Европазавр
| † Европазавр | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
Домен: Эукариоты Царство: Животные Подцарство: Эуметазои Без ранга: Двусторонне-симметричные Без ранга: Вторичноротые Тип: Хордовые Подтип: Позвоночные Инфратип: Челюстноротые Надкласс: Четвероногие Клада: Амниоты Клада: Завропсиды Клада: Архозавры Клада: Авеметатарзалии Клада: Динозавроморфы Клада: Динозавры Клада: Ящеротазовые Подотряд: † Завроподоморфы Клада: † Massopoda Клада: † Sauropodiformes Инфраотряд: † Завроподы Клада: † Макронарии Клада: † Camarasauromorpha Род: † Европазавр | |||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Europasaurus Mateus et al. in Sander et al., 2006[1] | |||||||||||||||||||||||||||||||||||||||||
| Единственный вид | |||||||||||||||||||||||||||||||||||||||||
Mateus et al. in Sander et al., 2006 | |||||||||||||||||||||||||||||||||||||||||
| Геохронология вымер 154 млн лет
◄ Наше время◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание◄ Массовоепермское вымирание ◄ Девонское вымирание◄ Ордовикско-силурийское вымирание◄ Кембрийский взрыв | |||||||||||||||||||||||||||||||||||||||||


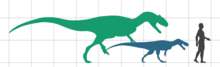
Европазавр[2] (лат. Europasaurus, буквально — ящер из Европы) — род завроподных динозавров из клады макронарий, включающий единственный вид — Europasaurus holgeri. Известен по ископаемым остаткам из отложений формации Зюнтель[англ.] на севере Германии, датируемых средним киммериджем (верхняя юра)[3]; абсолютный возраст материала составляет 154 млн лет[4]. Является примером островной карликовости[1]. Европазавр достигал около 6 м в длину, что делает его одним из самых маленьких завропод[5] (наряду с мадьярозавром[6]).
Согласно разным интерпретациям, занимает либо базальное положение в пределах клады Camarasauromorpha[1][7][8], либо относится к одной из подгрупп этой клады, к брахиозавридам, в пределах которых также занимает базальное положение[9][10][11][12].
История открытия

В 1998 году в действующем карьере на холме Лангенберг[англ.], между общинами Окер[англ.], Харлингероде[англ.] и Гёттингероде[англ.] в Германии, частный коллекционер окаменелостей Хольгер Людтке обнаружил изолированный зуб завропода. Меловой карьер Лангенберг действует более века; горные породы добываются взрывными работами и в основном перерабатываются в удобрения. Карьер обнажает почти непрерывную последовательность карбонатных пород толщиной в 203 м[4], относящихся к формации Зюнтель, возраст которых колеблется от нижнего оксфорда до верхнего киммериджа (верхняя юра). Породы были отложены в мелководном море глубиной не более чем в 30 м. Слои, обнажённые в карьере, ориентированы почти вертикально и слегка опрокинуты, что является результатом подъёма соседних гор Гарц в течение раннемеловой эпохи. Карьер широко известен как классическое обнажение среди геологов, хорошо изучен и десятилетиями посещается студентами-геологами. Хотя он богат окаменелостями морских беспозвоночных, окаменелости наземных животных здесь не столь часты; обнаруженный Людтке зуб стал первым образцом завроподового динозавра из юрских отложений северной Германии[13].
После того как было найдено больше ископаемого материала, включая кости, в апреле 1999 года начались раскопки костеносного слоя, организованные местным обществом частных коллекционеров окаменелостей. Хотя оператор карьера был настроен на сотрудничество, раскопки осложнились почти вертикальной ориентацией слоёв, которая ограничивала доступ, а также продолжавшейся разработкой карьера. Ископаемые остатки пришлось собирать из блоков, образовавшихся в результате взрывных работ. В связи с этим точное происхождение костного материала оставалось неясным, хотя позже его удалось проследить до конкретного пласта (пласт №83)[13][14].
Окаменелости препарировали и хранили в Парке динозавров Мюнхегаген[англ.], частном музее динозавров под открытым небом, расположенном неподалёку от Ганновера. Из-за очень хорошей сохранности костей укрепляющие вещества приходилось применять лишь изредка, и препарирование проходило сравнительно быстро, так как кости легко отделялись от породы. Кости простой формы иногда препарировали менее чем за час, тогда как препарация крестца заняла три недели. К январю 2001 года препараторы извлекли из породы около 200 отдельных костей позвоночных. К этому моменту наибольшая концентрация костей была обнаружена в блоке размером 70 х 70 см, который содержал около 20 костей[14]. К январю 2002 года при препарации ещё большего блока был обнаружен неполный череп завропода — первый, обнаруженный в Европе. Перед полным извлечением костей исследователи сделали кремниевый слепок блока для фиксации точного положения отдельных костей в трёхмерном виде[15].
Часть материала европазавра оказалась повреждена или уничтожена в результате поджога в ночь с 4 на 5 октября 2003 года. Пожар уничтожил лабораторию и выставочный зал Парка динозавров Мюнхегаген, в результате чего было утрачено 106 костей, что составляет 15% костей, отпрепарированных к тому времени. Кроме того, огонь затронул большую часть ещё не препарировавшихся блоков, а противопожарная вода, попав на нагретую породу, ещё больше раскрошила её[16]. К уничтоженным образцам относится DFMMh/FV 100, включавший наиболее хорошо сохранившийся позвоночник и единственный полный таз европазавра[7][16].

В 2006 году новый таксон был научно описан под названием Europasaurus holgeri. Родовое название означает «ящер из Европы», а видовое дано в честь Хольгера Людтке, открывшего первые окаменелости животного[1]. Учитывая сравнительно небольшой размер костей, первоначально предполагалось, что они принадлежат молодым особям[13]. Однако в публикации 2006 года учёные установили, что большинство обнаруженных особей были взрослыми, а сам европазавр являлся островным карликом[1][13]. К моменту открытия количество обнаруженных костей достигло примерно 650; известны ископаемые остатки, в том числе сочленённые кости, обнаруженные на площади 60—80 м²[4]. Из этих образцов был отобран голотип: разрозненный, но ассоциированный образец (DFMMh/FV 291). Голотип включает в себя несколько костей черепа (предчелюстная кость, верхняя челюсть и квадратно-скуловая кость), неполную черепную коробку, несколько костей нижней челюсти (зубные, угловые и надугловые), большое количество зубов, шейные позвонки, крестцовые позвонки и ребра от шеи и туловища. По меньшей мере 10 других особей были определены как принадлежащие европазавру на основе наличия перекрывающегося материала[1].
Летом 2012 года началась крупномасштабная раскопочная кампания, целью которой было извлечь кости европазавра не только из утерянных блоков, но и непосредственно из слоя породы. Для доступа к костесодержащему слою потребовалось удалить около 600 тонн породы с помощью экскаваторов и колесных погрузчиков, при этом постоянно откачивая воду из основания карьера. Раскопки продолжались в течение весны и лета 2013 года. Эта кампания привела к открытию окаменелостей рыб, черепах и крокодиломорф, а также приобретению ценной информации о костеносном слое, но дополнительных костей европазавра обнаружить не удалось. К 2014 году было отпрепарировано около 1300 костей позвоночных из слоя №83, бо́льшая часть которых принадлежит европазавру; по оценкам, ещё 3000 костей ожидают препарации. По челюстным костям было идентифицировано не менее 20 особей[13].
Описание
Предполагается, что европазавр достигал всего около 6,2 м в длину во взрослом возрасте, что выделяет его на фоне большинства других завропод. Эта оценка была произведена путём масштабирования неполной бедренной кости европазавра до размеров почти полного образца камаразавра. Известны и более молодые особи европазавра, достигавшие в длину от 1,75 м (самая молодая известная особь) до 3,7 м. Масса европазавра оценивается примерно в 800 кг. Титанозавр мадьярозавр (Magyarosaurus) имел сходные размеры и, по оценкам, весил приблизительно 900 кг. Другой небольшой титанозавр, Neuquensaurus, был крупнее обоих этих животных, достигая 9 м в длину при массе в 3,2 т[6].
Отличительные черты

Авторы описания европазавра выделили следующие аутапоморфии, по которым его можно отличить от ближайших родственников[1]:
- Носовой отросток предчелюстной кости изгибается вперёд и выступает вверх;
- На верхней поверхности тел шейных позвонков присутствует выемка;
- Акромион заметно выступает назад;
- Широта таранной кости в два раза превышает высоту.
Marpmann et al. (2014) пришли к выводу, что форма носового отростка является артефактом сохранения, а выемка на верхней поверхности тел шейных позвонков является широко распространённой чертой, что позволило им исключить первые два признака из списка аутапоморфий. Кроме того, были добавлены две новых аутапоморфии[17]:
- Скуловая кость принимает большое участие в формировании глазницы с вентральной стороны;
- Оптическое отверстие одно.
От камаразавра европазавр отличается менее коротким задним фланцем заглазничной кости, более коротким сочленением между носовой и лобной костями черепа, а также другой формой теменной кости (прямоугольной) и невральными отростками позвонков, которые не расщепляются перед тазом. Авторы описания провели также сравнение с костями жираффатитана[18] (на том момент считавшимся синонимом брахиозавра) и установили, что европазавр, в отличие от жираффатитана, обладает более короткой мордой, контактом между квадратно-скуловой и чешуйчатой костями, а также отличается плечевой костью, имеющей уплощённые и выравненные проксимальную и дистальную поверхности. Наконец, было проведено краткое сравнение с потенциальным брахиозавридом Lusotitan[англ.], который обладает другой формой подвздошной и таранной костей, и Duriatitan[19] (на тот момент Cetiosaurus humerocristatus), у которого не столь выражен дельтопекторальный гребень на плечевой кости[1].
Классификация
Авторы описания европазавра классифицировали его как базальную макронарию, не входящую в семейство брахиозаврид (Brachiosauridae) или в кладу Somphospondyli[20] (в оригинальной статье используется название Titanosauromorpha). Это указывает на то, что карликовость таксона стала результатом его эволюции, а не была унаследована от предков[1]. Были проанализированы три матрицы, в которые авторы описания добавили европазавра: по Wilson (2002[21]), по Upchurch (1998[22]) и по Upchurch et al. (2004[23]). Все анализы дали сходные филогенетические деревья, в которых европазавр более продвинут, чем камаразавр, но находится за пределами брахиозаврид и Somphospondyli[4]:
Sander et al., 2006[4]
| Carballido & Sander, 2013[7]
|
При описании позвонков европазавра Carballido & Sander (2013) провели ещё один филогенетический анализ. Матрица данных была расширена путём включения таких таксонов завропод как Bellusaurus, Cedarosaurus[англ.] и Tapuiasaurus. Кроме того, род Brachiosaurus был разделен на истинного брахиозавра Brachiosaurus altithorax и жираффатитана Giraffatitan brancai (ранее B. brancai), в соответствии с ревизией Taylor[англ.] (2009[18]). По результатам этого анализа европазавр занял на кладограмме аналогичное положение: как базальный представитель клады Camarasauromorpha, более близкий к титанозаврам, чем камаразавр. Euhelopus, Tehuelchesaurus[англ.], Tastavinsaurus и Galvesaurus[англ.] заняли на полученном дереве промежуточную позицию между европазавром и Titanosauriformes (брахиозавридами и Somphospondyli)[7].
В качестве брахиозаврида

Филогенетический анализ Titanosauriformes, проведённый D'Emic (2012), указывает на принадлежность европазавра к брахиозавридам. При этом некоторые таксоны, часто классифицировавшиеся как брахиозавриды (Sauroposeidon и Qiaowanlong), попали в кладу Somphospondyli. Д'Эмик отметил, что европазавр пока не может быть уверенно включён в число брахиозаврид, поскольку невозможно подтвердить или опровергнуть наличие у него некоторых синапоморфий. Это связано с тем, что ряд костей, таких как слёзная кость, IV плюсневая кость и хвостовые позвонки, ещё не описаны или обладают недостаточной сохранностью[9]:
D'Emic, 2012[9]
| Royo-Torres et al., 2017[12]
|
Mannion et al. (2013) согласились с D'Emic (2012) в вопросе филогенетического положения европазавра. По результатам проведённого ими анализа Titanosauriformes европазавр оказался в политомии[англ.] с брахиозавром и «французским Bothriospondylus[англ.]» (позже названным Vouivria[англ.]) в составе базальной группы брахиозаврид. Исходя из результатов анализа, далее идёт клада более продвинутых брахиозаврид, состоящая Lusotitan, Giraffatitan, Abydosaurus, Cedarosaurus и Venenosaurus[англ.]. Было обнаружено, что «искривлённые» зубы европазавра являются одной из апоморфий (уникальных признаков) брахиозаврид; если это действительно так, то аналогичные изолированные зубы завропод могут быть уверенно определены как принадлежащие брахиозавридам[10].
Дальнейший филогенетический анализ брахиозаврид D'Emic et al. (2016), основанный на анализе D'Emic (2012), показал очень похожее размещение европазавра внутри брахиозаврид, хотя Sonorasaurus был помещён в кладу с жираффатитаном, а Lusotitan — в политомию с Abydosaurus и Cedarosaurus. Остальное дерево повторяет дерево D'Emic (2012), за исключением того, что брахиозавр оказался свёрнут в политомию с остальными продвинутыми брахиозавридами[24]. Другой анализ, проведённый Mannion et al. (2017), дал аналогичные с анализами D'Emic (2012) и D'Emic et al. (2016) результаты. Согласно результатам этого анализа, европазавр — самый базальный представитель брахиозаврид, следом за которым идёт немного более продвинутая Vouivria. Брахиозавр оказался вне политомии всех остальных брахиозаврид, состоящей из жираффатитана, Abydosaurus, Sonorasaurus, Cedarosaurus и Venenosaurus[11]. Филогения Royo-Torres et al. (2017) разрешила более сложные филогенетические взаимоотношения между брахиозавридами. Помимо базального среди брахиозаврид положения европазавра, было продемонстрировано разделение брахиозаврид на две дочерние клады, одна из которых содержит жираффатитана, Sonorasaurus и Lusotitan, а другая — почти всех остальных брахиозаврид, включая Tastavinsaurus. Эта вторая группа была названа Laurasiformes[англ.] и определена как клада наибольшего объёма, включающая Tastavinsaurus, но не Saltasaurus. Брахиозавр оказался в политомии с обеими кладами брахиозаврид[12].
Carballido et al. (2020), проведя филогенетические анализы на основе матриц данных разных авторов, пришли к выводу о том, европазавр, скорее всего, входит в число базальных Camarasauromorpha; однако авторы использовали непараметрический тест и поэтому предупредили, что пока нельзя полностью исключать принадлежности европазавра к брахиозавридам[8].
Палеобиология
Онтогенез

Путём гистологического анализа палеонтологи установили, что E. holgeri был уникальным карликовым видом, а не детёнышем другого таксона, например, камаразавра. Гистологические данные говорят о том, что образец европазавра DFMMh/FV 009 не имеет признаков старения, таких как следы роста или пластинчатая костная ткань; прижизненная длина животного была оценена в 1,75 м, что делает его самым маленьким среди всех образцов европазавра. Пластинчатая костная ткань является показателем быстрого роста, поэтому скелет, у которого этой ткани нет, вероятно, принадлежит самому молодому известному европазавру. Более крупный образец (DFMMh/FV 291.9), оцененный как достигавший 2 м в длину, обладает значительным количеством пластинчатой ткани, но не имеет каких-либо следов роста, так что, скорее всего, он также представляет собой молодое животное. Следующий по размеру образец (DFMMh/FV 001) демонстрирует наличие следов роста (в частности, кольцеобразных) и, достигая при жизни около 3,5 м в длину, вероятно, умер в подростковом возрасте. DFMMh/FV 495, оцененный в 3,7 м в длину, обладает характерным для взрослых особей строением остеонов, а также кольцеобразными следами роста. Второй по величине описанный образец (DFMMh/FV 153) также имеет признаки роста, причём у него они встречаются ещё чаще. Предположительно, это животное достигало 4,2 м в длину. Неполная бедренная кость (DFMMh/FV 415) может принадлежать самой большой известной особи европазавра, длина которой была оценена в 6,2 м. Эта бедренная кость отличается наличием линий остановленного роста[англ.], указывающих на то, что динозавр умер уже после прекращения роста. Внутри кость частично пластинчатая, что говорит о том, что она перестала расти незадолго до смерти[1][4].
Описанные выше гистологические данные показывают, что небольшой размер разделялся всеми европазаврами вне зависимости от возраста. Европазавры отличались значительно сниженной скоростью роста, набирая размер медленнее, чем более крупные близкородственные таксоны, такие как камаразавр. Данное замедление темпов роста является противоположностью общей тенденции завропод и многих мезозойских теропод, которые часто достигали бо́льших размеров, обладая повышенными темпами роста[1]. Marpmann et al. (2014) предположили, что небольшой размер и сниженная скорость роста европазавров были следствием педоморфоза, то есть эволюционного процесса, в результате которого у взрослых особей закрепляются черты детёнышей[17].
Карликовость

Было высказано предположение о том, что предок европазавра быстро уменьшился в размерах после миграции на остров. В то время самый большой из островов в регионе нынешней Северной Германии был меньше 200 000 км² и, возможно, не был в состоянии поддерживать сообщество крупных завропод. С другой стороны, предки европазавра могли уменьшаться параллельно с площадью своего местообитания[1]. Предыдущие исследования островной карликовости динозавров в основном ограничивались маастрихтским островом Хацег[англ.] на территории современной Румынии, где обитали карликовые титанозавр мадьярозавр и гадрозавроид Telmatosaurus[1][25][26]. Известно, что голотип Telmatosaurus принадлежит взрослой особи[26], и хотя мадьярозавр был очень маленьким для титатанозавра, подтверждено, что его образцы принадлежат взрослым и старым особям[6]. Взрослые особи Magyarosaurus dacus имели примерно такой же размер тела, что и европазавр, хотя наибольшая известная особь последнего обладала более длинной бедренной костью. «Magyarosaurus» hungaricus был значительно больше, чем европазавр и M. dacus[6]. Карликовость европазавра — единственное задокументированное существенное быстрое изменение массы тела у продвинутого завроподоморфа. Европазавр представляет собой исключение из закона Копа — правила, постулирующего, что в ходе эволюционного развития видов размеры особей имеют тенденцию к увеличению[27].
Палеоэкология

Окаменелости европазавра описаны из отложений местонахождения Лангенберг в Германии, относимых к нижнему оксфорду—верхнему киммериджу. Здесь были обнаружены ископаемые остатки флоры и фауны островной экосистемы времён поздней юры[28]. Из местонахождения Лангенберг происходят остатки как морских, так и наземных животных и растений. Слои с остатками островной биоты могли сформироваться аллохтонно[29]. В известняковых отложениях карьера найдено множество окаменелостей двустворчатых и иглокожих, различные микрофоссилии[30].
Шишки и ветки хвойных идентифицированы как принадлежащие араукариевому Brachyphyllum[англ.]. Из породы было извлечено не менее 450 костей европазавра, причём около 1/3 из них имеют следы зубов[28], по размеру и форме совпадающие с зубами рыб, крокодиломорф и других падальщиков (при этом наличие следов зубов теропод не подтверждено)[28][31]. Большое количество обнаруженных в одном месте особей говорит о том, что, вероятно, стадо европазавров утонуло во время пересечения приливно-отливной зоны. Окаменелости европазавра доминируют среди остатков крупных позвоночных местонахождения. Кроме того, динозавры Лангенберга представлены неопределённым диплодоцидным завроподом, представителем клады стегозавров и тероподами. Диплодоцид известен по трём шейным позвонкам, исходя из размеров которых был сделан вывод, что, вероятно, как и европазавр, этот таксон был островным карликом. Стегозавры и тероподы представлены почти исключительно изолированными зубами. Также обнаружено нескольких костей, возможно, принадлежащих цератозавриду[28]. Изолированные зубы свидетельствуют о том, что в экосистеме присутствовало по меньшей мере четыре таксона теропод: мегалозаврид[англ.] Torvosaurus sp., неопределённые мегалозавриды, аллозавриды и цератозавры, а также, по всей видимости, велоцирапторины (зубы последних, в случае верной идентификации, являются древнейшим известным материалом велоцирапторин)[32][33].
Вымирание
Следы крупных динозавров, сохранившиеся в карьере Лангенберг, указывают на потенциальную причину вымирания европазавра и, возможно, других местных островных карликов. Они расположены на высоте 5 м над пластом с остатками европазавров, что свидетельствует о том, что по крайней мере через 35 000 лет после образования этого отложения произошло падение уровня моря, которое привело к фаунистическому перевороту. Обитавшие на острове тероподы, которые сосуществовали с европазавром, предположительно достигали примерно 4 м в длину, тогда как тероподы, которые прибыли по сухопутному мосту, описаны по следам длиной до 54 см, что указывает на размер тела между 7 и 8 м, если реконструировать их как аллозавроид[англ.]. Авторы описания следов (Lallensack et al., 2015) предположили, что эти инвазионные тероподы стали причиной вымирания эндемичной карликовой фауны, а пласт, из которого происходят следы (пласт Лангенберга №92), вероятно, является самым молодым из тех, в которых присутствуют остатки европазавра[34].
Примечания
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 Sander P. M., Mateus O. V., Laven T., Knotschke N. Bone histology indicates insular dwarfism in a new Late Jurassic sauropod dinosaur (англ.) // Nature : journal. — 2006. — Vol. 441, iss. 7094. — P. 739—741. — ISSN 1476-4687. — doi:10.1038/nature04633. — . — PMID 16760975. Архивировано 18 мая 2021 года.
- ↑ Архангельский М. С,, Иванов А. В. Картины прошлого Земли. Палеоэкологические этюды. — М.: Изд-во «Университетская книга», 2015. — 188 с. — ISBN 978-5-91304-370-2.
- ↑ Europasaurus (англ.) информация на сайте Paleobiology Database. (Дата обращения: 12 апреля 2021).
- ↑ 1 2 3 4 5 6 Sander P. M., Mateus O. V., Laven T., Knotschke N. Supplementary Information for: Bone histology indicates insular dwarfism in a new Late Jurassic sauropod dinosaur (англ.) // Nature : journal. — 2006. — Vol. 441, iss. 7094. — P. 739—741. — ISSN 1476-4687. — doi:10.1038/nature04633. — . — PMID 16760975.
- ↑ Molina-Pérez & Larramendi, 2020, p. 41.
- ↑ 1 2 3 4 Stein K., Csiki Z., Rogers K. C.[англ.], Weishampel D. B., Redelstorff R. Small body size and extreme cortical bone remodeling indicate phyletic dwarfism in Magyarosaurus dacus (Sauropoda: Titanosauria) (англ.) // Proceedings of the National Academy of Sciences : journal. — 2010. — Vol. 107, iss. 20. — P. 9258—9263. — ISSN 0027-8424. — doi:10.1073/pnas.1000781107. — PMID 20435913. Архивировано 27 мая 2020 года.
- ↑ 1 2 3 4 Carballido J. L., Sander P. M. Postcranial axial skeleton of Europasaurus holgeri (Dinosauria, Sauropoda) from the Upper Jurassic of Germany: implications for sauropod ontogeny and phylogenetic relationships of basal Macronaria (англ.) // Journal of Systematic Palaeontology[англ.] : journal. — 2014. — Vol. 12, iss. 3. — P. 335—387. — ISSN 1477-2019. — doi:10.1080/14772019.2013.764935. Архивировано 19 октября 2017 года.
- ↑ 1 2 Carballido J. L., Scheil M., Knotschke N., Sander P. M. The appendicular skeleton of the dwarf macronarian sauropod Europasaurus holgeri from the Late Jurassic of Germany and a re-evaluation of its systematic affinities (англ.) // Journal of Systematic Palaeontology[англ.] : journal. — 2020. — Vol. 18, iss. 9. — P. 739—781. — ISSN 1477-2019. — doi:10.1080/14772019.2019.1683770.
- ↑ 1 2 3 D'Emic M. The early evolution of titanosauriform sauropod dinosaurs (англ.) // Zoological Journal of the Linnean Society[англ.] : journal. — 2012. — Vol. 166, iss. 3. — P. 624—671. — ISSN 0024-4082. — doi:10.1111/j.1096-3642.2012.00853.x. Архивировано 25 июля 2020 года.
- ↑ 1 2 Mannion P. D., Upchurch P., Barnes R. N., Mateus O. Osteology of the Late Jurassic Portuguese sauropod dinosaur Lusotitan atalaiensis (Macronaria) and the evolutionary history of basal titanosauriforms (англ.) // Zoological Journal of the Linnean Society[англ.] : journal. — 2013. — Vol. 168, iss. 1. — P. 98—206. — ISSN 0024-4082. — doi:10.1111/zoj.12029.
- ↑ 1 2 Mannion P. D., Allain R., Moine O. The earliest known titanosauriform sauropod dinosaur and the evolution of Brachiosauridae (англ.) // PeerJ[англ.] : journal. — 2017. — Vol. 5. — ISSN 2167-8359. — doi:10.7717/peerj.3217. — PMID 28480136.
- ↑ 1 2 3 Royo-Torres R., Fuentes C., Meijide M., Meijide-Fuentes F., Meijide-Fuentes M. A new Brachiosauridae Sauropod dinosaur from the lower Cretaceous of Europe (Soria Province, Spain) (англ.) // Cretaceous Research : journal. — 2017. — Vol. 80. — P. 38—55. — ISSN 0195-6671. — doi:10.1016/j.cretres.2017.08.012.
- ↑ 1 2 3 4 5 Wings O. Auf Zwergdinojagd im Langenberg-Steinbruch bei Goslar (нем.) // Fossilien. — 2014. — Bd. 2. — S. 44—50.
- ↑ 1 2 Knotschke N. Erste Ergebnisse und Erfahrungen bei Bergung und Praparation einer Fauna aus dem oberen Jura (Kimmeridgegebirge) (нем.) // Der Praparator. — Hannover, 2001. — Bd. 47, Nr. 1. — S. 7–13. — ISSN 0032-6542.
- ↑ Knotschke N., Rossler D. Neue Ergebnisse des "Vereins zur Forderung der niedersachsischen Palaontologie e.V. (нем.) // Der Praparator. — Hannover, 2002. — Bd. 48, Nr. 1. — S. 21–30. — ISSN 0032-6542.
- ↑ 1 2 Knotschke N., Mastroianni M., Wings O. A song of blasting and fire: Europasaurus holgeri (англ.) // 74th Annual Meeting of the Society of Vertebrate Paleontology: Program and Abstracts. — Berlin, 2014.
- ↑ 1 2 Marpmann J. S., Carballido J. L., Sander P. M., Knotschke N. Cranial anatomy of the Late Jurassic dwarf sauropod Europasaurus holgeri (Dinosauria, Camarasauromorpha): ontogenetic changes and size dimorphism (англ.) // Journal of Systematic Palaeontology[англ.] : journal. — 2014. — Vol. 13, iss. 3. — P. 221—263. — ISSN 1477-2019. — doi:10.1080/14772019.2013.875074.
- ↑ 1 2 Taylor M. P.[англ.]. A re-evaluation of Brachiosaurus altithorax Riggs 1903 (Dinosauria, Sauropoda) and its generic separation from Giraffatitan brancai (Janensch 1914) (англ.) // Journal of Vertebrate Paleontology : journal. — 2009. — Vol. 29, iss. 3. — P. 787—806. — ISSN 0272-4634. — doi:10.1671/039.029.0309.
- ↑ Barrett P. M., Benson R. B. J., Upchurch P. Dinosaurs of Dorset: Part II, the sauropod dinosaurs (Saurischia, Sauropoda) with additional comments on the theropods (англ.) // Proceedings of the Dorset Natural History and Archaeological Society : journal. — 2010. — Vol. 131. — P. 113—126. — ISSN 0070-7112.
- ↑ Sereno P. C. Somphospondyli (англ.). TaxonSearch. The University of Chicago. Дата обращения: 10 апреля 2021. Архивировано 14 апреля 2021 года.
- ↑ Wilson J. A.[англ.]. Sauropod dinosaur phylogeny: critique and cladistic analysis (англ.) // Zoological Journal of the Linnean Society[англ.] : journal. — 2002. — Vol. 136, iss. 2. — P. 215—275. — ISSN 0024-4082. — doi:10.1046/j.1096-3642.2002.00029.x.
- ↑ Upchurch P. The phylogenetic relationships of sauropod dinosaurs (англ.) // Zoological Journal of the Linnean Society[англ.] : journal. — 1998. — Vol. 124, iss. 1. — P. 43—103. — ISSN 0024-4082. — doi:10.1111/j.1096-3642.1998.tb00569.x.
- ↑ Weishampel, Dodson & Osmolska, 2004, chpt. 13: "Sauropoda" by P. Upchurch, P. M. Barrett, P. Dodson, pp. 259—232.
- ↑ D’Emic M. D., Foreman B. Z., Jud N. A. Anatomy, systematics, paleoenvironment, growth, and age of the sauropod dinosaur Sonorasaurus thompsoni from the Cretaceous of Arizona, USA (англ.) // Journal of Paleontology[англ.] : journal. — 2016. — Vol. 90, iss. 1. — P. 102—132. — ISSN 1937-2337 0022-3360, 1937-2337. — doi:10.1017/jpa.2015.67. Архивировано 14 апреля 2021 года.
- ↑ Nopcsa F. Über das Vorkommen der Dinosaurier in Siebenbürgen (нем.) // Ver. Zool. Bot. Ges. Wien. — 1914. — Bd. 54. — S. 12—14.
- ↑ 1 2 Weishampel D. B., Norman D.B., Grigorescu D. Telmatosaurus transsylvanicus from the Late Cretaceous of Romania: the most basal hadrosaurid dinosaur (англ.) // Palaeontology[англ.] : journal. — 1933. — P. 361—385.
- ↑ Benson R. B. J., Campione N. E., Carrano M.T., Mannion P. D., Sullivan C., Upchurch P., Evans D.C. Rates of Dinosaur Body Mass Evolution Indicate 170 Million Years of Sustained Ecological Innovation on the Avian Stem Lineage (англ.) // PLOS Biology : journal. — 2014. — Vol. 12, iss. 5. — P. e1001853. — ISSN 1544-9173. — doi:10.1371/journal.pbio.1001853. Архивировано 29 марта 2020 года.
- ↑ 1 2 3 4 Wings O. The Langenberg Quarry near Goslar: Unique window into a terrestrial Late Jurassic ecosystem in Northern Germany (англ.) // Abstracts of the 12th Symposium on Mesozoic Terrestrial Ecosystems. — 2015. — P. 99—100. Архивировано 10 апреля 2019 года.
- ↑ Schwarz D., Raddatz M., Wings O. Knoetschkesuchus langenbergensis gen. nov. sp. nov., a new atoposaurid crocodyliform from the Upper Jurassic Langenberg Quarry (Lower Saxony, northwestern Germany), and its relationships to Theriosuchus (англ.) // PLOS One : journal. — 2017. — Vol. 12, iss. 2. — P. e0160617. — ISSN 1932-6203. — doi:10.1371/journal.pone.0160617. Архивировано 27 апреля 2022 года.
- ↑ Wings O. Unique taphonomy of the dwarfed sauropod Europasaurus from Late Jurassic marine strata of the Langenberg Quarry (Lower Saxony, Northern Germany) (англ.) // Journal of Vertebrate Paleontology, Program and Abstracts. 73rd Annual Meeting of the Society of Vertebrate Paleontology. — 2013. — P. 239.
- ↑ Slodownik M., Wings O. Bite marks on Europasaurus bones from the Langenberg Quarry near Goslar (Lower Saxony, Germany) (англ.) // 13th Annual Meeting of the European Association of Vertebrate Palaeontologists Opole, Poland, Abstracts. — 2015. — P. 141.
- ↑ Gerke O., Wings O. Multivariate and Cladistic Analyses of Isolated Teeth Reveal Sympatry of Theropod Dinosaurs in the Late Jurassic of Northern Germany (англ.) // PLOS One : journal. — 2016. — Vol. 11, iss. 7. — P. e0158334. — ISSN 1932-6203. — doi:10.1371/journal.pone.0158334. Архивировано 26 января 2022 года.
- ↑ Lubbe T. van Der, Richter U., Knotschke N. Velociraptorine Dromaeosaurid Teeth from the Kimmeridgian (Late Jurassic) of Germany (англ.) // Acta Palaeontologica Polonica : journal. — 2009. — Vol. 54, iss. 3. — P. 401—408. — ISSN 0567-7920. — doi:10.4202/app.2008.0007. Архивировано 14 апреля 2021 года.
- ↑ Lallensack J. N., Sander M. P., Knotschke N., Wings O. Dinosaur tracks from the Langenberg Quarry (Late Jurassic, Germany) reconstructed with historical photogrammetry: Evidence for large theropods soon after insular dwarfism // Palaeontologia Electronica[англ.] : journal. — 2015. — ISSN 1094-8074. — doi:10.26879/529.
Литература
- The Dinosauria[англ.] (англ.) / D. B. Weishampel, P. Dodson, H. Osmolska, eds. — 2nd ed. — Berkeley: University of California Press, 2004. — 880 p. — ISBN 978-0-520-25408-4.
- Molina-Pérez R., Larramendi A. Dinosaur Facts and Figures: The Sauropods and Other Sauropodomorphs (англ.) / Illustrations by A. Atuchin and S. Mazzei. — Princeton, New Jersey: Princeton University Press, 2020. — 272 p. — ISBN 978-0-691-20297-6.
















