Керберозавр
| † Керберозавр | |
|---|---|
 Реконструкция внешнего облика | |
| Научная классификация | |
Домен: Эукариоты Царство: Животные Подцарство: Эуметазои Без ранга: Двусторонне-симметричные Без ранга: Вторичноротые Тип: Хордовые Подтип: Позвоночные Инфратип: Челюстноротые Надкласс: Четвероногие Клада: Амниоты Клада: Завропсиды Клада: Архозавры Клада: Авеметатарзалии Клада: Динозавроморфы Клада: Динозавры Отряд: † Птицетазовые Подотряд: † Цераподы Инфраотряд: † Орнитоподы Парвотряд: † Игуанодонты Надсемейство: † Гадрозавроиды Семейство: † Гадрозавриды Подсемейство: † Гадрозаврины Род: † Керберозавр | |
| Международное научное название | |
| Kerberosaurus Bolotsky & Godefroit, 2004 | |
| Единственный вид | |
Bolotsky & Godefroit, 2004 | |
Керберозавр[1] (лат. Kerberosaurus) — род растительноядных птицетазовых динозавров семейства гадрозаврид из позднемеловой эпохи Азии. Типовой и единственный вид Kerberosaurus manakini назван и описан российским палеонтологом Юрием Болотским и бельгийским палеонтологом Паскалем Гофруа в 2004 году. Родовое имя является отсылкой к древнегреческой мифологии, в которой трёхглавый пёс Кербер (Цербер) охранял выход из царства мёртвых Аида. Видовое имя названо в честь русского полковника Михаила Манакина, впервые обнаружившего в 1902 году кости динозавров на берегу реки Амур[2].
История исследования
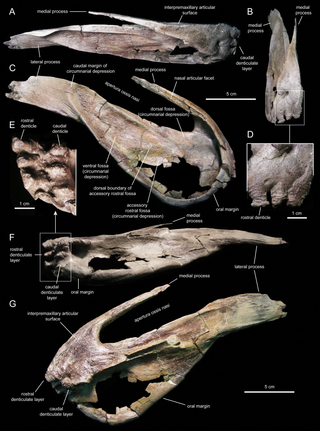
В 1984 году палеонтолог Амурского комплексного НИИ Юрий Болотский обнаружил на берегу реки Амур под Благовещенском, Амурская область, Россия, очень большой костеносный слой, датированный поздним маастрихтом, 70—66 млн лет назад. На 2004 год был раскопан участок площадью около 100 квадратных метров и извлечено несколько сотен костей динозавров. Большинство из них принадлежит ламбеозаврину Amurosaurus riabinini. Однако попадаются и скудные останки черепах, крокодилов, тероподов, нодозавридов и гадрозавриновых динозавров. Останки гадрозаврина скудны и разрозненны, но предположительно принадлежат одному виду. Голотип AENM 1/319 состоит из каудальной части черепа плюс имеется ещё несколько отдельных костей черепа[2].
Описание
Керберозавр диагностируется по следующим признакам: базисфеноидный отросток переднеушной кости сильно углубляется, образуя карман; жёлоб глазничной ветви тройничного нерва сильно расширен на латеральной стороне латеросфеноида; заушное отверстие рострально не ограничено выступающим хребтом; лобная кость сильно сужается в медио-латеральном направлении; ростральный край теменной кости сжат; циркумнариальное вдавление дорсокаудально ограничено большим, широким, уплощённым гребнем на латеральной стороне носовой кости вокруг внешних ноздрей; сильно выступающий крючковидный нёбный отросток челюстной кости[2].
Филогения
Целью филогенетического анализа было установление взаимоотношений керберозвра внутри гадрозавридов. Использовался 21 краниальный признак для 10 таксонов. Аутапоморфии керберозавра были исключены из анализа. В качестве внешней группы был выбран негадрозаврид Bactrosaurus johnsoni. По результатам анализа был выделен ряд синапоморфий гадрозавридов, наблюдающихся у керберозавра: ростральный отросток скуловой кости расширен дорсовентрально; предглазничное отверстие целиком окружено челюстной костью; сильно развит эктоптеригоид; на дистальном конце квадратной кости выделяется большой полусферический латеральный мыщелок; околоквадратное отверстие отсутствует. Эти синапоморфии не наблюдаются у бактрозавра. У керберозавра отсутствуют ламбеозавриновые синапоморфии, но присутствуют гадрозавриновые: циркумнариальное вдавление распространяется на носовую кость; внешние ноздри расширены; в латеральной проекции челюстная кость почти симметрична. Клада зауролофа внутри гадрозаврин характеризуется наличием узкого гребня на голове и каудальным расширением циркумнариального вдавления выше или сзади к ростральному краю глазницы. Анализ показал, что керберозавр является сестринским таксоном к кладе зауролофа на основе того, что лобная кость не является частью глазницы (конвергентно схожий признак с ламбеозавринами). Клада эдмонтозавра (Edmontosaurus и Anatotitan), характеризующаяся, например, массивной скуловой костью, образует с керберозавром и кладой зауролофа монофилетическую группу на основе синапоморфий: ростральный отросток скуловой кости асимметрично изогнут; каудальная граница циркумнариального вдавления отмечена хорошо развитым гребнем. Клады зауролофа и эдмонтозавра сходны также удлинённой нижнечелюстной диастемой и соответствующим расширением предчелюстной кости, но эти признаки ненаблюдаемы у керберозавра вследствие отсутствия соответствующего краниального материала. Клада брахилофозавра (Brachylophosaurus и Maiasaura) рассматривается как базальная среди гадрозаврин по отсутствию расширения верхнезатылочного/бокового затылочного выступа над большим затылочным отверстием, наблюдаемого у других форм. Брахилофозавр и майазавра сходны треугольным ростральным отростком скуловой кости; вентральным выступом скуловой кости; широком плотным гребнем[2].
Кладограмма на основе анализа Bolotsky & Godefroit, 2004:
| Hadrosauroidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Prieto-Marquez, 2010, в глобальном филогенетическом анализе пересмотрел родословную гадрозавров, восстановив подсемейство завролофин (Saurolophinae), и поместив в него керберозавра[3].
Кладограмма на основе анализа Prieto-Marquez, 2010:
| Saurolophinae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиогеография
Предполагается, что керберозавр отделился от группы клад зауролофа и эдмонтозавра не позднее конца кампана. Почти в течение всего позднего мела Северная Америка оказалась разделена внутренним морем на два материка: западный — Ларамидию и восточный — Аппалачию. В течение апта — альба, вероятно, открылся Берингов перешеек между Ларамидией и азиатским материком и сохранялся до конца мела. В течение кампана — маастрихта много групп позвоночных мигрировали через этот перешеек из Азии в Северную Америку. Это касается базальных неоцератопсов, цератопсов, анкилозавридов, тираннозавридов и троодонтидов. Изучение фауны Амурского региона указывает на то, что, возможно, имели место миграции нескольких линий гадрозавров из Западной Америки в Азию. Общий предок керберозавра и клады зауролофа, скорее всего, жил в Ларамидии около или в начале позднего кампана. Предки керберозавра мигрировали в восточную Азию перед поздним маастрихтом. Ламбеозаврин Charonosaurus jiayinensis из «среднего»-позднего маастрихта Азии (китайский берег реки Амур) является сестринским таксоном паразавролофу из позднего кампана Ларамидии. Эта ситуация также предполагает миграцию с востока на запад перед поздним маастрихтом. Завролоф известен из раннего маастрихта западной Канады и Монголии. Его сестринский таксон прозавролоф известен только из позднего кампана западной части Северной Америки. Таким образом, возможно, завролоф также мигрировал из Северной Америки в Азию в конце кампана или в начале маастрихта. Следовательно, можно сделать вывод, что несмотря на то, что Берингов перешеек располагался в полярном регионе, существовал обширный обмен фауной между Азией и Северной Америкой, так что оба эти региона с биогеографической точки зрения в течение кампана — маастрихта были фактически объединены в один.
Фауна маастрихта Амурского региона сильно отличается от фауны формации Лэнс из Ларамидии, датированной тем же временем. В последней преобладают либо цератопсы, либо титанозавры. По всей видимости, в позднем маастрихте члены клады эдмонтозавра и ламбеозаврины исчезли из Северной Америки, либо представлены крайне скудным и неопределённым материалом. С другой стороны, ламбеозаврины преобладают в маастрихте Амурского региона, в то время как цератопсы и титанозавры, по-видимому, отсутствуют. Причиной появления в течение позднего маастрихта большого многообразия популяций динозавров может быть географическая изоляция между восточной Азией и Северной Америкой в этот период, либо огромная разница в климате или палеоэкологических условиях между двумя этими регионами. Авторы предполагают, что местонахождение костей под Благовещенском датируется «средним»-поздним маастрихтом. Следовательно, не может быть исключено, что наблюдаемые различия фаун Амурского региона и формации Лэнс Северной Америки объясняются скорее неопределённостью в датировке, нежели пространственными разграничениями[2].
Примечания
- ↑ Алифанов В. Р., Кириллова И. В. Обзор дочетвертичных позвоночных острова Сахалин // Ученые записки Сахалинского государственного университета : сборник научных статей : [арх. 2 сентября 2017] / Гл. ред. и сост. Т. К. Злобин. — Южно-Сахалинск : СахГУ, 2008. — Вып. VII. — С. 40.
- ↑ 1 2 3 4 5 Bolotsky Y. L., Godefroit P. (2004). «A new hadrosaurine dinosaur from the Late Cretaceous of Far Eastern Russia». Journal of Vertebrate Paleontology 24 (2): 351—365.
- ↑ Prieto-Marquez, A. (2010). «Global phylogeny of Hadrosauridae (Dinosauria: Ornithopoda) using parsimony and Bayesian methods». Zoological Journal of the Linnean Society 159 (2): 435—502.
| Базальные | 
 | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hadrosauro- morpha |
| ||||||||||||||||||||||||||||||