Инактивация X-хромосомы


Инактивация X-хромосомы (англ. X-inactivation, lyonization) — эпигенетический процесс, в результате которого во время раннего эмбрионального развития одна из двух копий X-хромосом, представленных в клетках самок млекопитающих, становится транскрипционно молчащей. ДНК неактивной X-хромосомы при этом формирует факультативный гетерохроматин.
Инактивация X-хромосомы происходит в клетках самок млекопитающих для того, чтобы с двух копий X-хромосом не образовывалось вдвое больше продуктов соответствующих генов, чем у самцов млекопитающих. Такой процесс называется дозовой компенсацией генов. У плацентарных выбор X-хромосомы, которая будет инактивирована, случаен (что показано для клеток мышей и человека). Инактивированная X-хромосома будет оставаться неактивной во всех последующих дочерних клетках, образующихся в результате деления.
История
В 1959 году Сусуму Оно показал, что одна из двух X-хромосом у самок вела себя как аутосома, а другая находилась в состоянии гетерохроматина.[2] Двумя группами исследователей независимо друг от друга было предположено, что одна из двух X-хромосом подвергается инактивации. В 1961 году Мэри Лайон предположила, что инактивация одной из X-хромосом у самок обуславливает пятнистую окраску шерсти мышей у особей, гетерозиготных по генам окраски.[3] Гипотеза Лайон об инактивации одной X-хромосомы в клетках самок млекопитающих объясняла также тот факт, что мыши лишь с одной X-хромосомой имеют фенотип самки. Эрнест Бейтлер (англ. Ernest Beutler), изучая гетерозиготных самок, дефицитных по ферменту дегидрогеназе глюкозо-6-фосфатазы, независимо от Лайон, предположил существование у гетерозиготных организмов двух типов эритроцитов — дефицитных по ферменту и нормальных.[4]
Механизм
На стадии двух или четырёхклеточного зародыша мыши происходит инактивация отцовской X-хромосомы по механизму импринтинга.[5][6][7] На стадии ранней бластоцисты в клетках внутренней клеточной массы становятся активными обе X-хромосомы. Затем во всех клетках внутренней клеточной массы бластоцисты необратимо и независимо друг от друга инактивируется одна из X-хромосом. Инактивированная на этой ранней стадии развития зародыша, X-хромосома затем будет инактивирована во всех соматических клетках-потомках этой клетки. Инактивация X-хросомомы снимается в клетках зародышевого пути самки, и поэтому все ооциты содержат обе активные X-хромосомы.
Показано, что нормальным состоянием для X-хромосомы в клетках млекопитающих является инактивированное состояние, так как в организмах или клетках, содержащих более, чем две X-хромосомы, активной является лишь одна, в то время как остальные X-хромосомы неактивны. Показано наличие на X-хромосомах участка центр инактивации X-хромосомы — XIC (от англ. X inactivation center). Наличие центра инактивации необходимо и достаточно для инактивации X-хромосомы. Транслокация участка хромосомы, содержащего XIC на аутосому, приводит к инактивации соответствующей аутосомы, в то время как X-хромосомы, не имеющие XIC, остаются активными.

Участок XIC содержит два гена некодирующих РНК, которые не транслируются в белок — Xist и Tsix, принимающие участие в инактивации X-хромосомы. Также XIC содержит сайты связывания соответствующих регуляторных белков.
Некодирующие РНК Xist и Tsix
Xist представляет собой ген, кодирующий молекулу длинной некодирующей РНК, который опосредует специфическую инактивацию той X-хромосомы, с которой он был транскрибирован.[8] Неактивная X-хромосома покрыта РНК, транскрибированной с гена Xist,[9] активная X-хромосома такой РНК не покрыта. Ген Xist экспрессируется только с неактивной X-хромосомы, хромосомы, не содержащие ген Xist, не могут быть инактивированы.[10] Искусственное перемещение гена Xist на другие хромосомы и его экспрессия, приводит к инактивации других хромосом.[11][12]
До момента инактивации обе X-хромосомы слабо экспрессируют РНКовый продукт гена Xist, в ходе процесса инактивации экспрессия Xist на активной хромосоме снижается, а на инактивированной — увеличивается. Продукт гена Xist постепенно покрывает инактивированную хромосому, начиная от участка XIC;[11] Сайленсинг генов инактивированной хромосомы начинается вскоре после того, как хромосома становится покрытой транскриптом гена Xist.
Tsix представляет собой длинную молекулу РНК, не кодирующую белок. Транскрипт Tsix является антисмысловым к транскрипту гена Xist, то есть транскрибируется с противоположной цепи ДНК того же гена.[13] Tsix является негативным регулятором Xist; X-хромосомы, не экспрессирующие Tsix и имеющие поэтому повышенный уровень экспрессии Xist, инактивированы намного чаще, чем обычные хромосомы.
Как и в случае с геном Xist, перед инактивацией обе X-хромосомы слабо экспрессируют РНК гена Tsix с соответствующего гена. После начала инактивации X-хромосомы будущая инактивированная хромосома прекращает экспрессировать РНК Tsix, в то время как активная хромосома продолжает экспрессировать Tsix ещё несколько дней.
Тельце Барра
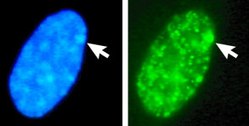
Тельцем Барра называют X-хромосому, ДНК которой находится в состоянии гетерохроматина.[14] Тельце Барра содержит продукт гена Xist, обычно располагается на периферии ядра, ДНК тельца Барра поздно реплицируется.
Примечания
- ↑ Gartler S. M., Varadarajan K. R., Luo P., Canfield T. K., Traynor J., Francke U., Hansen R. S. Normal histone modifications on the inactive X chromosome in ICF and Rett syndrome cells: implications for methyl-CpG binding proteins (англ.) // BMC Biology[англ.] : journal. — 2004. — Vol. 2. — P. 21. — doi:10.1186/1741-7007-2-21. Архивировано 25 мая 2011 года. — Figure 1 Архивная копия от 8 августа 2010 на Wayback Machine
- ↑ Ohno S., Kaplan W. D., Kinosita R. Formation of the sex chromatin by a single X-chromosome in liver cells of rattus norvegicus (англ.) // Exp Cell Res[англ.] : journal. — 1959. — Vol. 18. — P. 415—419. — doi:10.1016/0014-4827(59)90031-X. — PMID 14428474.
- ↑ Lyon M. F. Gene Action in the X-chromosome of the Mouse (Mus musculus L.) (англ.) // Nature : journal. — 1961. — Vol. 190, no. 4773. — P. 372—373. — doi:10.1038/190372a0. — PMID 13764598. Архивировано 17 февраля 2021 года.
- ↑ Beutler E., Yeh M., Fairbanks V. F. The normal human female as a mosaic of X-chromosome activity: Studies using the gene for G-6-PD-deficiency as a marker (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1962. — January (vol. 48). — P. 9—16. — PMID 13868717. — PMC 285481.
- ↑ Takagi N., Sasaki M. Preferential inactivation of the paternally derived X chromosome in the extraembryonic membranes of the mouse (англ.) // Nature : journal. — 1975. — Vol. 256. — P. 640—642. — doi:10.1038/256640a0. — PMID 1152998. Архивировано 6 августа 2010 года.
- ↑ Cheng M. K., Disteche C. M. Silence of the fathers: early X inactivation (англ.) // BioEssays[англ.] : journal. — 2004. — Vol. 26, no. 8. — P. 821—824. — doi:10.1002/bies.20082. — PMID 15273983. (недоступная ссылка)
- ↑ Okamoto I., Otte A., Allis C., Reinberg D., Heard E. Epigenetic dynamics of imprinted X inactivation during early mouse development (англ.) // Science : journal. — 2004. — Vol. 303, no. 5658. — P. 644—649. — doi:10.1126/science.1092727. — PMID 14671313.
- ↑ Hoki Y., Kimura N., Kanbayashi M., Amakawa Y., Ohhata T., Sasaki H., Sado T. A proximal conserved repeat in the Xist gene is essential as a genomic element for X-inactivation in mouse (англ.) // Development : journal. — 2009. — Vol. 136. — P. 139—146. — doi:10.1242/dev.026427. — PMID 19036803. Архивировано 26 июня 2009 года.
- ↑ Ng K., Pullirsch D., Leeb M., Wutz A. Xist and the order of silencing (англ.) // EMBO Rep[англ.] : journal. — 2007. — Vol. 8. — P. 34—9. — doi:10.1038/sj.embor.7400871. — PMID 17203100. — PMC 1796754.. — «Figure 1 Xist RNA encompasses the X from which it is transcribed.».
- ↑ Penny G. D., Kay G. F., Sheardown S. A., Rastan S., Brockdorff N. Requirement for Xist in X chromosome inactivation (англ.) // Nature. — 1996. — Vol. 379. — P. 116—117. — doi:10.1038/379131a0. — PMID 8538762.
- ↑ 1 2 Herzing L. B., Romer J. T., Horn J. M., Ashworth A. Xist has properties of the X-chromosome inactivation centre (англ.) // Nature : journal. — 1997. — Vol. 386. — P. 272—275. — doi:10.1038/386272a0. — PMID 9069284.
- ↑ Lee J. T., Jaenisch R. Long-range cis effects of ectopic X-inactivation centres on a mouse autosome (англ.) // Nature : journal. — 1997. — Vol. 386. — P. 275—279. — doi:10.1038/386275a0. — PMID 9069285.
- ↑ Lee J. T., Davidow L. S., Warshawsky D. Tisx, a gene antisense to Xist at the X-inactivation centre (англ.) // Nat Genet : journal. — 1999. — Vol. 21. — P. 400—404. — doi:10.1038/7734. Архивировано 6 августа 2010 года.
- ↑ Barr M. L., Bertram E. G. A Morphological Distinction between Neurones of the Male and Female, and the Behaviour of the Nucleolar Satellite during Accelerated Nucleoprotein Synthesis (англ.) // Nature : journal. — 1949. — Vol. 163, no. 4148. — P. 676—677. — doi:10.1038/163676a0.
Литература
- Шевченко А. И., Захарова И. С., Закиян С. М. Эволюционный путь процесса инактивации X-хромосомы у млекопитающих // Acta Naturae. — 2013. — Т. 5, № 2. — С. 40—54. (недоступная ссылка)
- Huynh K. D., Lee J. T. X-chromosome inactivation: a hypothesis linking ontogeny and phylogeny (англ.) // Nature Rev Genet : journal. — 2005. — Vol. 9, no. 5. — P. 41—8. — doi:10.1038/nrg1604. — PMID 15818384.
- X-inactivation as a possible cause for autoimmunity
- Goto T., Monk M. Regulation of X-chromosome inactivation in development in mice and humans (англ.) // Microbiology and Molecular Biology Reviews[англ.] : journal. — American Society for Microbiology[англ.], 1998. — 1 June (vol. 62, no. 2). — P. 362—378. — PMID 9618446.
- Lyon M. The Lyon and the LINE hypothesis (англ.) // Semin Cell Dev Biol[англ.] : journal. — 2003. — Vol. 14, no. 6. — P. 313—318. — doi:10.1016/j.semcdb.2003.09.015. — PMID 15015738.
- Ng K., Pullirsch D., Leeb M., Wutz A. Xist and the order of silencing (англ.) // EMBO Rep[англ.] : journal. — 2007. — Vol. 8. — P. 34—9. — doi:10.1038/sj.embor.7400871. — PMID 17203100. — PMC 1796754.











