копепод
В настоящее время диминуция хроматина обнаружена у 22 видов копепод Cyclopoida (Copepoda, Crustacea)[1]. Первые наблюдения диминуции хроматина у копепод относятся к 1911 году[6]. Диминуцию хроматина у веслоногих рачков изучала в 60-70-е годы XX века немецкая исследовательница Зигрид Берман (Sigrid Beermann), а также, начиная с 80-х годов XX века, российские учёные А. П. Акифьев, А. К. Гришанин и И. Ф. Жимулёв[7].
В отличие от нематод, где диминуция хроматина проходит последовательно в нескольких делениях дробления, диминуция хроматина у циклопа Cyclops kolensis происходит практически одновременно. Так, диминуция хроматина происходит во время 4-го деления дробления одновременно в 6 клетках из восьми; в 7-й клетке диминуция наступает в то время, когда идёт пятое деление в клетках, уже прошедших диминуцию; оставшаяся 8-я клетка диминуцию не претерпевает, формируя линию зародышевого пути. В результате диминуции хроматина у Cyclops kolensis соматические клетки содержат примерно на 90% ДНК меньше, чем клетки зародышевого пути[7].
Диминуция хроматина у инфузорий

Процесс диминуции хроматина характерен для всех представителей родов инфузорий Tertahymena, Paramecium (класс Oligohymenophorea), Stylonychia, Euplotes и Oxytricha (класс Spirotrichea)[1]. Диминуция хроматина была открыта у инфузорий американским исследователам Дэвидом Прескоттом (David M. Prescott) в 80-х годах XX века спустя сто лет после открытия этого процесса у аскарид Теодором Бовери[8].
Микронуклеус инфузорий является аналогом клеток зародышевой линии многоклеточных организмов, в то время как макронуклеус является соматическим ядром, которое образуется из копии микронуклеуса и которое поддерживает существование данной конкретной особи. Микронуклеус содержит хромосомы, типичные по размеру и составу для эукариот. Теломерами хромосом микронуклеуса служат сотни раз повторённые последовательности 5'-CCCCAAAA-3'. Макронуклеарная ДНК состоит из фрагментов размерами, варьирующими от нескольких сотен пар оснований (п.о.) до примерно 15 тысяч п.о. со средним значением около 2 тыс. п.о. За очень небольшим исключением каждый из фрагментов макронуклеарной ДНК представляет собой один ген (одну рамку считывания) с 5'-некодирующей регуляторной последовательностью и 3'-некодирующим «хвостом». Концы этих фрагментов имеют другую структуру теломер по сравнению с микронуклеарными хромосомами и состоят из многократных повторов последовательности 5'-C4A4C4A4C4- 3'[8].
В ходе созревания макронуклеуса микронуклеарные хромосомы вначале становятся политенными, затем эти политенные хромосомы разрезаются на фрагменты, к которым теломераза добавляет теломерные повторы, далее спейсерная (межгенная) ДНК элиминируются, а оставшиеся фрагменты, содержащие гены, многократно амплифицируются. У инфузории Oxytricha nova элиминуется около 95 % ДНК, первоначально имевшейся в микронуклеусе, а макронуклеус в итоге реорганизации имеет порядка 25*106 коротких фрагментов ДНК[8]. Самое поразительное в этих реорганизациях — изменение порядка расположения отрезков структурных генов, называемых «предназначенными для макронуклеуса последовательностями» — MDS (англ. macronuclear destined sequence). Например, ген актина 1 у Oxytricha nova имеет в микронуклеусе порядок MDS-отрезков 3-4-6-5-7-9-2-1, а в макронуклеусе MDS-отрезки становятся в порядке 1-2-3-4-5-6-7-8-9, обеспечивающем нормальную транскрипцию гена[9].
См. также
Примечания
- ↑ 1 2 3 4 Гришанин А.К., Шеховцов А.К., Бойкова Т.В., Акифьев А.П., Жимулев И.Ф. Проблема диминуции хроматина на рубеже XX и XXI веков // Цитология. — 2006. — Т. 48, № 5. — С. 379—397. — PMID 16892848. Архивировано 4 марта 2016 года.
- ↑ Коряков Д.Е., Жимулёв И.Ф. Хромосомы. Структура и функции / ред. Л.В.Высоцкая. — Новосибирск: Из-во СО РАН, 2009. — С. 178-198.
- ↑ 1 2 Satzinger H. Theodor and Marcella Boveri: chromosomes and cytoplasm in heredity and development (англ.) // Nat. Rev. Genet. : journal. — 2008. — March (vol. 9, no. 3). — P. 231—238. — doi:10.1038/nrg2311. — PMID 18268510. Архивировано 21 июля 2009 года.
- ↑ Müller F., Tobler H. Chromatin diminution in the parasitic nematodes ascaris suum and parascaris univalens (англ.) // International Journal for Parasitology[англ.] : journal. — Elsevier, 2000. — April (vol. 30, no. 4). — P. 391—399. — PMID 10731562.
- ↑ Jentsch S., Tobler H., Müller F. New telomere formation during the process of chromatin diminution in Ascaris suum (англ.) // Int. J. Dev. Biol.[англ.] : journal. — 2002. — January (vol. 46, no. 1). — P. 143—148. — PMID 11902675. Архивировано 30 июня 2012 года.
- ↑ Amma K. Über die Differenzierung der Keimbahnzellen bei den Copepoden (нем.) : дис. — 1911.
- ↑ 1 2 Grishanin A. K., Zagoskin M. V. Chromatin Diminution in Cyclops kolensis Lill.(Copepoda, Crustacea) as a Radical Way to Inactivate Redundant Genome in Somatic Cells (англ.) // Cytogenetic and genome research. — 2018. — Vol. 156, no. 3. — P. 165—172. — doi:10.1159/000494157. Архивировано 26 мая 2019 года.
- ↑ 1 2 3 Prescott D. M. The unusual organization and processing of genomic DNA in hypotrichous ciliates (англ.) // Trends Genet.[англ.] : journal. — 1992. — December (vol. 8, no. 12). — P. 439—445. — PMID 1337226.
- ↑ Акифьев А. П., Гришанин А. К., Дегтярев С. В. Диминуция хроматина — ключевой процесс для объяснения парадокса размера генома эукариот и некоторых механизмов генетической изоляции // Генетика. — 2002. — Т. 38, № 5. — С. 595—606. — PMID 12068542.
Литература
- Коряков Д.Е., Жимулёв И.Ф. Хромосомы. Структура и функции / ред. Л.В.Высоцкая. — Новосибирск: Из-во СО РАН, 2009. — С. 178-198.
Ссылки
- Акифьев А.П. Избыточная ДНК — генетическая квадратура круга? "Природа", №10, 2004 г. (1 сентября 2004). Дата обращения: 21 августа 2012.