
Митохо́ндрия — двумембранная сферическая или эллипсоидная органелла диаметром обычно около 1 микрометра. Характерна для большинства эукариотических клеток, как автотрофов, так и гетеротрофов. Энергетическая станция клетки; основная функция — окисление органических соединений и использование освобождающейся при их распаде энергии для генерации электрического потенциала, синтеза АТФ и термогенеза. Эти три процесса осуществляются за счёт движения электронов по электронно-транспортной цепи белков внутренней мембраны. Количество митохондрий в клетках различных организмов существенно отличается: так, одноклеточные зелёные водоросли и трипаносомы имеют лишь одну гигантскую митохондрию, тогда как ооцит и амёба Chaos chaos содержат 300 000 и 500 000 митохондрий соответственно; у кишечных анаэробных энтамёб и некоторых других паразитических простейших митохондрии отсутствуют. В специализированных клетках органов животных содержатся сотни и даже тысячи митохондрий.

Органе́ллы, раннее называвшиеся также органо́идами, — постоянные компоненты клетки. Располагаются во внутренней части клетки — цитоплазме, в которой, наряду с органеллами, могут находиться различные включения.

Цитохром — небольшой гем-содержащий белок, относится к классу цитохромов, содержит в структуре гем типа c. Выполняет в клетке две функции. С одной стороны, он является одноэлектронным переносчиком, свободно связанным с внутренней мембраной митохондрий, и необходимым компонентом дыхательной цепи. Он способен окисляться и восстанавливаться, но не связывает при этом кислород. С другой стороны, при определённых условиях он может отсоединяться от мембраны, переходить в раствор в межмембранном пространстве и активировать апоптоз. Такая двойственность связана со специфичными свойствами молекулы цитохрома c.

Клеточное, или тканевое дыхание — совокупность биохимических реакций, протекающих в клетках живых организмов, в ходе которых происходит окисление углеводов, липидов и аминокислот до углекислого газа и воды, а также образование энергии. Высвобожденная энергия запасается в химических связях макроэргических соединений и может быть использована по мере необходимости. Входит в группу процессов катаболизма. О физиологических процессах транспортировки к клеткам многоклеточных организмов кислорода и удалению от них углекислого газа см. статью Дыхание.
Дыхательная цепь переноса электронов, также электрон-транспортная цепь (сокр. ЭТЦ, англ. ETC, Electron transport chain) — система трансмембранных белков и переносчиков электронов, необходимых для поддержания энергетического баланса. ЭТЦ поддерживает баланс за счёт переноса электронов и протонов из НАД∙Н и ФАДН2 в акцептор электронов. В случае аэробного дыхания акцептором может быть молекулярный кислород (О2). В случае анаэробного дыхания акцептором могут быть NO3−, NO2−, Fe3+, фумарат, диметилсульфоксид, сера, SO42−, CO2 и т. д. ЭТЦ у прокариот локализована в ЦПМ, у эукариот — на внутренней мембране митохондрий. Переносчики электронов расположены в порядке уменьшения сродства к электрону, то есть по своему окислительно-восстановительному потенциалу, где у акцептора самое сильное сродство к электрону. Поэтому транспорт электрона на всём протяжении цепи протекает самопроизвольно с выделением энергии. Выделение энергии в межмембранное пространство при переносе электронов происходит ступенчато, в виде протона (H+). Протоны из межмембранного пространства попадают в протонную помпу, где наводят протонный потенциал. Протонный потенциал преобразуется АТФ-синтазой в энергию химических связей АТФ. Сопряжённая работа ЭТЦ и АТФ-синтазы носит название окислительного фосфорилирования.
Хемиосмос — биохимический механизм, с помощью которого осуществляется превращение энергии цепи переноса электронов в энергию АТФ. Включает изменение электрохимического потенциала клеточной мембраны.
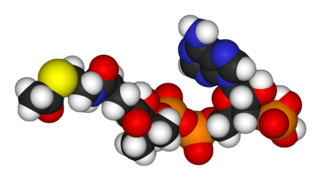
Ацетилкофермент А, ацетил-коэнзим А, сокращённо ацетил-КоА — важное для обмена веществ соединение, используемое во многих биохимических реакциях. Его главная функция — доставлять атомы углерода с ацетил-группой в цикл трикарбоновых кислот, чтобы те были окислены с выделением энергии. По своей химической структуре ацетил-КоА — тиоэфир между коферментом А (тиолом) и уксусной кислотой. Ацетил-КоА образуется во время второго шага кислородного клеточного дыхания, декарбоксилирования пирувата, который происходит в матриксе митохондрии. Ацетил-КоА затем поступает в цикл трикарбоновых кислот.
Липидный обмен, или метаболизм липидов — сложный биохимический и физиологический процесс, происходящий в некоторых клетках живых организмов.

Железосерные кластеры — элементоорганические соединения, группа белковых кофакторов, обладающих окислительно-восстановительным (Red/Ox) потенциалом в районе от −500 мВ до +300 мВ. Red/Ox-потенциал зависит от структуры и конформации белка, что делает эти кофакторы важнейшими участниками окислительно-восстановительных реакций в клетке. Железосерные кластеры способны принимать или отдавать электроны. Белки, содержащие железосерные кластеры, являются эволюционно древними и распространены во всех царствах, включая животных, растения, грибы, бактерии и археи. Мутации по генам метаболизма Fe—S-кластеров являются причиной многих тяжёлых заболеваний или летальны.
Митоптоз — это запрограммированная смерть митохондрий, включающая последовательность биохимических реакций, приводящих к удалению повреждённых митохондрий, или в случае более серьёзных изменений — к интоксикации клетки и её последующему апоптозу. Последовательность событий «сигнал к самоликвидации — митоптоз — апоптоз» отражает основные положения теории самоубийства клеток, которая подразумевает неспособность обеспечить скорость восстановления клеток, соразмерной их гибели.
Бе́та-окисле́ние (β-окисление), также цикл Кноопа — Линена, — метаболический процесс деградации жирных кислот. Своё название процесс получил по 2-му углеродному атому (С-3 или β-положение) от карбоксильной группы (-СООН) жирной кислоты, который подвергается окислению и последовательному отделению от молекулы. Продуктами каждого цикла β-окисления являются ФАДH2, НАДH и ацетил-КоА. Реакции β-окисления и последующего окисления ацетил-КоА в цикле Кребса служат одним из основных источников энергии для синтеза АТФ по механизму окислительного фосфорилирования.

Внутренняя мембрана митохондрий — митохондриальная мембрана, разделяющая митохондриальный матрикс и межмембранное пространство.

Карнитин-О-пальмитоилтрансфераза, также карнитинацилтрансфераза или карнитинпальмитоилтрасфераза, сокр. КПТ — фермент, из семейства ацилтрансфераз, катализирующий перенос ацильной группы с молекулы ацил-КоА на молекулу карнитина и обратно. Относится к интегральным белкам, располагается на внешней стороне внешней мембраны и внутренней стороне внутренней мембраны митохондрий. Является неотъемлемой частью карнитиновой транспортной системы. Имеет несколько различных форм. Участвует в деградации жирных кислот.

Карнитин-пальмитоилтрансфераза I, также карнитинацилтрансфераза I, карнитинацил-СоА-трансфераза I или пальмитоил-CoA-трансфераза I — митохондриальный фермент, одна из форм карнитин-О-пальмитоилтрансферазы, относится к семейству ацилтрансфераз. Катализирует перенос ацильной группы (—COR) от молекулы ацил-CoA жирных кислот с длинной углеводородной цепью на молекулу карнитина, с образованием ацилкарнитина и свободной молекулы кофермента А. Часто продуктом реакции является пальмитоилкарнитин, однако, в качестве субстрата могут выступать и другие остатки жирных кислот. Один из нескольких ферментов карнитиновой транспортной системы. Ген, кодирующий данный фермент локализуется в 11-й хромосоме — CPT1A Архивная копия от 10 сентября 2016 на Wayback Machine.

Карнитин-пальмитоилтрансфераза II, также карнитинацилтрансфераза II, карнитинацил-СоА-трансфераза II или пальмитоил-CoA-трансфераза II — митохондриальный фермент, одна из форм карнитин-О-пальмитоилтрансферазы, относится к семейству ацилтрансфераз. Катализирует перенос ацильной группы (—COR) жирной кислоты с длинной цепью, в частности пальмитоила, от ацилкарнитина на молекулу кофермента А, то есть обратную по отношению к реакции, катализируемой карнитин-пальмитоилтрансферазой I, по уравнению:

Карнитин-О-октаноилтрансфераза, также карнитин-октаноилтрансфераза — митохондриальный фермент, из семейства ацилтрансфераз, который катализирует реакцию переноса ацильной группы (-COR), в частности октаноила, жирных кислот со средней длиной цепи (С6-С8) на молекулу субстрата — карнитин, по уравнению:
Карнитин-ацилкарнитин транслоказа также карнитин-ацилкарнитиновый транспортёр — митохондриальный белок-переносчик, осуществляющий пассивный транспорт ацилированного карнитина (карнитин-COR) внутрь матрикса и карнитина из него, путём антипорта. Локализован на внутренней мембране митохондрий. Ген, кодирующий белок расположен на 3-й хромосоме — SLC25A20 Архивная копия от 17 сентября 2016 на Wayback Machine. Данный белок относится к трансмембранным белкам.
Митохондриа́льный ма́трикс или просто ма́трикс — ограниченное внутренней мембраной пространство, расположенное внутри митохондрий. Слово «матрикс» происходит из того, что эта среда является намного более вязкой по сравнению с более водянистой цитоплазмой. В состав матрикса входит множество веществ, включая ферменты, митохондриальную ДНК (кольцевая), рибосомы, малые органические молекулы, нуклеотидные коферменты и неорганические ионы. Ферменты матрикса содействуют реакциям биохимических процессов, в ходе которых синтезируется АТФ, таких как цикл трикарбоновых кислот, окислительное фосфорилирование, окисление пирувата и бета-окисление жирных кислот.

Гликосо́ма — органелла, окружённая мембраной и содержащая ферменты гликолиза. Термин был введён Скотом и Стиллом в 1968 году, когда они показали, что гликоген, содержащийся в клетке, есть не статичная, а динамичная молекула. Гликосома имеется у нескольких видов протистов, а именно у ряда представителей класса кинетопластид (Kinetoplastea), среди которых есть возбудители таких болезней человека, как сонная болезнь, болезнь Шагаса и лейшманиоз. Органелла окружена одной мембраной и содержит плотный белковый матрикс. Считается, что она произошла от пероксисомы. Эта точка зрения была подтверждена работами по генетике лейшманий (Leishmania). В настоящее время разрабатываются лекарства, действующие на функционирование гликосом.
Транслоказы — отдельный класс ферментов, катализирующих перенос ионов или молекул через мембраны или их разделение в мембранах. Этот класс ферментов сформировался из ранее принадлежащих другим классам ферментов.
