Доминантность
Домина́нтность, или домини́рование, — форма взаимоотношений между аллелями одного гена, при которой один из них (доминантный) подавляет (маскирует) проявление другого (рецессивного) и таким образом определяет проявление признака как у доминантных гомозигот, так и у гетерозигот.
Рецесси́вный аллель — вариант гена, действие которого на фенотип не проявляется в присутствии доминантного аллеля. Рецессивный аллель способен обеспечить проявление определяемого им признака только в том случае, если находится в гомозиготном состоянии (в паре с таким же рецессивным аллелем).
Полное доминирование
При полном доминировании фенотип гетерозиготы не отличается от фенотипа доминантной гомозиготы по данному аллелю. Видимо, в чистом виде полное доминирование встречается крайне редко или не встречается вовсе. Например, люди, гетерозиготные по гену гемофилии А (сцепленный с Х-хромосомой рецессивный аллель), имеют половинное количество нормального фактора свёртывания по сравнению с гомозиготными по нормальному аллелю людьми, и активность фактора свёртывания VIII у них в среднем вдвое ниже, чем у здоровых людей. В то же время у здоровых людей активность этого фактора варьирует от 40 до 300 % по сравнению со средней для популяции. Поэтому наблюдается значительное перекрывание признаков у здоровых и носителей-гетерозигот. При фенилкетонурии (аутосомно-рецессивный признак) гетерозиготы обычно считаются здоровыми, однако активность печёночного фермента фенилаланин-4-гидроксилазы у них вдвое ниже нормы, а содержание фенилаланина в клетках повышено, что, по некоторым данным, приводит к снижению IQ и повышенному риску развития некоторых психотических расстройств.
Неполное доминирование
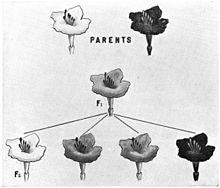
При неполном доминировании гетерозиготы имеют фенотип, промежуточный между фенотипами доминантной и рецессивной гомозиготы. Например, при скрещивании чистых линий львиного зева и многих других видов цветковых растений с пурпурными и белыми цветками особи первого поколения имеют розовые цветки. При скрещивании чистых линий андалузских кур чёрной и белой окраски в первом поколении рождаются куры серой окраски. На молекулярном уровне самым простым объяснением неполного доминирования может быть как раз двукратное снижение активности фермента или другого белка (если доминантный аллель даёт функциональный белок, а рецессивный — дефектный). Например, за белую окраску может отвечать дефектный аллель, который даёт неактивный фермент, а за красную — нормальный аллель, который даёт фермент, производящий красный пигмент. При половинной активности этого фермента у гетерозигот количество красного пигмента снижается вдвое, и окраска розовая. Могут существовать и другие механизмы неполного доминирования.
При неполном доминировании во втором поколении моногибридного скрещивания наблюдается одинаковое расщепление по генотипу и фенотипу в соотношении 1:2:1.
В некоторых источниках неполное доминирование характеризуют как такой тип взаимодействия аллелей, когда признак у гибридов F1 занимает не среднее положение, а отклоняется в сторону родителя с доминирующим признаком. Полностью же средний вариант (как, например, приведённый выше пример наследования окраски цветков) относят к промежуточному характеру наследования, то есть отсутствию доминирования[1].
Кодоминирование

При кодоминировании, в отличие от неполного доминирования, у гетерозигот признаки, за которые отвечает каждый из аллелей, проявляются одновременно и в полной мере. Типичный пример кодоминирования — наследование групп крови системы АВ0 у человека. Всё потомство людей с генотипами АА (вторая группа) и ВВ (третья группа) будет иметь генотип АВ (четвёртая группа). Их фенотип не является промежуточным между фенотипами родителей, так как на поверхности эритроцитов присутствуют оба агглютиногена (А и В). При кодоминировании назвать один из аллелей доминантным, а другой — рецессивным нельзя, эти понятия теряют смысл: оба аллеля в равной степени влияют на фенотип. На уровне РНК и белковых продуктов генов, видимо, подавляющее большинство случаев аллельных взаимодействий генов — это кодоминирование, ведь каждый из двух аллелей у гетерозигот обычно кодирует РНК и/или белковый продукт, и оба белка или РНК присутствуют в организме.
Другие варианты доминирования
- Сверхдоминирование — более сильное проявление признака у гетерозиготной особи, чем у любой гомозиготной. На этом типе аллельного взаимодействия основано явление гетерозиса (превосходство над родителями по жизнеспособности, энергии роста, плодовитости, продуктивности)[1].
- Доминирование, связанное с полом, происходит тогда, когда одна и та же аллель у самцов проявляется как доминантная, а у самок — как рецессивная. Например, у овцематок доминирует комолость (R), а у баранов — рогатость (R1)[1].
Относительный характер доминирования
Как уже отмечалось выше, характер доминирования зависит от уровня анализа признака. Рассмотрим это на примере серповидно-клеточной анемии. Гетерозиготные носители гена гемоглобина S (AS) на уровне моря имеют нормальную форму эритроцитов и нормальную концентрацию гемоглобина в крови (полное доминирование А над S). На больших высотах (более 2500—3000 м) у гетерозигот концентрация гемоглобина понижена (хотя и намного выше, чем у больных), появляются эритроциты серповидной формы (неполное доминирование А над S). Этот пример показывает, что доминантность может зависеть от условий. Гетерозиготы AS и гомозиготы SS обладают примерно одинаковой устойчивостью к малярии, гомозиготы АА подвержены малярии в большей степени. По данному проявлению ген S доминирует над А. Наконец, в эритроцитах носителей АS в равных количествах присутствуют оба варианта бета-глобиновых цепей — нормальный А и мутантный S (то есть наблюдается кодоминирование).
Молекулярные механизмы
Молекулярные основы доминирования были неизвестны Менделю. В настоящее время ясно, что локус, соответствующий определённому гену, состоит из длинных последовательностей, включающих сотни и тысячи нуклеотидов ДНК. Центральная догма молекулярной биологии гласит, что ДНК → РНК → белок, то есть ДНК транскрибируется в мРНК, а мРНК транслируется в белок. В этом процессе различные аллели могут транскрибироваться или не транскрибироваться, а будучи затранскрибированными, транслироваться в различные формы одного и того же белка — изоформы. Часто белки функционируют как ферменты, катализирующие химические реакции в клетке, которые прямо или косвенно определяют фенотип. У любого диплоидного организма аллели, соответствующие одному локусу, являются либо одинаковыми (у гомозигот), либо разными (у гетерозигот). Даже если на уровне последовательностей ДНК аллели различны, то их белки могут быть идентичными. В отсутствие различий между белковыми продуктами невозможно сказать, какой из аллелей доминирует (в этом случае имеет место кодоминирование). Даже если два белковых продукта слегка отличны друг от друга, они, вероятно, дают одинаковый фенотип и могут осуществлять одинаковые ферментативные реакции (если они являются ферментами). В этом случае также невозможно сказать, какой из аллелей доминирует.
Доминирование, как правило, возникает, когда один из аллелей является нефункциональным на молекулярном уровне, то есть не транскрибируется или даёт нефункциональный белковый продукт. Это может быть результатом мутации, изменяющей последовательность ДНК аллеля. У гомозиготы по нефункциональным аллелям, как правило, проявляется характерный фенотип из-за отсутствия определённого белка. Например, у людей и других животных непигментированная кожа альбиносов проявляются из-за гомозиготности по аллелю, препятствующему синтезу кожного пигмента меланина. Важно понимать, что рецессивность определяется у аллеля не по отсутствию какой-либо функции: у гетерозигот это является результатом взаимодействия с альтернативным аллелем. Возможны три основных типа таких взаимодействий:
- В типичном случае единичный функциональный аллель даёт достаточно белка, чтобы получить фенотип, идентичный фенотипу гомозиготы по функциональному аллелю. Это называется гаплодостаточностью (англ. haplosufficiency). Например, если принять количество фермента, производимого функциональной гетерозиготой, за 100%, то каждый из функциональных аллелей будет ответственна за выработку 50% общего количества фермента. Единственный функциональный аллель гетерозиготы даёт 50% фермента, и этого достаточно для поддержания нормального фенотипа. Если гетерозигота и гомозигота по функциональному аллелю имеют одинаковый фенотип, то функциональный аллель доминирует над нефункциональным. Так происходит с геном альбинизма: гетерозигота производит количество фермента, которого достаточно для образования предшественника меланина, и особь имеет нормальную пигментацию.
- Реже наличие единственного функционального аллеля не обеспечивает нормальный фенотип, однако его дефектность выражена не так ярко, как у гомозиготы по нефункциональным аллелям. Это происходит тогда, когда функциональный аллель не является гаплодостаточным. Обычно к этим случаям относят понятия гаплонедостаточности и неполного доминирования. Промежуточный вариант этого взаимодействия имеет место тогда, когда гетерозигота имеет фенотип, промежуточный между двумя гомозиготами. В зависимости от того, к какой из гомозигот ближе вариант признака гетерозиготы, говорят о неполном доминировании одной аллели над другой. Примером такого взаимодействия может служить описанный выше случай с гемоглобином человека.
- Редко единственная функциональная аллель гетерозиготы даёт неполноценный генный продукт, и её фенотип схож с фенотипом гомозиготы по нефункциональным аллелям. Такие случаи гаплонедостаточности крайне необычны. В этих случаях нефункциональная аллель доминирует над функциональной. Такая ситуация может происходить тогда, когда нефункциональная аллель даёт дефектный белок, который подавляет функцию белка, образуемого нормальной аллелью. Дефектный белок «доминирует» над стандартным, и фенотип гетерозиготы более походит на фенотип гомозиготы по дефектным аллелям. Следует обратить внимание на то, что доминантными часто некорректно называют дефектные аллели, вызываемый которыми в гомозиготном состоянии фенотип не изучен, однако в сочетании с нормальным аллелем они дают характерный фенотип. Этот феномен происходит при некоторых генетических заболеваниях, вызванных тринуклеотидными повторами, например, болезни Хантингтона.
Эволюция доминантности
Новые мутации могут, конечно, сразу обладать доминантным проявлением в фенотипе диплоидных особей, но вероятность выживания мутантов вообще невелика, и поэтому преимущественно сохраняются именно рецессивные мутации. Впоследствии, если при каких-либо изменениях внешних условий новый признак окажется благоприятным, обусловливающий его мутантный аллель может вторично приобрести доминантное фенотипическое выражение (следует подчеркнуть, что доминантны и рецессивны, собственно говоря, не сами аллели, а их проявления в фенотипе). Переход аллеля от рецессивного к доминантному состоянию может быть обусловлен различными механизмами, действующими на разных уровнях преобразований наследственной информации в онтогенезе. Генетически такой переход может быть достигнут через отбор особых генов-модификаторов, влияющих на фенотипическое проявление мутантного аллеля (гипотеза Р. Фишера), или же через отбор аллелей с большей физиологической активностью (обеспечивающих более интенсивный синтез ферментов), чем первоначальный рецессивный вариант (гипотезы С. Райта и Д. Холдейна). В сущности, эти гипотезы не исключают, а взаимно дополняют друг друга, и эволюция доминантности может происходить путём отбора малых мутаций как структурных генов, так и генов-модификаторов.
Так или иначе, степень доминантности фенотипического проявления аллелей может эволюционировать, повышаясь под контролем отбора, если данный аллель становится благоприятным для его носителя при изменениях внешних условий. Примером этого может служить повышение доминантности аллеля, контролирующего тёмную окраску бабочек берёзовой пяденицы (Biston betu-laria), которое, по некоторым данным, произошло в течение последних ста лет в индустриальных районах Европы (явление, получившее название «индустриальный меланизм») (Н. Н. Иорданский «Эволюция жизни»).
См. также
- Аллель
- Гетерозигота
- Гомозигота
- Доминантный признак
- Рецессивный признак
- Законы Менделя
- Взаимодействие неаллельных генов
Примечания
- ↑ 1 2 3 Максимов Г. В., Василенко В. Н., Кононенко О. И., Максимов А. Г., Максимов В. Г. Сборник задач по генетике. — М.: Вузовская книга, 2010. — С. 15—20. — 144 с. — 300 экз. — ISBN 978-5-9502-0420-3.
Ссылки
- «OMIM Entry — *612349 — Phenylalanine hydroxylase; PAH». OMIM.org.
- Cat Coat Color. Vgl.ucdavis.edu. doi:10.1111/j.1365-2052.2009. Дата обращения: 2 ноября 2011. Архивировано 19 мая 2012 года.
- Carr, Steven M. Extensions to Mendelian Analysis. Mun.ca. Memorial University of Newfoundland. Дата обращения: 2 ноября 2011. Архивировано 19 мая 2012 года.







