Происхождение четвероногих — эволюционный процесс, в результате которого образовалась группа четвероногих животных, приспособленных к жизни на суше.

Происхождение рептилий — один из важных вопросов в теории эволюции, процесс, в результате которого появились первые животные, относящиеся к классу Пресмыкающиеся.

Эдафозавры (лат. Edaphosauridae) — семейство растительноядных пеликозавров. Обитали в Северной Америке и Европе в конце каменноугольного — начале пермского периодов. Одни из первых крупных доминирующих растительноядных. Голова маленькая, зубы колышковидные, равного размера. Нёбо и внутренняя сторона нижней челюсти покрыты дополнительными зубами. Есть глубокая нижняя вырезка скуловой дуги. Возможно, имелись мягкие щёки, позволявшие удерживать пищу во рту. Конечности короткие, тело массивное, широкая грудная клетка. У всех эдафозавров был «парус», образованный остистыми отростками спинных позвонков. На отростках имелись поперечные выросты, верхушки нескольких передних отростков часто расширены. Вероятно, как и у сфенакодонтов, «парус» служил для терморегуляции, поперечные отростки могли увеличивать его площадь. Кости «паруса» были полыми. Нельзя также исключить, что у основания «паруса» могли откладываться запасы жира.

Казеиды (лат. Caseidae) — семейство растительноядных пеликозавров, существовавших в течение всего пермского периода. Появились в конце нижнепермской эпохи, дожили до середины перми.

Темноспонди́льные, или расчленённопозвонковые (лат. Temnospondyli, от др.-греч. τέμνειν — резать и σπόνδυλος — позвонок) — в высшей степени разнообразная группа вымерших животных, включающая от маленьких до гигантских примитивных земноводных. Название группы отсылает к тому, что каждый позвонок темноспондильных разделён на несколько частей.

Proterogyrinus (лат.) — род примитивных рептилиоморф из семейства Proterogyrinidae. Жили во времена карбона.

Уинтатерии — род вымерших млекопитающих из отряда диноцерат. Один из самых крупных представителей отряда. Наиболее характерная черта — три пары рогоподобных выростов на крыше черепа, более развитые у самцов. Выросты были покрыты кожей, как оссиконы жирафов.

Эдопоиды. Разнообразная группа пермокарбоновых примитивных темноспондилов. Исходно — полуназемные формы, позднее появились преимущественно водные. Выделяют следующие надсемейства:

Трематопиды — семейство раннепермских темноспондилов. Входят в состав надсемейства диссорофоид (Dissorophoidea), куда относятся самые наземные из темноспондилов.

Капториниды (лат. Captorhinidae) — семейство примитивных пресмыкающихся верхнего палеозоя. Традиционно относились к так называемым анапсидным рептилиям, лишённым височных окон. По последним исследованиям, стоят ближе к диапсидным рептилиям, чем к другим анапсидам. С капторинидами сближают также древнейших известных амниот — семейство Protorothyrididae, самые известные представители которых: гилономус и палеотирис. По последним данным, проторотиридиды находятся в основании ствола диапсид.

Диадекты — род примитивных четвероногих, живший с верхнего карбона по нижнюю пермь, 299—271 млн лет назад. Описаны Э. Д. Коупом в 1878 году. Считаются либо относительно продвинутыми рептилиоморфами, либо примитивными амниотами (парарептилии). В классе завропсид входят в отряд котилозавров, либо выделяются в отряд/подотряд диадектоморфов, более примитивных, чем прочие амниоты.

Э́риопс — представитель темноспондилов позднекаменноугольной — раннепермской эпохи. Типовой род семейства Eryopidae, один из самых известных темноспондилов.

Бафетиды (лат. Baphetidae) — семейство примитивных стегоцефалов, живших в каменноугольном периоде. Одни из первых ископаемых «земноводных», ставших известными науке — впервые описаны У. Даусоном в 1850 году. В то же время, это одни из самых древних последевонских стегоцефалов. Известны практически только по ископаемым черепам. Черепа обычно низкие, по структуре костей близки к примитивным темноспондилам. Желобки боковой линии обычно присутствуют. Орбита расширена вниз и вперёд, образуя подобие «замочной скважины», расширение превосходит орбиту по размерам. Есть довольно значительно развитая «ушная» вырезка. Нёбо закрытое, как у антракозавров, строение таблитчатых костей сходно с таковым у темноспондилов. Есть шов между щекой и крышей черепа, слуховая капсула доходит до таблитчатой кости. Зубы лабиринтодонтные. Развиты крупные клыки на нёбе, краевые зубы также крупные, двухрядные на нижней челюсти. Нижняя челюсть высокая, выше чем верхняя челюсть. По-видимому, рыбоядные хищники, постоянноводные. Назначение расширения глазницы неясно. Высказываются следующие предположения: в «окне» могло быть крепление мышц, какие-либо железы, электрические органы.

Диссорофиды — семейство темноспондилов позднего карбона — перми. Наиболее наземные из всех темноспондилов. Скелет массивный, тело короткое, конечности длинные, сильные. Голова огромная относительно размеров тела. Череп высокий и широкий, огромные глазницы, крупное теменное отверстие. Огромные ушные вырезки, судя по строению окружающей кости, содержали барабанную перепонку. Затылочный мыщелок парный. Челюстное сочленение на уровне затылочного. Скульптура черепа гребнистая. Края орбит и теменного отверстия окружены костными валиками, что предполагает толстую кожу. У большинства известен дермальный панцирь на спине из парных рядов костных пластин, приросших к остистым отросткам позвонков, у какопса пластины лежали в два слоя. Кожа, по-видимому, мягкая, следов чешуи не выявлено. Хвост короткий. Вероятно, прибрежные амфибиотические и полуназемные хищники. Есть свидетельства, что крупные диссорофиды могли питаться и падалью.

Офиакодонты — группа примитивных хищных пеликозавров. Древнейшие из пеликозавров, фрагментарные остатки известны уже с середины каменноугольного периода. Единственное семейство — Ophiacodontidae. Отличаются длинным, у многих — относительно высоким черепом, у поздних форм череп очень крупный относительно размеров тела. Зубы многочисленные, некрупные, «клыки» выражены слабо. Отогнутой пластинки угловой кости нет. Плечевой пояс массивный, кости запястья и плюсны слабо окостеневшие, у поздних представителей когти могли отсутствовать. Некоторые виды, вероятно, были рыбоядными водными хищниками, более примитивные представители могли охотиться на суше на крупных насекомых и мелких позвоночных.

Архерия — род антракозавров из семейства Archeriidae. Жили во времена позднего карбона — ранней перми на территории современных США.

Лэдлерия — раннетриасовый темноспондил. Животное, из группы трематозавров. Череп очень плоский, в виде равнобедренного треугольника, с острой мордой. Зубы нижней челюсти крупнее, чем на верхней, есть нижнечелюстные «клыки». Верхнечелюстная кость отделена от хоаны шовным соединением между сошником и нёбом. Более 8 нёбных зубов. Орбиты некрупные, вблизи середины черепа, сближены, направлены вверх. Есть боковой вырост квадратоскуловой кости. Стремя массивное, барабанной перепонки не было. Орнамент черепа в виде ямок и гребней. Сохраняются желобки «боковой линии» на передне-верхней поверхности черепа. Тело уплощённое, массивное, покрытое мелкими многоугольными костными пластинками. Конечности плохо известны, хвост, вероятно, короткий. Длина черепа до 10 см, общая длина 30—40 см. Происходит из раннего триаса Южной Африки. Единственный вид — L. gracilis, описан Дж. Китчингом в 1958 году. Близкий род Uruyiella с единственным видом U. liminea описан в 2007 году из низов триаса Уругвая на основании фрагментов черепа. Образ жизни лэдлерий малопонятен. Иногда их считают наземными животными, но против этого свидетельствует наличие органов «боковой линии». Вероятно, всё же эти родичи плагиозавров были в основном водными малоподвижными охотниками за некрупной добычей.
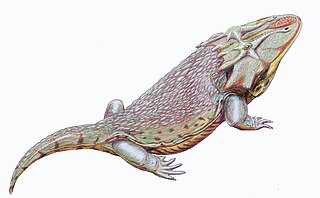
Затрахидиды или затрахеиды — семейство темноспондилов из ранней перми.

Евгенеодонтообразные (лат. Eugeneodontiformes) — отряд вымерших, преимущественно верхнепалеозойских, хрящевых рыб из подкласса цельноголовых. Близки к общим предкам химер и акул, иногда считаются «настоящими» акулами и сближаются с кладоселяхиями.
Диадектоморфы (лат. Diadectomorpha) — отряд крупных рептилиоморф из карбона и нижней перми Еврамерики и верхней перми Азии, очень близкий к родоначальникам амниот. Включают крупные плотоядные и еще более крупные травоядные формы, некоторые — полуводные, другие — полностью наземные. Диадектоморфы, видимо, возникли в верхнем миссисипии, но широкое распространение получили после коллапса каменноугольных лесов и процветали во времена верхнего пенсильвания и нижней перми. Несмотря на широкое распространение и видовое разнообразие диадектоморфы не были доминантами. Часто их описывают по фрагментарным останкам. В Еврамерике полностью вымирают на границе нижней и средней перми, из средней перми Татарстана известен один образец, который, возможно, относится к диадектам. Позднейший из найденных образцов Alveusdectes fenestratus происходит из верхней перми Цзиюань (Китай).
![Реконструкция скелета и модель диплокаулуса в Денверском музее природы и науки[англ.]](https://upload.wikimedia.org/wikipedia/commons/thumb/c/c1/Diplocaulus_-_skeleton_and_model.jpg/275px-Diplocaulus_-_skeleton_and_model.jpg)

