Динамика белков

Считается, что белки имеют уникальные структуры, определяемые их аминокислотными последовательностями. Однако белки не являются строго статическими объектами, а скорее представляют ансамбли (иногда похожие) конформаций. Переходы между этими состояниями происходят в различных масштабах длины (от десятых Å до нм) и временных масштабах (от нс до с) и связаны с функционально значимыми явлениями, такими как аллостерическая передача сигналов[1] и ферментативный катализ[2].
Изучение динамики белков наиболее непосредственно связано с переходами между этими состояниями, но также может включать природу и равновесные популяции самих состояний. Эти две точки зрения — кинетика и термодинамика соответственно — могут быть концептуально синтезированы в парадигме «энергетического ландшафта»[3]: распространённые состояния и кинетика переходов между ними может быть описана глубиной энергетических ям и высотой энергетических барьеров, соответственно.
Локальная гибкость: атомы и аминокислотные остатки
Части белковых структур часто отклоняются от состояния равновесия. Некоторые из таких отклонений являются гармоническими, например, стохастические колебания химических связей и валентных углов. Другие являются ангармоническими, например, боковые цепи, которые прыгают между отдельными дискретными минимумами энергии, или ротамеры[4].
Доказательства локальной гибкости часто получают с помощью ЯМР-спектроскопии. Гибкие и потенциально неупорядоченные области белка можно обнаружить с помощью индекса случайной катушки[англ.]. Гибкость свернутых белков можно определить путем анализа спиновой релаксации[англ.] отдельных атомов в белке. Гибкость также можно наблюдать на картах электронной плотности с очень высоким разрешением, созданных с помощью рентгеновской кристаллографии[5] особенно когда дифракционные данные собираются при комнатной температуре вместо традиционной криогенной температуры (обычно около 100 К)[6]. Информация о частотном распределении и динамике локальной гибкости белка может быть получена с помощью рамановской спектроскопии и оптической спектроскопии эффекта Керра в терагерцевой частотной области[7].
Региональная гибкость: внутридоменное соединение с несколькими остатками
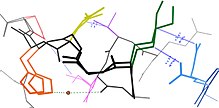
Многие остатки в белковых структурах находятся в непосредственной пространственной близости. Это верно для большинства остатков, которые являются смежными в первичной последовательности, но также и для многих остатков, которые являются дистальными по последовательности, но вступают в контакт в окончательной складчатой структуре. Из-за этой близости энергетические ландшафты этих остатков становятся связанными на основе различных биофизических явлений, таких как водородные связи, ионные связи и ван-дер-ваальсовы взаимодействия (см. рисунок). Таким образом, переходы между состояниями для таких наборов остатков становятся коррелированными[8].
Это, пожалуй, наиболее очевидно для открытых петель, которые часто коллективно смещаются, принимая различные конформации в различных кристаллических структурах (см. рисунок). Однако сопряженная конформационная гетерогенность также иногда очевидна во вторичной структуре[9]. Например, последовательные остатки и остатки, смещенные на 4 в первичной последовательности, часто взаимодействуют в α-спиралях. Кроме того, остатки, смещенные на 2 в первичной последовательности, направляют свои боковые цепи к одной и той же стороне β-листов и достаточно близки для стерического взаимодействия, как и остатки на соседних нитях одного и того же β-листа. Некоторые из этих конформационных изменений индуцируются посттрансляционными модификациями структуры белка, такими как фосфорилирование и метилирование[9][10].
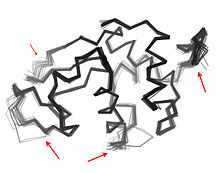
Когда эти связанные остатки образуют пути, связывающие функционально важные части белка, они могут участвовать в аллостерической передаче сигналов. Например, когда молекула кислорода связывается с одной субъединицей тетрамера гемоглобина, эта информация аллостерически распространяется на другие три субъединицы, тем самым повышая их сродство к кислороду. В этом случае сопряженная гибкость гемоглобина позволяет кооперативному связыванию кислорода, что является физиологически полезным, поскольку оно обеспечивает быструю кислородную нагрузку в ткани легких и быструю разгрузку кислородом в тканях, лишенных кислорода (например, в мышцах).
Глобальная гибкость: несколько доменов
Наличие нескольких доменов в белках дает большую гибкость и подвижность, что приводит к динамике белковых доменов[1]. Движения доменов могут быть выведены путем сравнения различных структур белка (как в базе данных молекулярных движений), или их можно непосредственно наблюдать с помощью спектров[11][12] измеренных с помощью спектроскопии спинового эха нейтронов. Они также могут быть предложены путем отбора проб в обширных траекториях молекулярной динамики[13] и анализа главных компонент[14]. Движение домена важно для:
- ABC транспортеры[15]
- Катализ[16]
- Клеточная локомоции и моторные белки[17]
- Образование белковых комплексов[18]
- Ионные каналы[19]
- Механорецепторы и механотрансдукция[20]
- Регулирующая деятельность[21]
- Транспорт метаболитов через клеточные мембраны
Одно из самых крупных наблюдаемых перемещений домена — это «поворотный» механизм в пируватфосфатдикиназе. Фосфоинозитидный домен переключается между двумя состояниями, чтобы переместить фосфатную группу из активного центра нуклеотид-связывающего домена в фосфоенолпируватный/пируватный домен[22]. Фосфатная группа перемещается на расстояние 45 Å с движением домена на около 100 градусов вокруг единственного остатка. В ферментах замыкание одного домена на другой захватывает субстрат за счет индуцированной подгонки, позволяя реакции протекать контролируемым образом. Подробный анализ Герштейна привел к классификации двух основных типов движения домена; шарнир и ножницы[19]. Только относительно небольшая часть цепи, а именно междоменный линкер и боковые цепи претерпевают значительные конформационные изменения при перестройке домена[23].
Шарниры с помощью вторичных структур
Исследование Хейворда[24] показало, что концы α-спиралей и β-листов во многих случаях образуют шарниры. Было обнаружено, что многие петли включают в себя два вторичных элемента конструкции, действующих как дверные петли, позволяя открывать и закрывать двери. Это может возникнуть, когда две соседние нити в β-листе, расположенные в одном домене, расходятся, когда они присоединяются к другому домену. Два результирующих конца затем образуют области изгиба между двумя доменами. Было обнаружено, что α-спирали, которые сохраняют свою сеть водородных связей при изгибе, ведут себя как механические шарниры, накапливая «упругую энергию», которая управляет закрытием доменов для быстрого захвата субстрата[24].
От спиральной до расширенной конформации
Взаимопревращение спиральных и протяженных конформаций на участке границы домена не является чем-то необычным. В кальмодулине торсионные углы изменяются для пяти остатков в середине домена, связывающего α-спираль. Спираль разделена на две почти перпендикулярные более мелкие спирали, разделенные четырьмя остатками удлиненной цепи[25][26].
Сдвиговые движения
Сдвиговые движения включают в себя небольшое скользящее движение границ раздела доменов, контролируемое боковыми цепями аминокислот внутри границы раздела. Белки, демонстрирующие сдвиговые движения, часто имеют многоуровневую архитектуру: складывание вторичных структур. Междоменный линкер выполняет лишь роль удержания доменов в непосредственной близости.
Движение домена и функциональная динамика в ферментах
Анализ внутренней динамики структурно различных, но функционально подобных ферментов выявил общую взаимосвязь между расположением активного сайта и двух основных субдоменов белка. Фактически, для некоторых членов суперсемейства гидролаз каталитический центр расположен близко к границе раздела двух основных квазижестких доменов[13]. Такое позиционирование, по-видимому, является инструментом для поддержания точной геометрии активного сайта, в то же время обеспечивая заметную функционально ориентированную модуляцию фланкирующих областей в результате относительного движения двух субдоменов.
Последствия для макромолекулярной эволюции
Данные свидетельствуют о том, что динамика белков важна для функционирования, например, для ферментативного катализа при DHFR, но также предполагается, что они облегчают приобретение новых функций в результате молекулярной эволюции[27]. Этот аргумент предполагает, что белки эволюционировали, чтобы иметь стабильные, в основном уникальные складчатые структуры, но неизбежная остаточная гибкость приводит к некоторой степени функциональной неразборчивости, которая может быть усилена/задействована/отклонена с помощью последующих мутаций.
Однако растет понимание того, что внутренне неструктурированные белки довольно распространены в геномах эукариот[28] ставит под сомнение простейшую интерпретацию догмы Анфинсена: «последовательность определяет структуру (единую)». Фактически, новая парадигма характеризуется добавлением двух оговорок: «последовательность и клеточная среда определяют структурный ансамбль».
Примечания
- ↑ 1 2 Protein Structure and Diseases. — 2011. — P. 163–221. — ISBN 9780123812629. — doi:10.1016/B978-0-12-381262-9.00005-7.
- ↑ "Hidden alternative structures of proline isomerase essential for catalysis". Nature. 462 (7273): 669—673. Dec 2009. Bibcode:2009Natur.462..669F. doi:10.1038/nature08615. PMID 19956261.
- ↑ "The energy landscapes and motions of proteins". Science. 254 (5038): 1598—1603. Dec 1991. Bibcode:1991Sci...254.1598F. doi:10.1126/science.1749933. PMID 1749933.
- ↑ Dunbrack, Roland L (August 2002). "Rotamer Libraries in the 21st Century". Current Opinion in Structural Biology. 12 (4): 431—440. doi:10.1016/s0959-440x(02)00344-5. PMID 12163064.
- ↑ "The backrub motion: how protein backbone shrugs when a sidechain dances". Structure. 14 (2): 265—274. Feb 2006. doi:10.1016/j.str.2005.10.007. PMID 16472746.
- ↑ "Accessing protein conformational ensembles using room-temperature X-ray crystallography". Proceedings of the National Academy of Sciences of the United States of America. 108 (39): 16247—16252. Sep 2011. Bibcode:2011PNAS..10816247F. doi:10.1073/pnas.1111325108. PMID 21918110.
- ↑ "Terahertz underdamped vibrational motion governs protein-ligand binding in solution". Nature Communications. 5: 3999. June 2014. Bibcode:2014NatCo...5.3999T. doi:10.1038/ncomms4999. PMID 24893252.
- ↑ "Dynamic regimes and correlated structural dynamics in native and denatured alpha-lactalbumin". Journal of Molecular Biology. 312 (4): 865—873. Sep 2001. doi:10.1006/jmbi.2001.5006. PMID 11575938.
- ↑ 1 2 "Computational study of conformational changes in human 3-hydroxy-3-methylglutaryl coenzyme reductase induced by substrate binding". Journal of Biomolecular Structure & Dynamics. 37 (16): 4374—4383. October 2019. doi:10.1080/07391102.2018.1549508. PMID 30470158.
- ↑ "Conformational changes in protein loops and helices induced by post-translational phosphorylation". PLOS Computational Biology. 2 (4): e32. April 2006. Bibcode:2006PLSCB...2...32G. doi:10.1371/journal.pcbi.0020032. PMID 16628247.
{{cite journal}}: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (ссылка) - ↑ "Activation of nanoscale allosteric protein domain motion revealed by neutron spin echo spectroscopy". Biophysical Journal. 99 (10): 3473—3482. Nov 2010. Bibcode:2010BpJ....99.3473F. doi:10.1016/j.bpj.2010.09.058. PMID 21081097.
- ↑ "Coupled protein domain motion in Taq polymerase revealed by neutron spin-echo spectroscopy" (PDF). Proceedings of the National Academy of Sciences of the United States of America. 102 (49): 17646—17651. Dec 2005. Bibcode:2005PNAS..10217646B. doi:10.1073/pnas.0503388102. PMID 16306270. Архивировано (PDF) 27 апреля 2021. Дата обращения: 23 августа 2021.
- ↑ 1 2 "Coarse-grained description of protein internal dynamics: an optimal strategy for decomposing proteins in rigid subunits". Biophysical Journal. 96 (12): 4993—5002. Jun 2009. Bibcode:2009BpJ....96.4993P. doi:10.1016/j.bpj.2009.03.051. PMID 19527659.
- ↑ "LSD1/CoREST is an allosteric nanoscale clamp regulated by H3-histone-tail molecular recognition". Proceedings of the National Academy of Sciences of the United States of America. 109 (31): 12509—14. Jul 2012. Bibcode:2012PNAS..10912509B. doi:10.1073/pnas.1207892109. PMID 22802671.
- ↑ ABC Transporters in Microorganisms. — ISBN 978-1-904455-49-3.
- ↑ "At the dawn of the 21st century: Is dynamics the missing link for understanding enzyme catalysis?". Proteins. 78 (6): 1339—75. May 2010. doi:10.1002/prot.22654. PMID 20099310.
- ↑ Mechanics of motor proteins and the cytoskeleton. — 2001. — ISBN 9780878933334.
- ↑ "Controllable Activation of Nanoscale Dynamics in a Disordered Protein Alters Binding Kinetics". Journal of Molecular Biology. 429 (7): 987—998. Apr 2017. doi:10.1016/j.jmb.2017.03.003. PMID 28285124.
- ↑ 1 2 "Structural mechanisms for domain movements in proteins". Biochemistry. 33 (22): 6739—49. Jun 1994. doi:10.1021/bi00188a001. PMID 8204609.
- ↑ "Alpha-catenin structure and nanoscale dynamics in solution and in complex with F-actin". Biophysical Journal. 115 (4): 642—654. Aug 21, 2018. Bibcode:2018BpJ...115..642N. doi:10.1016/j.bpj.2018.07.005. PMID 30037495.
- ↑ Biochemistry. — 2011. — ISBN 9780470570951.
- ↑ "Swiveling-domain mechanism for enzymatic phosphotransfer between remote reaction sites". Proceedings of the National Academy of Sciences of the United States of America. 93 (7): 2652—7. Apr 1996. Bibcode:1996PNAS...93.2652H. doi:10.1073/pnas.93.7.2652. PMID 8610096.
- ↑ "Structural domains in proteins and their role in the dynamics of protein function". Progress in Biophysics and Molecular Biology. 42 (1): 21—78. 1983. doi:10.1016/0079-6107(83)90003-2. PMID 6353481.
- ↑ 1 2 "Structural principles governing domain motions in proteins". Proteins. 36 (4): 425—35. Sep 1999. doi:10.1002/(SICI)1097-0134(19990901)36:4<425::AID-PROT6>3.0.CO;2-S. PMID 10450084.
- ↑ "Target enzyme recognition by calmodulin: 2.4 A structure of a calmodulin-peptide complex". Science. 257 (5074): 1251—1255. Aug 1992. Bibcode:1992Sci...257.1251M. doi:10.1126/science.1519061. PMID 1519061.
- ↑ "Solution structure of a calmodulin-target peptide complex by multidimensional NMR". Science. 256 (5057): 632—638. May 1992. Bibcode:1992Sci...256..632I. doi:10.1126/science.1585175. PMID 1585175.
- ↑ "Protein dynamism and evolvability". Science. 324 (5924): 203—207. Apr 2009. Bibcode:2009Sci...324..203T. doi:10.1126/science.1169375. PMID 19359577.
- ↑ "Intrinsically unstructured proteins and their functions". Nature Reviews Molecular Cell Biology. 6 (3): 197—208. Mar 2005. doi:10.1038/nrm1589. PMID 15738986.













