Кальпонин


Кальпони́н (англ. calponin) — белок из семейства актин-связывающих белков[англ.], взаимодействующий с актином в глобулярной и фибриллярной форме. Типичен для гладкомышечных клеток, в меньшем количестве кальпонины экспрессируются в других типах клеток.
Этимология
Кальпонин получил своё название в связи с наличием домена кальпониновой гомологии[англ.] (англ. calponin homology domain), который также характерен для многих других актин-связывающих белков. При этом необычной особенностью кальпонина стало то, что домен кальпониновой гомологии в нём не связывает актин[1], но вместо этого взаимодействует с фосфолипидами[2][3]
Разновидности
Кальпонины являются продуктом 3 генов и подразделяются на так называемые α-кальпонин (h1 или основная, 292 а.к., 34кДа), β-кальпонин (252 а.к.) — являющийся продуктом одного и того же гена, что и α-кальпонин, но имеющий делецию аминокислот в отрезке 216—256. Две другие изоформы - нейтральный кальпонин (h2 кальпонин) и кислый кальпонин, экспрессируются в гладких мышцах и в немышечных клетах. Другими представителями кальпонинов являются SM22 и трансгелин. Актин-связывающий сайт, как считается (Mezgueldi et al., 1992), расположен в участке между CH-доменом и кальпонин-подобными повторами (CLIK - calponin-like repeat). Кальпонин термостабилен. Молекулярная масса гладкомышечной изоформы - 34 кДа. В других тканях представлены более кислые изоформы кальпонина, не связывающие кальций, но связывающие кальмодулин. Связывание Ca2+/кальмодулина ингибирует связывание с актином. К аналогичному эффекту приводит Фосфорилирование кальпонина серин-треониновыми киназами. Кальпонин имеет также тропомиозин-связывающий сайт.
Свойства
Кальпонин является актин-, кальмодулин-, и тропомиозин-связывающим белком, который был обнаружен в гладкомышечных тканях нескольких позвоночных и в некоторых немышечных клетках. В ходе препарирования тонких нитей куриного желудка было показано, что они содержат актин, тропомиозин, кальдесмон и кальпонин в молярном соотношении 7:0.9:0.6:0.7. Подобно кальдесмону кальпонин ингибирует активность актомиозиновой АТФазы в растворе, скольжение актина относительно фосфорилированного миозина in vitro, и это ингибирование запускается Ca2+/кальмодулином. Ингибирование ликвидируется фосфорилированием белка in vitro. Эти свойства предполагают, что кальпонин может быть дополнительным регулятором актин-миозиновых взаимодействий.
Переваривание кальпонина
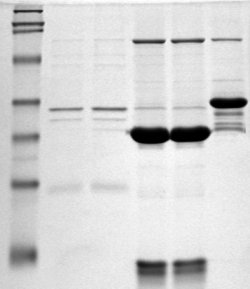
Полное или частичное химотриптическое переваривание кальпонина осуществляется при 25°С, используя весовое отношение протеазы к субстрату 1:100 или 1:1000, соответственно, в 10 мМ имидазол-HCl, 30 мМ NaCl, 2 mM MgCl2, 4 мМ NaN3, pH 7.0, в отсутствие или в присутствии F-актина (4 мг/мл) (молярное соотношение актин/кальпонин = 3:1). При нужных интервалах времени (0-60 мин), вполне определённые количества белков смешиваются с равным объёмом кипящего буфера образцов Laemmli и подготовляются для гель электрофореза. Для реакций сшивания и экспериментов связывания, переваривание прекращается с 2.5 мМ фенилметилсульфонил флуоридом (PMSF). Общий гидролиз ковалентного комплекса кальпонин-актин (11 мг/мл) с CNBr (67 мг/мл, добавленный в двух равных количествах) проводится в течение 24 часов в 70 % муравьиная кислота-содержащем 14 мМ β-меркаптоэтаноле.
Реакции сшивания
Кальпонин (0.7-1.3 мг/мл) в 10 мМ имидазол-HCl, 30 мМ NaCl, 2мМ MgCl2, 4мМ NaN3, pH 7.0, смешивается с нативным или субтилизин-расщепленным F-актином (2.4-4.5 мг/мл)(кальпонин/актин молярное соотношение = 1:3) в присутствии 2 мМ EDC и 5 мМ NHS. Реакция проводилась при 25 °С в течение 0-30 мин и контролируется SDS-гель электрофорезом. Кальдесмон (1.4 мг/мл) сшивается с F-актином при одинаковых условиях с использованием белкового молярного отношения = 1:6. Ковалентная связь между F-актином и 22 кДа NH2-терминальным химотриптическим фрагментом кальпонина получается следующим образом: 60 минутное химотриптическое переваривание кальпонина, при весовом соотношении фермента к субстрату 1:1000, дополняется с F-актином при молярном соотношении 1:3, после центрифугирования при 180,000 х g в течение 1 часа при 4оС, осадок ресуспензируется в имидазол-HCl, pH 7.0, буфере. Для препаративного разделения 76 кДа F-актин-кальпонинового аддукта от несшитого кальпонина, раствор сшитого F-актин-кальпонина (110 мг белка), полученного после 45 мин реакции, доводится в 3 мМ β-меркаптоэтаноле, диализированный в течение ночи при 4 °C относительно деполимеризующего буфера (50 мМ Трис-HCl, 0.6 м КI, 5 мМ тиосульфат натрия, 0.5 мМ АТФ, 0.5 мМ CaCl2, и 0.5 мМ β-меркаптоэтанол, рН 7.5), а затем гель-фильтруется через колонку Sephacryl-S-300 (1.6 х 120 см), уравновешивая при 4 °C в том же буфере. Начальные белковые фракции эльюируются содержащий 76 кДа аддуктом лишенный свободного кальпонина, как и анализируется гель-электрофорезом. Объединенные фракции подвергаются диализу дистиллированной водой, лиофилизируют, и подвергают CNBr расщеплению.
Примечания
- ↑ Gimona M., Mital R. The single CH domain of calponin is neither sufficient nor necessary for F-actin binding // Journal of Cell Science. — 1998. — Vol. 111. — P. 1813—1821. — doi:10.1242/jcs.111.13.1813.
- ↑ Bogatcheva N. V., Gusev N. B. Interaction of smooth muscle calponin with phospholipids // FEBS letters. — 1995. — Vol. 371. — С. 123—126. — doi:10.1016/0014-5793(95)00868-a.
- ↑ Fujii T., Yamana K., Ogoma Y., Kondo Y. Interaction of calponin with phospholipids // Journal of Biochemistry. — 1995. — Vol. 117. — P. 999—1003. — doi:10.1093/oxfordjournals.jbchem.a124833.
Ссылки
- The Calponin Family (англ.). Maciver Lab (4 февраля 2003). Дата обращения: 17 января 2024.
- Mapping of the Functional Domains in the Amino-terminal Region of Calponin (англ.). Дата обращения: 17 января 2024.