Изомери́я — явление, заключающееся в существовании химических соединений — изомеров, — одинаковых по атомному составу и молекулярной массе, но различающихся по строению или расположению атомов в пространстве и, вследствие этого, по свойствам.

Ферме́нты, или энзи́мы , — обычно сложные белковые соединения, РНК (рибозимы) или их комплексы, ускоряющие химические реакции в живых системах. Каждый фермент, свернутый в определённую структуру, ускоряет соответствующую химическую реакцию: реагенты в такой реакции называются субстратами, а получающиеся вещества — продуктами. Ферменты специфичны к субстратам: АТФ-аза катализирует расщепление только АТФ, а киназа фосфорилазы фосфорилирует только фосфорилазу.
Трансфера́зы (КФ2) — отдельный класс ферментов, катализирующих перенос функциональных групп и молекулярных остатков от одной молекулы к другой. Широко распространены в растительных и животных организмах, участвуют в превращениях углеводов, липидов, нуклеиновых и аминокислот.
Лиа́зы (КФ4) — отдельный класс ферментов, катализирующих реакции негидролитического и неокислительного разрыва различных химических связей субстрата, обратимые реакции образования и разрыва двойных связей, сопровождающиеся отщеплением или присоединением групп атомов по её месту, а также образованием циклических структур.

Алкогольдегидрогеназа (Алкоголь: НАД-оксидоредуктаза, КФ 1.1.1.1) — фермент класса дегидрогеназ, катализирующий окисление спиртов и ацеталей до альдегидов и кетонов в присутствии никотинамид-аденин-динуклеотида (НАД). Алкогольдегидрогеназы (алкоголь-НАД+оксидоредуктазы) являются димерами, состоящими из субъединиц с молекулярным весом около 40000 и содержащими ион цинка Zn2+.
Лигаза — фермент, катализирующий соединение двух молекул с образованием новой химической связи — лигирование. При этом обычно происходит отщепление (гидролиз) небольшой химической группы от одной из молекул.
Оксидоредукта́зы (КФ1) — отдельный класс ферментов, катализирующих лежащие в основе биологического окисления реакции, сопровождающиеся переносом электронов с одной молекулы на другую.
Шифр КФ, или код фермента, англ. EC number — классификационный номер фермента по международной иерархической классификации. Принятая система классифицирует ферменты по группам и индексирует индивидуальные ферменты, что важно для стандартизации исследований.
Гидрола́зы (КФ 3) — это класс ферментов, катализирующий гидролиз ковалентной связи. Общий вид реакции, катализируемой гидролазой, выглядит следующим образом:
- A-B + H2O → A-OH + B-H
Рибонуклеазы — ферменты-нуклеазы, катализирующие деградацию РНК. Рибонуклеазы классифицируют на эндорибонуклеазы и экзорибонуклеазы. К рибонуклеазам относят некоторые подклассы КФ 2.7 и КФ 3.1.

Поли(АДФ-рибоза)-полимеразы (PARP) — ферменты, катализирующие поли-АДФ-рибозилирование, один из видов посттрансляционной модификации белков.

Сирогем — гемоподобная простетическая группа нитрит- и сульфитредуктаз, катализирующих шестиэлектронное восстановление серы сульфита или азота нитрита до сульфида и аммиака.

Альдолаза, также фруктозобисфосфат-(фруктозодифосфат)-альдолаза, реже альдолаза А — фермент из семейства альдолазы, катализирующий реакцию негидролитического расщепления связей C-С в молекуле фруктозо-1,6-дифосфата, в результате которой образуются дигидроксиацетонфосфат и глицеральдегид-3-фосфат в процессе гликолиза, а также обратную реакцию альдольной конденсации, происходящей в глюконеогенезе, по схеме:
Арабино́зный оперо́н — бактериальный оперон, кодирующий белки, необходимые для метаболизма арабинозы. Он подчиняется как позитивному, так и негативному контролю: экспрессию оперона стимулирует субстрат, то есть арабиноза, но при наличии глюкозы в среде его экспрессия подавлена. В состав арабинозного оперона входит три структурных гена: araB, araA и araD, а также регуляторный ген araC, сцеплённый со структурными генами, и регуляторные участки. Арабинозный оперон был открыт и исследован у кишечной палочки в 1970-х.
Бе́та-окисле́ние (β-окисление), также цикл Кноопа — Линена, — метаболический процесс деградации жирных кислот. Своё название процесс получил по 2-му углеродному атому (С-3 или β-положение) от карбоксильной группы (-СООН) жирной кислоты, который подвергается окислению и последовательному отделению от молекулы. Продуктами каждого цикла β-окисления являются ФАДH2, НАДH и ацетил-КоА. Реакции β-окисления и последующего окисления ацетил-КоА в цикле Кребса служат одним из основных источников энергии для синтеза АТФ по механизму окислительного фосфорилирования.
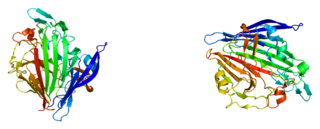
Галактозмутаротаза, также альдоза-1-эпимераза — фермент из семейства эпимераз, которая катализирует превращение бета-D-галактозы в альфа-D-галактозу процессом мутаротации. Ген, кодирующий данный фермент GALM, локазизован во 2-й хромосоме.
Сукцинил-Кофермент А, сокращенно сукцинил-КоА, представляет собой тиоэфир дикарбоновой янтарной кислоты и кофермента А. Принимает участие во многих биохимических путях, в частности, является интермедиатом цикла Кребса. Служит предшественником синтеза δ-аминолевулиновой кислоты, являющейся первым специфическим интермедиатом в синтезе порфиринов.
Транслоказы — отдельный класс ферментов, катализирующих перенос ионов или молекул через мембраны или их разделение в мембранах. Этот класс ферментов сформировался из ранее принадлежащих другим классам ферментов.
Фукозилтрансферазы — ферменты, переносящие L-фукозу с субстрата-донора ГДФ-фукозы на субстрат-акцептор. Акцептором может быть другой сахарид, такой как остаток N-ацетилглюкозамина полисахаридной цепи, что приводит к N-гликозилированию, либо белок, что приводит к O-гликозилированию под действием O-фукозилтрансферазы. У млекопитающих существует несколько фукозилтрансфераз, большинство из которых локализуются на мембранах аппарата Гольджи. O-Фукозилтрансферазы находятся на эндоплазматическом ретикулуме (ЭР).

6-Фосфоглюконолактоназа — цитозольный фермент, обнаруженный во всех организмах, который катализирует гидролиз 6-фосфоглюконолактона до 6-фосфоглюконовой кислоты в окислительной фазе пентозофосфатного пути. Третичная структура 6PGL использует α/β гидролазную складку с остатками активного сайта, сгруппированными на петлях α-спиралей. Основываясь на кристаллической структуре фермента, предполагается, что механизм зависит от переноса протона остатком гистидина в активном центре. 6PGL избирательно катализирует гидролиз δ-6-фосфоглюконолактона и не проявляет активности в отношении γ-изомера.