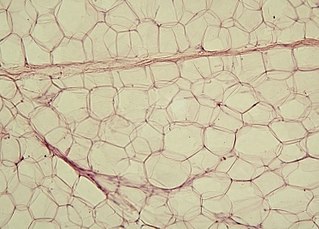
Жировая ткань

Жирова́я ткань — разновидность соединительной ткани животных, образующаяся из мезенхимы и состоящая из специализированных клеток, накапливающих жиры — адипоцитов. Почти весь адипоцит заполняет жировая капля, окружённая ободком цитоплазмы с оттеснёнными на периферию клеточным ядром и относительно небольшим количеством митохондрий. Помимо адипоцитов, в состав жировой ткани входят клетки так называемой стромальной васкулярной фракции: преадипоциты, фибробласты, клетки эндотелия сосудов и ряд иммунных клеток, такие как макрофаги жировой ткани[англ.][1].
Главная функция жировой ткани — запасание липидов, кроме того, она обеспечивает теплоизоляцию тела. Жировая ткань продуцирует ряд гормонов (эстроген, лептин, резистин[англ.], а также цитокины, такие как фактор некроза опухоли α), и в последние годы была признана важной частью эндокринной системы. У взрослого человека жировая ткань располагается под кожей, вокруг внутренних органов (висцеральная жировая ткань), внутри костей (жёлтый костный мозг), между мышечными волокнами и в молочных железах[1].
Жировую ткань подразделяют на белую и бурую. Белая жировая ткань запасает липиды, а главной функцией бурой жировой ткани является термогенез. Бурая жировая ткань наиболее развита у новорождённых, а также животных, впадающих в спячку[2]. У взрослого человека бурая ткань присутствует и метаболически активна, однако она регрессирует с возрастом.
Строение

Клетки жировой ткани, накапливающие жир, называют адипоцитами. Одиночные адипоциты имеют шарообразную форму. Жировую ткань принято подразделять на белую и бурую согласно её цвету. Адипоцит белой жировой ткани содержит одну большую каплю нейтрального жира (такие адипоциты также называют унилокулярными), которая занимает центральную часть клетки и окружена тонким слоем цитоплазмы, в утолщённой части которого залегает уплощённое ядро. В цитоплазме адипоцитов содержатся в небольших количествах и другие липиды: холестерин, фосфолипиды, а также свободные жирные кислоты. Эти мелкие жировые включения особенно выражены у незрелых адипоцитов. Зрелый адипоцит имеет крупные размеры, от 50 до 150 мкм. Так как липиды вымываются ксилолом и другими растворителями, использующимися при приготовлении гистологических препаратов, унилокулярные адипоциты при рассмотрении с помощью светового микроскопа выглядят пустыми[3].
Рыхлая[англ.] волокнистая соединительная ткань образует прослойки, которые делят жировую ткань на дольки разного размера и формы. В дольках адипоциты близко прилегают друг к другу, однако в жировой ткани также присутствуют клетки так называемой стромальной васкулярной фракции: преадипоциты, фибробласты, клетки эндотелия сосудов и ряд иммунных клеток, такие как макрофаги жировой ткани[1]. На клетки стромальной васкулярной фракции приходится около половины всех клеток жировой ткани[4]. Жировые клетки разделяются тонкими коллагеновыми волокнами, ориентированными во всех направлениях, а также оплетены ретикулярными волокнами[4]. Группы адипоцитов или отдельные дольки тесно охватываются кровеносными и лимфатическими капиллярами[5].

У новорождённых детей и некоторых животных (грызунов и животных, впадающих в спячку) выражена бурая жировая ткань. Адипоциты бурой жировой ткани, по сравнению с клетками белой жировой ткани, имеют больше митохондрий и вместо одной крупной жировой капли содержат множество мелких жировых включений в цитоплазме (такие адипоциты называют мультилокулярными[6]). Бурый цвет обеспечивается железосодержащими пигментами цитохромами, расположенными в митохондриях. Изменения бурой жировой ткани при голодании выражены меньше, чем белой[7].
Термином «бежевый жир» называют белую жировую ткань, которая приобретает некоторые черты бурой жировой ткани, например, в её адипоцитах вместо одной крупной жировой ткани имеется несколько включений меньшего размера, увеличивается количество митохондрий и повышается уровень экспрессии гена UCP1, кодирующего белок термогенин[8].
Четвёртый тип адипоцитов был недавно описан в составе подкожной жировой ткани мышей во время беременности и лактации, когда жировая ткань в молочных железах существенно сокращается, а железистая часть, наоборот, разрастается. Новосформированные эпителиальные клетки, входящие в состав железистой части, называют розовыми адипоцитами. Они появляются в результате прямой трансдифференцировки[англ.] белых адипоцитов в эпителиальные клетки, продуцирующие молоко. Образование розовых адипоцитов обратимо, и по завершении лактации они превращаются обратно в белые адипоциты, восстанавливая жировую часть молочной железы[9].
Плотность жировой ткани составляет около 0,9 г/мл против 1,06 г/мл для мышечной ткани, поэтому человек, имеющий больше жира, будет держаться на воде легче, чем человек той же массы, но с большей долей мышечной массы[10][11].
Анатомия

У взрослого человека белая жировая ткань располагается под кожей, особенно в нижней части брюшной стенки, на ягодицах и бёдрах (в составе подкожной ткани), вокруг внутренних органов (висцеральная жировая ткань), внутри костей (жёлтый костный мозг[англ.]), между мышечными волокнами и в молочных железах. Бурая жировая ткань, выраженная у новорождённых детей и некоторых животных (грызунов и млекопитающих, впадающих в спячку), располагается на шее, около лопаток, за грудиной, вдоль позвоночника, под кожей и между мышцами[7]. У взрослого человека бурая ткань присутствует и метаболически активна[12][13], однако она регрессирует с возрастом[14]. У человека типичная бурая жировая ткань находится между лопаток, вокруг почек, в шее, надключичной области и вдоль позвоночника. Кроме того, по всей белой жировой ткани встречаются так называемые бежевые адипоциты — белые адипоциты, приобретшие некоторые черты бурых адипоцитов[15].
Преимущественные места развития жировой ткани и накопления в них излишнего жира имеют половые отличия, отсюда разделяется ожирение по женскому (гиноидное) или мужскому (абдоминальное) типу. При некоторых эндокринных нарушениях у мужчин может наблюдаться ожирение по женскому типу[].
Физиология
Метаболизм жиров
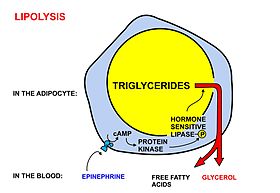
Жировая ткань играет важную роль в поддержании уровня свободных жирных кислот и триглицеридов в крови, а также вносит вклад в развитие инсулинорезистентности (особенно абдоминальный жир). Адипоциты также могут запасать триглицериды, поступающие с пищей и циркулирующие в крови в составе хиломикронов, липиды, синтезируемые печенью и циркулирующие в кровотоке в виде липопротеинов очень низкой плотности, кроме того, свободные жирные кислоты и глицерин могут синтезироваться в самих адипоцитах. Хиломикроны и липопротеины очень низкой плотности при поступлении в жировую ткань гидролизуются липопротеинлипазой на люминальной поверхности кровеносных капилляров. Свободные жирные кислоты поступают в адипоциты по механизму активного транспорта и диффузии. В адипоцитах жирные кислоты в ходе реакции этерификации присоединяются к глицеринфосфату с образованием триглицеридов, которые поступают в жировую каплю[16].
В жировой ткани идёт постоянное поступление и выход свободных жирных кислот. Результирующее направление движения свободных жирных кислот контролируются гормонами инсулином и лептином. Если инсулин повышен, то вход свободных жирных кислот в жировую ткань превышает её выход, и выход жирных кислот из жировой ткани возможен только при низком уровне инсулина в крови. Уровень инсулина повышается при поступлении в организм углеводной пищи, которое приводит к росту концентрации сахара в крови[17]. Инсулин также стимулирует поглощение глюкозы адипоцитами и способствует её преобразованию в жир[18].
При нервной или гуморальной стимуляции адипоцитов жировые запасы мобилизуются и клетки высвобождают жирные кислоты и глицерин. Норадреналин, выделяемый надпочечниками и постганглионарными[англ.] симпатическими окончаниями, активирует гормончувствительную липазу, которая расщепляет триглицериды на поверхности липидных капель. Эта липаза также активируется гипофизарным гормоном роста. Свободные жирные кислоты диффундируют через мембраны адипоцитов и эндотелиальных клеток, выходят в кровоток и связываются с белком альбумином. Более гидрофильный глицерин свободно плавает в крови и поглощается печенью. Инсулин ингибирует гормончувствительную липазу[19]. Мобилизацию адипоцитов также запускают адреналин[20] и адренокортикотропный гормон[21][22].
Продукция гормонов

Молекулы, продуцируемые жировой тканью, играют важнейшую роль в поддержании метаболического гомеостаза, и нарушения в их образовании могут приводить к развитию ожирения и ряда патологических состояний, связанных с ожирением, поэтому жировую ткань рассматривают как эндокринный орган. Гормоны жировой ткани в совокупности называют адипокинами. Адипокины представляют собой разновидность цитокинов (сигнальных белков). Первым открытым адипокином стал гормон лептин, описанный в 1994 году. Лептин играет роль в поддержании нормальной массы тела и передаёт сигнал, свидетельствующий о насыщении, в гипоталамус. Лептин также контролирует липогенез в гепатоцитах, подавляя путь биосинтеза жирных кислот, и способствует окислению жирных кислот в мышцах. Наиболее обильно продуцируется адипокин, известный как адипонектин. Он повышает чувствительность к инсулину, и его введение мышам, страдающим ожирением, позволило частично преодолеть инсулинорезистентность. К числу адипокинов также относится фактор некроза опухоли α (TNFα), который вовлечён в формирование инсулинорезистентности за счёт подавления сигнального пути инсулина[англ.]. В жировой ткани TNFα продуцируют макрофаги и другие иммунные клетки. У людей и мышей, страдающих ожирением, в жировой ткани возрастает экспрессия провоспалительного цитокина[англ.] интерлейкина 6 (IL-6), однако его роль в метаболизме глюкозы неясна[9]. Также к числу адипокинов относят аспросин[23], резистин[англ.][24], апелин[25], хемерин[англ.][26], CCL2[27] и некоторые другие цитокины. Лептин и резистин продуцируются преимущественно подкожной жировой тканью[28]. Кроме того, и у женщин, и у мужчин жировая ткань является главным периферическим источником ароматазы, которая участвует в синтезе эстрогенов[29].
Термогенез
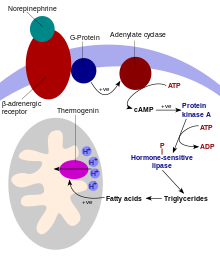
Основная функция бурой жировой ткани — термогенез. У животных в конце спячки и новорождённых детей в бурую жировую ткань поступает норадреналин, который, как и в белой жировой ткани, стимулирует гормончувствительную липазу и запускает гидролиз триглицеридов. Однако, в отличие от белых адипоцитов, в бурых адипоцитах свободные жирные кислоты не высвобождаются в кровь, а быстро метаболизируются, что сопровождается повышением потребления кислорода и продукцией тепла. Локальное повышение температуры в бурой жировой ткани приводит к нагреванию омывающей её крови, которая передаёт тепло на весь организм. Усиленная продукция тепла в бурых адипоцитах возможна благодаря тому, что в их внутренних митохондриальных мембранах в большом количестве содержится трансмембранный разобщающий белок термогенин, или UCP1. В присутствии свободных жирных кислот термогенин позволяет протонам поступать из межмембранного пространства непосредственно в матрикс митохондрии без прохождения протонов через АТФ-синтазу. Вместо образования АТФ энергия протонов идёт на выделение тепла[30]. Считается, что термогенин является симпортером протонов и свободных жирных кислот, но конкретный механизм его действия неясен[31]. Известно, что термогенин ингибируют АТФ, АДФ и ГТФ[32]. Термогенез в бурых адипоцитах также может активироваться при переедании[33].
Развитие
Как и другие клетки соединительной ткани, адипоциты происходят от мезенхимальных стволовых клеток[англ.]. Мезенхимальные стволовые клетки дают начало преадипоцитам, которые похожи на крупные фибробласты с цитоплазматическими липидными включениями. Первоначально липидные капли молодого белого адипоцита изолированы друг от друга, но вскоре они сливаются с образованием единой большой жировой капли. Белые адипоциты развиваются вместе с меньшей популяцией бежевых адипоцитов, которые присутствуют в зрелой белой жировой ткани. При адаптации к низким температурам белые адипоциты частично обратимо превращаются в бурые, приобретают большое количество мелких липидных капель вместо одной крупной, их профиль экспрессии генов становится близок к таковому у бурых адипоцитов (в частности, возрастает экспрессия гена UCP1, кодирующего термогенин), и так называемые бежевые адипоциты приступают к термогенезу[34]. При возвращении к нормальным условиям часть бежевых адипоцитов вновь становятся белыми. У мышей «побурение» белой жировой ткани полностью нивелируется за 21 день после окончания пребывания на холоде, а снижение экспрессии UCP1, кодирующего термогенин, наступает уже через 24 часа[35]. При повторном попадании на холод в бежевые адипоциты превращаются каждый раз одни и те же белые адипоциты[36]. Превращение белого адипоцита в бежевый контролируется несколькими транскрипционными факторами[37]: PPARγ[англ.], PRDM16[англ.][38], PGC-1α[англ.] и EBF2[39][40][41]. «Побурение» белого жира также стимулируют иризин, секретируемый мышечной тканью в ответ на физическую нагрузку[42], и FGF21[англ.], выделяемый печенью[8]. У мышей «побурение» стимулируют метионин-энкефалиновые пептиды, продуцируемые лимфоидными клетками врождённого иммунитета 2 типа в ответ на действие интерлейкина 33 (IL-33)[43].
Бурые адипоциты также развиваются от мезенхимальных стволовых клеток, но в других локациях тела эмбриона, отличных от тех, где происходит дифференцировка белых адипоцитов. Бурые адипоциты в ходе эмбрионального развития возникают раньше белых. У человека объём бурой жировой ткани относительно массы тела максимален при рождении, когда наиболее высока потребность в термогенезе, и в детстве почти полностью исчезает через инволюцию и апоптоз адипоцитов. У взрослых бурый жир наиболее активен у людей худощавого телосложения. При адаптации к холоду бежевые адипоциты могут превращаться в бурые, кроме того, возможна пролиферация и дифференцировка бурых адипоцитов от мезенхимальных клеток-предшественников. Автономные нервы не только стимулируют термогенную активность бурых адипоцитов, но также способствуют их дифференцировке и предотвращают апоптоз зрелых бурых адипоцитов[44].
Клиническое значение

Белые адипоциты могут давать начало часто встречающимся доброкачественным образованиям — липомам. Злокачественные опухоли, происходящие из жировой ткани — липосаркомы — относительно редки. Доброкачественные опухоли, образованные бурыми адипоцитами, иногда называют гиберномами[англ.][3].

Под ожирением понимают состояние, при котором в организме накапливается избыток жировой ткани[45]. Ожирение повышает риск возникновения многих заболеваний и патологических состояний: сердечно-сосудистых, сахарного диабета 2-го типа, обструктивного апноэ во сне[англ.], некоторых видов рака, а также остеоартрита[46]. Избыточное разрастание висцерального жира, в особенности, вокруг желудка называют центральным, или висцеральным ожирением, а чрезмерно увеличенный, выдающийся живот при этом состоянии известен как «пивной живот». Поскольку жировая ткань продуцирует множество цитокинов, в том числе и провоспалительных, ожирение часто сопровождается умеренным хроническим воспалением. Сахарный диабет и болезни сердца относят к воспалительным заболеваниям, связанным с ожирением[34]. Избыток жировой ткани, особенно висцерального жира, может приводить к появлению инсулинорезистентности[47]. У большинства пациентов, страдающих ожирением, адипоциты производят нормальное или повышенное количество лептина, однако иногда его клетки-мишени имеют недостаточно рецепторов лептина или несут дефектные рецепторы, поэтому эффект насыщения, опосредуемый лептином, не наступает[34]. Однако мутации в гене, кодирующем лептин, могут объяснить лишь небольшую долю случаев ожирения[48]. Весьма частой причиной развития ожирения у взрослых являются возрастные метаболические нарушения, при которых снижается активность гормончувствительной липазы. Повышенное количество адипоцитов, сформированных при детском ожирении, повышает риск ожирения у человека в старшем возрасте[6]. Конвертацию белой жировой ткани в бурую рассматривают как перспективную стратегию терапии ожирения[49].
В настоящее время жировую ткань можно использовать в качестве источника стволовых клеток у взрослых[англ.]. Стволовые клетки жировой ткани можно легко перепрограммировать в индуцированные плюрипотентные стволовые клетки[50]. Получение стволовых клеток из клеточного материала самого организма пациента снижает риск отторжения трансплантата и позволяет избежать многих этических проблем, связанных с использованием эмбриональных стволовых клеток[51]. Имеются сведения, что стволовые клетки из разных локаций жировой ткани (абдоминального жира, эпикардиального жира и других) имеют разные свойства[51][52]: скорость пролиферации, иммунофенотип, потенциал дифференцировки и устойчивость к гипоксии[53].
Возрастные изменения
Во время старения висцеральная жировая ткань и межмышечный жир имеют тенденцию к увеличению, тогда как периферическая подкожная жировая ткань с возрастом значительно снижается[54][55][56][57].
Возможная причина истончения подкожной жировой ткани у пожилых кроется в многократном росте синтеза транскрипционного фактора Pu.1 в некоторых клетках подкожной жировой ткани при старении.[58] Обнаружено также, что задерживать уменьшение толщины подкожной белой жировой ткани и накопление в ней стареющих клеток может делеция гена Nrip1 (Nuclear receptor-interacting protein 1)[59].
История изучения

Жировая ткань (точнее, бурая жировая ткань) впервые была описана в 1551 году швейцарским натуралистом Конрадом Геснером[60]. В 1902 году было отмечено сходство между шейными жировыми отложениями у новорождённых младенцев и млекопитающих, впадающих в спячку. Активное исследование бурой жировой ткани возобновилось в 1960-х годах (в 1964 году Силверман и коллеги доказали, что у человека бурый жир также отвечает за термогенез), и к 1980-м годам установилось мнение, что у взрослых людей бурой жировой ткани нет. Это представление было пересмотрено в конце 2000-х годов[61].
Белые адипоциты, или «жировые везикулы», а также их вклад в рост жировых отложений впервые были описаны в XIX веке. Активное исследование жировой ткани началось лишь в 1940-х годах. В 1940 году было показано, что жировая ткань иннервируется и снабжается кровью. В 1950-х годах была прояснена роль белых адипоцитов в метаболизме липидов, и дальнейшее изучение регуляции работы жировой ткани продолжалось во всей второй половине XX века[62]. Первые данные, свидетельствующие об эндокринной функции белой жировой ткани, появились в 1980-х годах[63].
Примечания
- ↑ 1 2 3 Aarsland A., Chinkes D., Wolfe R. R. Hepatic and whole-body fat synthesis in humans during carbohydrate overfeeding. (англ.) // The American Journal Of Clinical Nutrition. — 1997. — June (vol. 65, no. 6). — P. 1774—1782. — doi:10.1093/ajcn/65.6.1774. — PMID 9174472.
- ↑ Gesta S., Tseng Y. H., Kahn C. R. Developmental origin of fat: tracking obesity to its source. (англ.) // Cell. — 2007. — 19 October (vol. 131, no. 2). — P. 242—256. — doi:10.1016/j.cell.2007.10.004. — PMID 17956727.
- ↑ 1 2 Mescher, 2016, p. 122.
- ↑ 1 2 Mescher, 2016, p. 123.
- ↑ Афанасьев и др., 2004, с. 230—231.
- ↑ 1 2 Mescher, 2016, p. 126.
- ↑ 1 2 Афанасьев и др., 2004, с. 231—232.
- ↑ 1 2 Harms M., Seale P. Brown and beige fat: development, function and therapeutic potential. (англ.) // Nature Medicine. — 2013. — October (vol. 19, no. 10). — P. 1252—1263. — doi:10.1038/nm.3361. — PMID 24100998.
- ↑ 1 2 Colaianni Graziana, Colucci Silvia, Grano Maria. Anatomy and Physiology of Adipose Tissue (англ.) // Multidisciplinary Approach to Obesity. — 2014. — 15 October. — P. 3—12. — ISBN 9783319090443. — doi:10.1007/978-3-319-09045-0_1.
- ↑ Farvid M. S., Ng T. W., Chan D. C., Barrett P. H., Watts G. F. Association of adiponectin and resistin with adipose tissue compartments, insulin resistance and dyslipidaemia. (англ.) // Diabetes, Obesity & Metabolism. — 2005. — July (vol. 7, no. 4). — P. 406—413. — doi:10.1111/j.1463-1326.2004.00410.x. — PMID 15955127.
- ↑ Urbanchek M. G., Picken E. B., Kalliainen L. K., Kuzon Jr. W. M. Specific force deficit in skeletal muscles of old rats is partially explained by the existence of denervated muscle fibers. (англ.) // The Journals Of Gerontology. Series A, Biological Sciences And Medical Sciences. — 2001. — May (vol. 56, no. 5). — P. 191—197. — doi:10.1093/gerona/56.5.b191. — PMID 11320099.
- ↑ Nedergaard J., Bengtsson T., Cannon B. Unexpected evidence for active brown adipose tissue in adult humans. (англ.) // American Journal Of Physiology. Endocrinology And Metabolism. — 2007. — August (vol. 293, no. 2). — P. 444—452. — doi:10.1152/ajpendo.00691.2006. — PMID 17473055.
- ↑ Saito M., Okamatsu-Ogura Y., Matsushita M., Watanabe K., Yoneshiro T., Nio-Kobayashi J., Iwanaga T., Miyagawa M., Kameya T., Nakada K., Kawai Y., Tsujisaki M. High incidence of metabolically active brown adipose tissue in healthy adult humans: effects of cold exposure and adiposity. (англ.) // Diabetes. — 2009. — July (vol. 58, no. 7). — P. 1526—1531. — doi:10.2337/db09-0530. — PMID 19401428.
- ↑ Graja A., Schulz T. J. Mechanisms of aging-related impairment of brown adipocyte development and function. (англ.) // Gerontology. — 2015. — Vol. 61, no. 3. — P. 211—217. — doi:10.1159/000366557. — PMID 25531079.
- ↑ Cedikova M., Kripnerová M., Dvorakova J., Pitule P., Grundmanova M., Babuska V., Mullerova D., Kuncova J. Mitochondria in White, Brown, and Beige Adipocytes. (англ.) // Stem Cells International. — 2016. — Vol. 2016. — P. 6067349—6067349. — doi:10.1155/2016/6067349. — PMID 27073398.
- ↑ Mescher, 2016, p. 123—124.
- ↑ Amitani M., Asakawa A., Amitani H., Inui A. The role of leptin in the control of insulin-glucose axis. (англ.) // Frontiers In Neuroscience. — 2013. — Vol. 7. — P. 51—51. — doi:10.3389/fnins.2013.00051. — PMID 23579596.
- ↑ Mescher, 2016, p. 124.
- ↑ Mescher, 2016, p. 124—125.
- ↑ Stallknecht B., Simonsen L., Bülow J., Vinten J., Galbo H. Effect of training on epinephrine-stimulated lipolysis determined by microdialysis in human adipose tissue. (англ.) // The American Journal Of Physiology. — 1995. — December (vol. 269, no. 6 Pt 1). — P. 1059—1066. — doi:10.1152/ajpendo.1995.269.6.E1059. — PMID 8572197.
- ↑ Spirovski M. Z., Kovacev V. P., Spasovska M., Chernick S. S. Effect of ACTH on lipolysis in adipose tissue of normal and adrenalectomized rats in vivo. (англ.) // The American Journal Of Physiology. — 1975. — February (vol. 228, no. 2). — P. 382—385. — doi:10.1152/ajplegacy.1975.228.2.382. — PMID 164126.
- ↑ Kiwaki K., Levine J. A. Differential effects of adrenocorticotropic hormone on human and mouse adipose tissue. (англ.) // Journal Of Comparative Physiology. B, Biochemical, Systemic, And Environmental Physiology. — 2003. — November (vol. 173, no. 8). — P. 675—678. — doi:10.1007/s00360-003-0377-1. — PMID 12925881.
- ↑ Romere C., Duerrschmid C., Bournat J., Constable P., Jain M., Xia F., Saha P. K., Del Solar M., Zhu B., York B., Sarkar P., Rendon D. A., Gaber M. W., LeMaire S. A., Coselli J. S., Milewicz D. M., Sutton V. R., Butte N. F., Moore D. D., Chopra A. R. Asprosin, a Fasting-Induced Glucogenic Protein Hormone. (англ.) // Cell. — 2016. — 21 April (vol. 165, no. 3). — P. 566—579. — doi:10.1016/j.cell.2016.02.063. — PMID 27087445.
- ↑ Wang H., Chu W. S., Hemphill C., Elbein S. C. Human resistin gene: molecular scanning and evaluation of association with insulin sensitivity and type 2 diabetes in Caucasians. (англ.) // The Journal Of Clinical Endocrinology And Metabolism. — 2002. — June (vol. 87, no. 6). — P. 2520—2524. — doi:10.1210/jcem.87.6.8528. — PMID 12050208.
- ↑ Guo L., Li Q., Wang W., Yu P., Pan H., Li P., Sun Y., Zhang J. Apelin inhibits insulin secretion in pancreatic beta-cells by activation of PI3-kinase-phosphodiesterase 3B. (англ.) // Endocrine Research. — 2009. — Vol. 34, no. 4. — P. 142—154. — doi:10.3109/07435800903287079. — PMID 19878074.
- ↑ MacDougald O. A., Burant C. F. The rapidly expanding family of adipokines. (англ.) // Cell Metabolism. — 2007. — September (vol. 6, no. 3). — P. 159—161. — doi:10.1016/j.cmet.2007.08.010. — PMID 17767903.
- ↑ Christiansen T., Richelsen B., Bruun J. M. Monocyte chemoattractant protein-1 is produced in isolated adipocytes, associated with adiposity and reduced after weight loss in morbid obese subjects. (англ.) // International Journal Of Obesity (2005). — 2005. — January (vol. 29, no. 1). — P. 146—150. — doi:10.1038/sj.ijo.0802839. — PMID 15520826.
- ↑ Katja Hoehn, Elaine N. Marieb. Anatomy & Physiology. — 3rd. — San Francisco, Calif. : Pearson/Benjamin Cummings, 2008. — ISBN 978-0-8053-0094-9.
- ↑ Stocco C. Tissue physiology and pathology of aromatase. (англ.) // Steroids. — 2012. — January (vol. 77, no. 1-2). — P. 27—35. — doi:10.1016/j.steroids.2011.10.013. — PMID 22108547.
- ↑ Mescher, 2016, p. 126—127.
- ↑ Fedorenko A., Lishko P. V., Kirichok Y. Mechanism of fatty-acid-dependent UCP1 uncoupling in brown fat mitochondria. (англ.) // Cell. — 2012. — 12 October (vol. 151, no. 2). — P. 400—413. — doi:10.1016/j.cell.2012.09.010. — PMID 23063128.
- ↑ Azzu V., Brand M. D. The on-off switches of the mitochondrial uncoupling proteins. (англ.) // Trends In Biochemical Sciences. — 2010. — May (vol. 35, no. 5). — P. 298—307. — doi:10.1016/j.tibs.2009.11.001. — PMID 20006514.
- ↑ Busiello R. A., Savarese S., Lombardi A. Mitochondrial uncoupling proteins and energy metabolism. (англ.) // Frontiers In Physiology. — 2015. — Vol. 6. — P. 36—36. — doi:10.3389/fphys.2015.00036. — PMID 25713540.
- ↑ 1 2 3 Mescher, 2016, p. 125.
- ↑ Gospodarska E., Nowialis P., Kozak L. P. Mitochondrial turnover: a phenotype distinguishing brown adipocytes from interscapular brown adipose tissue and white adipose tissue. (англ.) // The Journal Of Biological Chemistry. — 2015. — 27 March (vol. 290, no. 13). — P. 8243—8255. — doi:10.1074/jbc.M115.637785. — PMID 25645913.
- ↑ Rosenwald M., Perdikari A., Rülicke T., Wolfrum C. Bi-directional interconversion of brite and white adipocytes. (англ.) // Nature Cell Biology. — 2013. — June (vol. 15, no. 6). — P. 659—667. — doi:10.1038/ncb2740. — PMID 23624403.
- ↑ Lo K. A., Sun L. Turning WAT into BAT: a review on regulators controlling the browning of white adipocytes. (англ.) // Bioscience Reports. — 2013. — 6 September (vol. 33, no. 5). — doi:10.1042/BSR20130046. — PMID 23895241.
- ↑ Harms M. J., Ishibashi J., Wang W., Lim H. W., Goyama S., Sato T., Kurokawa M., Won K. J., Seale P. Prdm16 is required for the maintenance of brown adipocyte identity and function in adult mice. (англ.) // Cell Metabolism. — 2014. — 1 April (vol. 19, no. 4). — P. 593—604. — doi:10.1016/j.cmet.2014.03.007. — PMID 24703692.
- ↑ Wang W., Kissig M., Rajakumari S., Huang L., Lim H. W., Won K. J., Seale P. Ebf2 is a selective marker of brown and beige adipogenic precursor cells. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2014. — 7 October (vol. 111, no. 40). — P. 14466—14471. — doi:10.1073/pnas.1412685111. — PMID 25197048.
- ↑ Kissig M., Shapira S. N., Seale P. SnapShot: Brown and Beige Adipose Thermogenesis. (англ.) // Cell. — 2016. — 30 June (vol. 166, no. 1). — P. 258—258. — doi:10.1016/j.cell.2016.06.038. — PMID 27368105.
- ↑ Shapira S. N., Lim H. W., Rajakumari S., Sakers A. P., Ishibashi J., Harms M. J., Won K. J., Seale P. EBF2 transcriptionally regulates brown adipogenesis via the histone reader DPF3 and the BAF chromatin remodeling complex. (англ.) // Genes & Development. — 2017. — 1 April (vol. 31, no. 7). — P. 660—673. — doi:10.1101/gad.294405.116. — PMID 28428261.
- ↑ Boström P., Wu J., Jedrychowski M. P., Korde A., Ye L., Lo J. C., Rasbach K. A., Boström E. A., Choi J. H., Long J. Z., Kajimura S., Zingaretti M. C., Vind B. F., Tu H., Cinti S., Højlund K., Gygi S. P., Spiegelman B. M. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. (англ.) // Nature. — 2012. — 11 January (vol. 481, no. 7382). — P. 463—468. — doi:10.1038/nature10777. — PMID 22237023.
- ↑ Brestoff J. R., Kim B. S., Saenz S. A., Stine R. R., Monticelli L. A., Sonnenberg G. F., Thome J. J., Farber D. L., Lutfy K., Seale P., Artis D. Group 2 innate lymphoid cells promote beiging of white adipose tissue and limit obesity. (англ.) // Nature. — 2015. — 12 March (vol. 519, no. 7542). — P. 242—246. — doi:10.1038/nature14115. — PMID 25533952.
- ↑ Mescher, 2016, p. 127.
- ↑ Obesity and overweight Fact sheet N°311. WHO (январь 2015). Дата обращения: 2 февраля 2016. Архивировано 22 апреля 2018 года.
- ↑ Singh A. K., Corwin R. D., Teplitz C., Karlson K. E. Consecutive repair of complex congenital heart disease using hypothermic cardioplegic arrest--its results and ultrastructural study of the myocardium. (англ.) // The Thoracic And Cardiovascular Surgeon. — 1984. — February (vol. 32, no. 1). — P. 23—26. — doi:10.1055/s-2007-1023339. — PMID 6198769.
- ↑ Elmquist J. K., Maratos-Flier E., Saper C. B., Flier J. S. Unraveling the central nervous system pathways underlying responses to leptin. (англ.) // Nature Neuroscience. — 1998. — October (vol. 1, no. 6). — P. 445—450. — doi:10.1038/2164. — PMID 10196541.
- ↑ Morris D. L., Rui L. Recent advances in understanding leptin signaling and leptin resistance. (англ.) // American Journal Of Physiology. Endocrinology And Metabolism. — 2009. — December (vol. 297, no. 6). — P. 1247—1259. — doi:10.1152/ajpendo.00274.2009. — PMID 19724019.
- ↑ Giordano Antonio, Frontini Andrea, Cinti Saverio. Convertible visceral fat as a therapeutic target to curb obesity (англ.) // Nature Reviews Drug Discovery. — 2016. — 11 March (vol. 15, no. 6). — P. 405—424. — ISSN 1474-1776. — doi:10.1038/nrd.2016.31.
- ↑ Sugii S., Kida Y., Kawamura T., Suzuki J., Vassena R., Yin Y. Q., Lutz M. K., Berggren W. T., Izpisúa Belmonte J. C., Evans R. M. Human and mouse adipose-derived cells support feeder-independent induction of pluripotent stem cells. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2010. — 23 February (vol. 107, no. 8). — P. 3558—3563. — doi:10.1073/pnas.0910172106. — PMID 20133714.
- ↑ 1 2 Atzmon G., Yang X. M., Muzumdar R., Ma X. H., Gabriely I., Barzilai N. Differential gene expression between visceral and subcutaneous fat depots. (англ.) // Hormone And Metabolic Research = Hormon- Und Stoffwechselforschung = Hormones Et Metabolisme. — 2002. — November (vol. 34, no. 11-12). — P. 622—628. — doi:10.1055/s-2002-38250. — PMID 12660871.
- ↑ Baglioni S., Cantini G., Poli G., Francalanci M., Squecco R., Di Franco A., Borgogni E., Frontera S., Nesi G., Liotta F., Lucchese M., Perigli G., Francini F., Forti G., Serio M., Luconi M. Functional differences in visceral and subcutaneous fat pads originate from differences in the adipose stem cell. (англ.) // PloS One. — 2012. — Vol. 7, no. 5. — P. e36569—36569. — doi:10.1371/journal.pone.0036569. — PMID 22574183.
- ↑ Russo V., Yu C., Belliveau P., Hamilton A., Flynn L. E. Comparison of human adipose-derived stem cells isolated from subcutaneous, omental, and intrathoracic adipose tissue depots for regenerative applications. (англ.) // Stem Cells Translational Medicine. — 2014. — February (vol. 3, no. 2). — P. 206—217. — doi:10.5966/sctm.2013-0125. — PMID 24361924.
- ↑ Yu, P., Yuan, R., Yang, X., & Qi, Z. Adipose tissue, aging, and metabolism (англ.) // Current Opinion in Endocrine and Metabolic Research. — 2019. — Vol. 5. — P. 11—20. — doi:10.1016/j.coemr.2019.02.003.
- ↑ Ibrahim, M. M. Subcutaneous and visceral adipose tissue: structural and functional differences (англ.) // Obesity reviews. — 2010. — Vol. 11, iss. 1. — P. 11—18. — doi:10.1111/j.1467-789X.2009.00623.x. — PMID 19656312.
- ↑ Bouchi, R., Takeuchi, T., et al. High visceral fat with low subcutaneous fat accumulation as a determinant of atherosclerosis in patients with type 2 diabetes (англ.) // Cardiovascular diabetology. — 2015. — Vol. 14, iss. 1, no. 136. — P. 1—7. — doi:10.1186/s12933-015-0302-4. — PMID 26445876. — PMC 4597374. Архивировано 19 декабря 2022 года.
- ↑ Akazawa, N., et al. Relationship Between Aging and Intramuscular Adipose Tissue in Older Inpatients (англ.) // Journal of the American Medical Directors Association. — 2020. — Vol. 22, iss. 6. — P. 1287—1291. — doi:10.1016/j.jamda.2020.09.017. — PMID 33127329.
- ↑ Nguyen H. P., et al. Aging-dependent regulatory cells emerge in subcutaneous fat to inhibit adipogenesis (англ.) // Developmental Cell. — 2021. — Vol. 56, iss. 10. — P. 1437—1451. — doi:10.1016/j.devcel.2021.03.026.
- ↑ Hu, Y., et al. Deletion of Nrip1 delays skin aging by reducing adipose-derived mesenchymal stem cells (ADMSCs) senescence, and maintaining ADMSCs quiescence (англ.) // GeroScience. — 2021. — Vol. 43, iss. 4. — P. 1815—1833. — doi:10.1007/s11357-021-00344-y. — PMID 33704619. Архивировано 19 декабря 2022 года.
- ↑ Cannon B., Nedergaard J. Developmental biology: Neither fat nor flesh. (англ.) // Nature. — 2008. — 21 August (vol. 454, no. 7207). — P. 947—948. — doi:10.1038/454947a. — PMID 18719573.
- ↑ Lee Paul, Swarbrick Michael M., Ho Ken K. Y. Brown Adipose Tissue in Adult Humans: A Metabolic Renaissance (англ.) // Endocrine Reviews. — 2013. — 1 June (vol. 34, no. 3). — P. 413—438. — ISSN 0163-769X. — doi:10.1210/er.2012-1081.
- ↑ Lafontan Max. Historical perspectives in fat cell biology: the fat cell as a model for the investigation of hormonal and metabolic pathways (англ.) // American Journal of Physiology-Cell Physiology. — 2012. — 15 January (vol. 302, no. 2). — P. C327—C359. — ISSN 0363-6143. — doi:10.1152/ajpcell.00168.2011.
- ↑ Krug A. W., Ehrhart-Bornstein M. Newly discovered endocrine functions of white adipose tissue: possible relevance in obesity-related diseases. (англ.) // Cellular And Molecular Life Sciences : CMLS. — 2005. — June (vol. 62, no. 12). — P. 1359—1362. — doi:10.1007/s00018-005-4555-z. — PMID 15924267.
Литература
- Афанасьев Ю. И., Кузнецов С. Л., Юрина Н. А., Котовский Е. Ф. и др. Гистология, цитология и эмбриология. — 6-е изд., перераб. и доп.. — М.: Медицина, 2004. — 768 с. — ISBN 5-225-04858-7.
- Anthony L. Mescher. Junqueira's Basic Histology. — McGraw-Hill Education, 2016. — ISBN 978-0-07-184270-9.