Инозитолтрифосфат
| Инозитолтрифосфат | |
|---|---|
 | |
| Общие | |
| Хим. формула | C6H15O15P3 |
| Классификация | |
| Рег. номер CAS | 2068-89-5 и 88269-39-0 |
| PubChem | 55310 |
| SMILES | |
| InChI | |
| ChEBI | 191121 |
| ChemSpider | 49951 |
| Приведены данные для стандартных условий (25 °C, 100 кПа), если не указано иное. | |
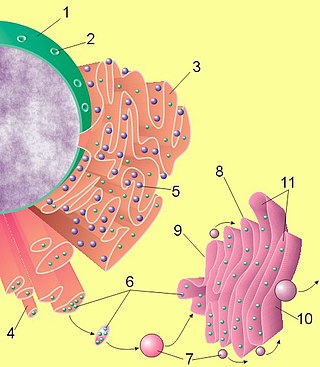
Инозитолтрифосфат или инозитол-1,4,5-трифосфат, сокращенно InsP 3 или Ins3P или IP3, представляет собой сигнальную молекулу инозитолфосфата. Он производится путем гидролиза фосфатидилинозитол-4,5-бисфосфата (PIP2), фосфолипида, расположенного в плазматической мембране, фосфолипазой C (PLC). Вместе с диацилглицерином (DAG) IP3 представляет собой молекулу вторичного мессенджера, используемую для передачи сигналов в биологических клетках. Пока DAG остается внутри мембраны, IP3 растворим и диффундирует через клетку, где связывается со своим рецептором, который представляет собой кальциевый канал, расположенный в эндоплазматическом ретикулуме. Когда IP3 связывается с рецептором, кальций высвобождается в цитозоль, тем самым активируя различные внутриклеточные сигналы, регулируемые кальцием.
Характеристики
Химическая формула и молекулярная масса
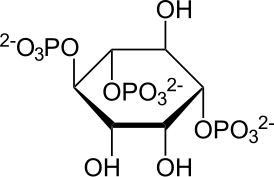
IP3 представляет собой органическую молекулу с молекулярной массой 420,10 г/моль. Его эмпирическая формула — C6H15O15P3. Он состоит из инозитолового кольца с тремя фосфатными группами, связанными в положениях углерода 1, 4 и 5, и тремя гидроксильными группами, связанными в положениях 2, 3 и 6[1].
Химические свойства
Стыковка IP3 с его рецептором, который называется рецептором инозитолтрифосфата (InsP3R), впервые была изучена с помощью делеционного мутагенеза в начале 1990-х годов.[2] Исследования были сосредоточены на N-концевой стороне рецептора IP3. В 1997 году исследователи локализовали область рецептора IP3, участвующую в связывании IP3, между аминокислотными остатками 226 и 578 в 1997 году. Учитывая, что IP3 представляет собой отрицательно заряженную молекулу, предполагалось, что в этом участвуют положительно заряженные аминокислоты, такие как аргинин и лизин. Было обнаружено, что два остатка аргинина в положениях 265 и 511 и один остаток лизина в положении 508 являются ключевыми в докинге IP3. Используя модифицированную форму IP3, было обнаружено, что все три фосфатные группы взаимодействуют с рецептором, но не одинаково. Фосфаты в 4-м и 5-м положениях взаимодействуют более интенсивно, чем фосфат в 1-м положении и гидроксильная группа в 6-м положении инозитольного кольца[3].
Связывание с рецептором

Фосфатные группы могут существовать в трех различных формах в зависимости от pH раствора. Атомы фосфора могут связывать три атома кислорода одинарными связями и четвёртый атом кислорода с помощью двойной/дативной связи. pH раствора и, следовательно, форма фосфатной группы определяют её способность связываться с другими молекулами. Связывание фосфатных групп с инозитольным кольцом осуществляется путем связывания эфиров фосфора (см. Фосфорные кислоты и фосфаты). Эта связь включает объединение гидроксильной группы инозитолового кольца и свободной фосфатной группы посредством реакции дегидратации. Учитывая, что среднее физиологическое значение pH составляет примерно 7,4, основной формой фосфатных групп, связанных с инозитоловым кольцом in vivo, является PO42- . Это придает IP3 чистый отрицательный заряд, что важно для возможности его присоединения к рецептору посредством связывания фосфатных групп с положительно заряженными остатками на рецепторе. IP3 имеет три донора водородных связей в виде трех гидроксильных групп. Гидроксильная группа на 6-м атоме углерода в инозитольном кольце также участвует в стыковке IP3[4].
Открытие
Открытие того, что гормон может влиять на метаболизм фосфоинозитидов, было сделано Мейбл Р. Хокин (1924—2003) и её мужем Лоуэллом Э. Хокином в 1953 году, когда они обнаружили, что радиоактивный фосфат 32Р включается в фосфатидилинозитол срезов поджелудочной железы при стимуляции ацетилхолин. До этого считалось, что фосфолипиды являются инертными структурами, используемыми клетками только в качестве строительных блоков для строительства плазматической мембраны[5].
В течение следующих 20 лет мало что было обнаружено о важности метаболизма PIP2 с точки зрения передачи клеточных сигналов, пока в середине 1970-х годов Роберт Х. Мичелл не выдвинул гипотезу о связи между катаболизмом PIP2 и увеличением внутриклеточного кальция (Ca2+) уровни. Он предположил, что активируемый рецептором гидролиз PIP 2 приводит к образованию молекулы, которая вызывает увеличение внутриклеточной мобилизации кальция[6]. Эта идея была тщательно исследована Мичеллом и его коллегами, которые в 1981 году смогли показать, что PIP2 гидролизуется до DAG и IP3 неизвестной тогда фосфодиэстеразой. В 1984 году было обнаружено, что IP3 действует как вторичный мессенджер, способный перемещаться через цитоплазму в эндоплазматический ретикулум, где он стимулирует высвобождение кальция в цитоплазму[7].
Дальнейшие исследования предоставили ценную информацию о пути IP3, например, открытие в 1986 году того, что одна из многих ролей кальция, высвобождаемого IP3, заключается в работе с DAG для активации протеинкиназы C (PKC)[8]. В 1989 году было обнаружено, что фосфолипаза C (PLC) является фосфодиэстеразой, ответственной за гидролиз PIP2 до DAG и IP3[9]. Сегодня сигнальный путь IP3 хорошо изучен и известен как важный в регуляции множества кальций-зависимых клеточных сигнальных путей.
Сигнальный путь

Увеличение внутриклеточной концентрации Ca2+ часто является результатом активации IP3 . Когда лиганд связывается с рецептором, связанным с G-белком (GPCR), который связан с гетеротримерным G-белком Gq, α-субъединица Gq может связываться с изозимом PLC PLC-β и индуцировать активность, что приводит к расщеплению PIP2 в IP3 и DAG[10].
Если в активации этого пути участвует рецепторная тирозинкиназа (RTK), изофермент PLC-γ имеет остатки тирозина, которые могут фосфорилироваться при активации RTK, и это активирует PLC-γ и позволяет ему расщеплять PIP2 до DAG и IP3. Это происходит в клетках, которые способны реагировать на факторы роста, такие как инсулин, поскольку факторы роста являются лигандами, ответственными за активацию RTK[11].
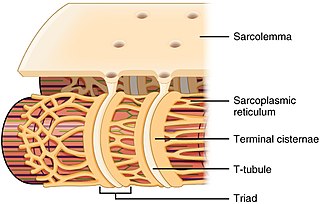
IP3 (также сокращенно Ins(1,4,5)P3) представляет собой растворимую молекулу и способен диффундировать через цитоплазму в эндоплазматический ретикулум или саркоплазматический ретикулум в случае мышечных клеток, как только он был продуцирован под действием PLC. Попав в эндоплазматический ретикулум, IP3 способен связываться с рецептором Ins(1,4,5)P3 — Ins(1,4,5)P3R, который представляет собой лиганд-управляемый Ca2+ — канал, который находится на поверхностb эндоплазматического ретикулума. Связывание IP3 (в данном случае лиганда) с Ins(1,4,5)P3R запускает открытие канала Ca2+ и, таким образом, высвобождение Ca2+ в цитоплазму[11]. В клетках сердечной мышцы это увеличение Ca2+ активирует канал саркоплазматического ретикулума, управляемый рианодиновым рецептором, что приводит к дальнейшему увеличению Ca2+ посредством процесса, известного как кальций-индуцированное высвобождение кальция. IP3 может также активировать каналы Ca2+ на клеточной мембране косвенно, увеличивая внутриклеточную концентрацию Ca2+[10].
Функция
Человек
Основные функции IP3 заключаются в мобилизации Ca2+ из запасающих органелл и регулировании пролиферации клеток и других клеточных реакциях, требующих свободного кальция. Например, в гладкомышечных клетках увеличение концентрации цитоплазматического Са2+ приводит к сокращению мышечной клетки[12].
В нервной системе IP3 служит вторичным мессенджером, при этом мозжечок содержит самую высокую концентрацию рецепторов IP3[13]. Имеются данные о том, что рецепторы IP3 играют важную роль в индукции пластичности клеток Пуркинье мозжечка[14].
Яйца морских ежей
Медленный блок полиспермии у морского ежа опосредуется системой вторичного мессенджера PIP2. Активация связывающих рецепторов активирует PLC, который расщепляет PIP2 в плазматической мембране яйцеклетки, высвобождая IP3 в цитоплазму яйцеклетки. IP3 диффундирует в эдоплазматический ретикулум, где открывает каналы Ca2+.
Нарушения обмена
Болезнь Хантингтона
Болезнь Хантингтона возникает, когда цитозольный белок Хантингтин (Htt) имеет дополнительные 35 остатков глутамина, добавленных к его аминоконцевой области. Эта модифицированная форма Htt называется Httexp . Httexp делает рецепторы IP3 типа 1 более чувствительными к IP3, что приводит к высвобождению слишком большого количества Ca2+ из эндоплазматического ретикулума. Высвобождение Ca2+ вызывает увеличение цитозольной и митохондриальной концентрации Ca2+. Считается, что это увеличение Ca2+ является причиной ГАМКергической деградации MSN[15].
Болезнь Альцгеймера
Болезнь Альцгеймера включает в себя прогрессирующую дегенерацию мозга, серьёзно влияющую на умственные способности[16]. С тех пор как в 1994 году была предложена Ca2+-гипотеза болезни Альцгеймера, несколько исследований показали, что нарушения передачи сигналов Ca2+ являются основной причиной болезни Альцгеймера. Семейная болезнь Альцгеймера тесно связана с мутациями в генах пресенилина 1 (PS1), пресенилина 2 (PS2) и белка-предшественника амилоида (APP). Было обнаружено, что все мутированные формы этих генов, наблюдаемые на сегодняшний день, вызывают аномальную передачу сигналов Ca2+ в эндоплазматический ретикулум. Было показано, что мутации в PS1 увеличивают IP3 -опосредованное высвобождение Ca2+ из эндоплазматического ретикулума на нескольких моделях животных. Блокаторы кальциевых каналов с некоторым успехом использовались для лечения болезни Альцгеймера, а использование лития для уменьшения обмена IP3 также было предложено в качестве возможного метода лечения[17][18].
См. также
Рекомендации
- ↑ PubChem 439456
- ↑ Mignery, GA (1990). "The ligand binding site and transduction mechanism in the inositol-1,4,5-triphosphate receptor". The EMBO Journal. 9 (12): 3893—8. doi:10.1002/j.1460-2075.1990.tb07609.x. PMID 2174351.
- ↑ Taylor, Colin W. (2004). "IP3 receptors: The search for structure" (PDF). Trends in Biochemical Sciences. 29 (4): 210—9. doi:10.1016/j.tibs.2004.02.010. PMID 15082315. Архивировано из оригинала (PDF) 8 августа 2017. Дата обращения: 13 декабря 2023.
- ↑ Bosanac, Ivan (2004). "Structural insights into the regulatory mechanism of IP3 receptor". Biochimica et Biophysica Acta (BBA) - Molecular Cell Research. 1742 (1—3): 89—102. doi:10.1016/j.bbamcr.2004.09.016. PMID 15590059.
- ↑ Hokin, LE (1953). "Enzyme secretion and the incorporation of 32P into phosphlipids of pancreas slices". Journal of Biological Chemistry. 203 (2): 967—977. doi:10.1016/S0021-9258(19)52367-5. PMID 13084667.
- ↑ Michell, RH (1975). "Inositol phospholipids and cell surface receptor function". Biochimica et Biophysica Acta (BBA) - Reviews on Biomembranes. 415 (1): 81—147. doi:10.1016/0304-4157(75)90017-9. PMID 164246.
- ↑ Michell, RH (1981). "The stimulation of inositol lipid metabolism that accompanies calcium mobilization in stimulated cells: defined characteristics and unanswered questions". Philosophical Transactions of the Royal Society B. 296 (1080): 123—137. Bibcode:1981RSPTB.296..123M. doi:10.1098/rstb.1981.0177. PMID 6121338.
- ↑ Nishizuka, Y (1986). "Studies and perspectives of protein kinase C". Science. 233 (4761): 305—312. Bibcode:1986Sci...233..305N. doi:10.1126/science.3014651. PMID 3014651.
- ↑ Rhee, SG (1989). "Studies of inositol phospholipid-specific phospholipase C". Science. 244 (4904): 546—550. Bibcode:1989Sci...244..546R. doi:10.1126/science.2541501. PMID 2541501. Архивировано 1 июня 2023. Дата обращения: 13 декабря 2023.
- ↑ 1 2 Biaggioni I., Robertson D. (2011). Chapter 9. Adrenoceptor Agonists & Sympathomimetic Drugs. In: B.G. Katzung, S.B. Masters, A.J. Trevor (Eds), Basic & Clinical Pharmacology, 11e. Retrieved October 11, 2011 from AccessMedicine | Case Study. Дата обращения: 30 ноября 2011. Архивировано из оригинала 30 сентября 2011 года..
- ↑ 1 2 Barrett KE, Barman SM, Boitano S, Brooks H. Chapter 2. Overview of Cellular Physiology in Medical Physiology. In: K.E. Barrett, S.M. Barman, S. Boitano, H. Brooks (Eds), Ganong’s Review of Medical Physiology, 23e. AccessMedicine | Objectives. Дата обращения: 30 ноября 2011. Архивировано из оригинала 14 июня 2012 года..
- ↑ Somlyo, AP (1994). "Signal transduction and regulation in smooth muscle". Nature. 372 (6503): 231—6. Bibcode:1994Natur.372..231S. doi:10.1038/372231a0. PMID 7969467.
- ↑ Worley, PF (1989). "Inositol 1,4,5-trisphosphate receptor binding: autoradiographic localization in rat brain". J. Neurosci. 9 (1): 339—46. doi:10.1523/JNEUROSCI.09-01-00339.1989. PMID 2536419.
- ↑ Sarkisov, DV (2008). "Order-dependent coincidence detection in cerebellar Purkinje neurons at the inositol trisphosphate receptor". J. Neurosci. 28 (1): 133—42. doi:10.1523/JNEUROSCI.1729-07.2008. PMID 18171931.
- ↑ Bezprozvanny, I. (2004). "Deranged neuronal calcium signaling and Huntington disease". Biochemical and Biophysical Research Communications. 322 (4): 1310—1317. doi:10.1016/j.bbrc.2004.08.035. PMID 15336977.
- ↑ Alzheimer’s Society of Canada. (2009). Alzheimer’s Disease:What is Alzheimer’s? Retrieved from: http://www.alzheimer.ca/english/disease/whatisit-intro.htm Архивировано {{{2}}}.
- ↑ Stutzmann, G. E. (2005). "Calcium Dysregulation, IP3 Signaling, and Alzheimer's Disease". Neuroscientist. 11 (2): 110—115. doi:10.1177/1073858404270899. PMID 15746379.
- ↑ Berridge, M. J. (2016). "The Inositol Trisphosphate/Calcium Signaling Pathway in Health and Disease". Physiological Reviews. 96 (4): 1261—1296. doi:10.1152/physrev.00006.2016. PMID 27512009.