Желе́зо — химический элемент 8-й группы четвёртого периода периодической системы химических элементов Д. И. Менделеева с атомным номером 26.

Цитохром — небольшой гем-содержащий белок, относится к классу цитохромов, содержит в структуре гем типа c. Выполняет в клетке две функции. С одной стороны, он является одноэлектронным переносчиком, свободно связанным с внутренней мембраной митохондрий, и необходимым компонентом дыхательной цепи. Он способен окисляться и восстанавливаться, но не связывает при этом кислород. С другой стороны, при определённых условиях он может отсоединяться от мембраны, переходить в раствор в межмембранном пространстве и активировать апоптоз. Такая двойственность связана со специфичными свойствами молекулы цитохрома c.

Циани́ды — соли цианистоводородной (синильной) кислоты. В номенклатуре IUPAC к цианидам относят также C-производные синильной кислоты — нитрилы.

Карбони́лы мета́ллов, карбони́льные_ко́мплексы — координационные комплексы переходных металлов с монооксидом углерода, являющегося лигандом. Многие карбонилы металлов очень летучи.

Цистеи́н — алифатическая серосодержащая аминокислота. Оптически активна, существует в виде L- и D- изомеров. L-Цистеин входит в состав белков и пептидов, играет важную роль в процессах формирования тканей кожи. Имеет значение для дезинтоксикационных процессов.
Дыхательная цепь переноса электронов, также электрон-транспортная цепь (сокр. ЭТЦ, англ. ETC, Electron transport chain) — система трансмембранных белков и переносчиков электронов, необходимых для поддержания энергетического баланса. ЭТЦ поддерживает баланс за счёт переноса электронов и протонов из НАД∙Н и ФАДН2 в акцептор электронов. В случае аэробного дыхания акцептором может быть молекулярный кислород (О2). В случае анаэробного дыхания акцептором могут быть NO3−, NO2−, Fe3+, фумарат, диметилсульфоксид, сера, SO42−, CO2 и т. д. ЭТЦ у прокариот локализована в ЦПМ, у эукариот — на внутренней мембране митохондрий. Переносчики электронов расположены в порядке уменьшения сродства к электрону, то есть по своему окислительно-восстановительному потенциалу, где у акцептора самое сильное сродство к электрону. Поэтому транспорт электрона на всём протяжении цепи протекает самопроизвольно с выделением энергии. Выделение энергии в межмембранное пространство при переносе электронов происходит ступенчато, в виде протона (H+). Протоны из межмембранного пространства попадают в протонную помпу, где наводят протонный потенциал. Протонный потенциал преобразуется АТФ-синтазой в энергию химических связей АТФ. Сопряжённая работа ЭТЦ и АТФ-синтазы носит название окислительного фосфорилирования.

Окисли́тельное фосфорили́рование — метаболический путь, при котором энергия, образовавшаяся при окислении питательных веществ, запасается в митохондриях клеток в виде АТФ. Хотя различные формы жизни на Земле используют разные питательные вещества, АТФ является универсальным соединением, в котором запасается энергия, необходимая для других метаболических процессов. Почти все аэробные организмы осуществляют окислительное фосфорилирование. Вероятно, широкому распространению этого метаболического пути способствовала его высокая энергетическая эффективность по сравнению с анаэробным брожением.

Кластер дифференцировки — номенклатура дифференцировочных антигенов лейкоцитов человека. Данная классификация была предложена в 1982 году для идентификации и исследования поверхностных мембранных белков лейкоцитов. CD-антигенами могут быть белки, которые служат рецепторами или лигандами, участвующими во взаимодействии клеток между собой и являющимися компонентами каскада определённых сигнальных путей. Однако, они могут быть и белками, выполняющими другие функции. Список CD-антигенов, внесённых в номенклатуру, постоянно пополняется и в настоящее время содержит 350 CD-антигенов и их подтипов.
Кофактор — небольшое небелковое соединение, которое присоединяется к функциональному участку белка и участвует в его биологической деятельности. Такие белки обычно являются ферментами, поэтому кофакторы называют «молекулами-помощниками», которые участвуют в биохимических превращениях.

Пируватдегидрогена́зный ко́мплекс, ПДК (англ. Pyruvate dehydrogenase complex, PDH, PDC) — белковый комплекс, осуществляющий окислительное декарбоксилирование пирувата. Он включает в себя три фермента и два вспомогательных белка, а для его функционирования необходимы пять кофакторов (СоА, NAD+, тиаминпирофосфат (ТРР), FAD и липоевая кислота (липоат)). PDH локализован у бактерий в цитозоле, а у эукариот — в митохондриальном матриксе. Суммарное уравнение катализируемой реакции таково:

Гем b — наиболее часто встречающийся гем, как в гемоглобине, так и в миоглобине. Такое семейство ферментов как пероксидазы тоже содержат в себе гем Б. Циклооксигеназа-1 и циклооксигеназа-2 из малоизвестного семейства семейства циклооксигеназ содержат в себе гем Б как один из своих активных сайтов.
Гипотеза мира сульфидов железа — гипотетический этап возникновения жизни на Земле и ранней эволюции, предложенный Гюнтером Вэхтерсхойзером, юристом из Мюнхена, имеющим также научную степень по химии. Опубликовал свои идеи при поддержке философа Карла Р. Поппера. Как следует из названия теории, она предполагает, что жизнь могла зародиться на поверхности кристаллов сульфидов железа.

Гем С — вид гема, отличается от гема B наличием тиольных групп.
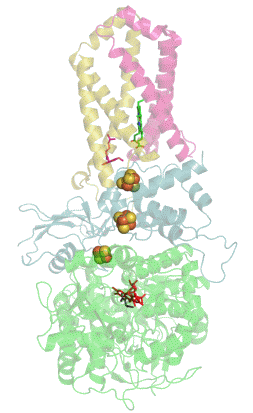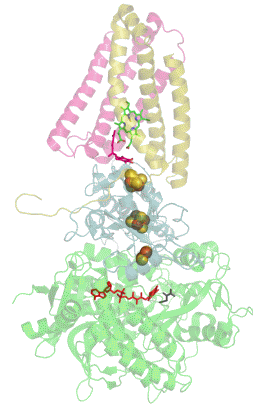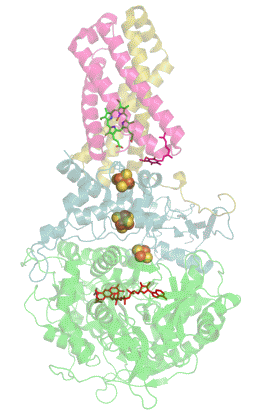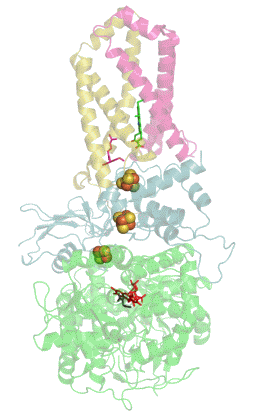
Сукцинатдегидрогеназа или сукцинат-убихинон-оксидорекдуктаза, также известная как комплекс II — белковый комплекс, расположенный во внутренней мембране митохондрий и мембранах многих прокариотических организмов. Одновременно участвует в цикле трикарбоновых кислот и дыхательной цепи переноса электронов.

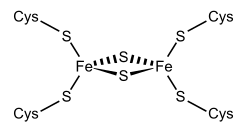
Железосерные кластеры — элементоорганические соединения, группа белковых кофакторов, обладающих окислительно-восстановительным (Red/Ox) потенциалом в районе от −500 мВ до +300 мВ. Red/Ox-потенциал зависит от структуры и конформации белка, что делает эти кофакторы важнейшими участниками окислительно-восстановительных реакций в клетке. Железосерные кластеры способны принимать или отдавать электроны. Белки, содержащие железосерные кластеры, являются эволюционно древними и распространены во всех царствах, включая животных, растения, грибы, бактерии и археи. Мутации по генам метаболизма Fe—S-кластеров являются причиной многих тяжёлых заболеваний или летальны.
Ферредокси́ны — группа небольших растворимых белков, содержащих железосерные кластеры и являющихся подвижными переносчиками электронов в ряде метаболических процессов. Обычно они переносят один или два электрона за счёт изменения окисленности атомов железа.

Фотосисте́ма I, или пластоциани́н-ферредокси́н-оксидоредукта́за — второй функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он принимает электрон от пластоцианина и, поглощая световую энергию, формирует сильный восстановитель П700, способный через цепь переносчиков электронов осуществить восстановление НАДФ+. Таким образом, при участии ФСI синтезируется источник электронов (НАДФН) для последующих реакций восстановления углерода в хлоропластах в цикле Кальвина. Кроме того, ФСI может осуществлять циклический транспорт электронов, сопряжённый с синтезом АТФ, обеспечивая дополнительный синтез АТФ в хлоропластах.

Фотосисте́ма II (втора́я фотосисте́ма, фотосисте́ма два, ФСII), или H2O-пластохиноноксидоредуктаза — первый функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он расположен в мембранах тилакоидов всех растений, водорослей и цианобактерий. Поглощая энергию света в ходе первичных фотохимических реакций, он формирует сильный окислитель — димер хлорофилла a (П680+), который через цепь окислительно-восстановительных реакций способен вызвать окисление воды.

НАДН-дегидрогена́зный ко́мплекс, также называемый ко́мплекс I или НАДН-убихино́н-оксидоредукта́за — первый мультибелковый комплекс дыхательной цепи переноса электронов. Множество копий комплекса расположены в мембранах прокариотических организмов, способных к кислородному дыханию и внутренних мембранах митохондрий эукариотических клеток. По отношению к белкам человека комплекс I часто называют НАДН-дегидрогеназой.

Белки́, зая́коренные липи́дами, или липидосвя́занные белки́ — белки клеточной мембраны, ковалентно связанные с липидами клеточной мембраны. Эти липиды вставлены в мембрану бок о бок с хвостами жирных кислот. Белки, заякоренные липидами, могут находиться с любой стороны клеточной мембраны. Таким образом, липид служит своего рода якорем, закрепляющим белок вблизи клеточной мембраны.