Кренархеоты
| Кренархеоты | |
|---|---|
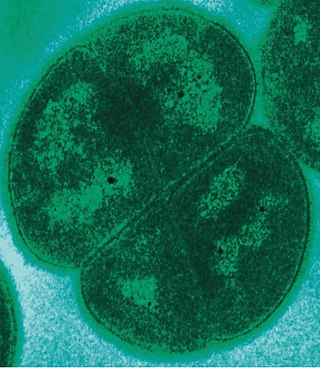
![Sulfolobus, инфицированный вирусом Sulfolobus tengchongensis spindle-shaped virus 1 (STSV-1)[1][2][3][4]. В левой и нижней частях фотографии видны две вирусные частицы веретеновидной формы, отпочковывающиеся от клетки археи. Масштаб = 1 мкм](https://upload.wikimedia.org/wikipedia/commons/thumb/4/46/RT8-4.jpg/275px-RT8-4.jpg) Sulfolobus, инфицированный вирусом Sulfolobus tengchongensis spindle-shaped virus 1 (STSV-1)[1][2][3][4]. В левой и нижней частях фотографии видны две вирусные частицы веретеновидной формы, отпочковывающиеся от клетки археи. Масштаб = 1 мкм | |
| Научная классификация | |
Домен: Археи Тип: Кренархеоты | |
| Международное научное название | |
| Crenarchaeota Garrity and Holt 2002 | |
| Синонимы | |
Кренархео́ты[6] (лат. Crenarchaeota, от др.-греч. κρήνη — ручей, ἀρχαῖος — древний) — тип в составе домена архей, включающий единственный класс Thermoprotei Reysenbach 2002[7][8]. Тип выделен на основании анализа последовательностей 16S рРНК.
Описание
Кренархеоты — морфологически разнородная группа. Клетки кокковидной, палочковидной, дискообразной, нитевидной форм. Размеры клеток варьируют от кокков диаметром меньше микрометра (мкм) до филаментов длиной более 100 мкм[9].
Клеточный цикл у исследованных кренархеот устроен одинаково: период до удвоения ДНК (пререпликативный) короткий, а после удвоения (пострепликативный) — длинный, на него приходится от 64 до 77 % всего клеточного цикла[10].
Физиолого-биохимические свойства группы разнообразны: среди кренархеот есть ацидофилы и нейтрофилы, строгие и факультативные анаэробы и строгие аэробы, хемолитоавтотрофы и хемоорганотрофы. Многие кренархеоты используют серу в метаболизме. Наиболее общий энергетический процесс у кренархеот выглядит как H2 + S0 → H2S[6].
Первоначально кренархеот отделили от остальных архей на основании последовательностей рРНК. В дальнейшем было показано, что кренархеоты также отличаются от других архей отсутствием гистонов, однако впоследствии у некоторых кренархеот они были выявлены[11].
Среда обитания
Впервые представители кренархеот были обнаружены в проточной горячей воде. Сегодня их находят в термальных источниках, вулканических котлах и почвах вулканических полей на суше и горячих ключах, «чёрных курильщиках», мини-вулканах и илах в геотермальных участках под водой. Таким образом, предполагали, что в состав кренархеот входят только термофильные и гипертермофильные организмы[12].
Однако в 1996 году был обнаружен психрофильный вид Crenarchaeum symbiosum, который живёт при 10 °C в ассоциации с морскими губками. Также в составе этого типа архей обнаружены фантомные формы, живущие при температуре до −2 °C[12].
Сейчас кренархеоты обнаруживают в полевых и лесных почвах, палеопочвах, озёрных осадках, морском детрите и активном иле анаэробных очистных сооружений[12]. Анализ рРНК из разных сред показал, что кренархеоты — самые многочисленные археи в морской среде[13].
Кренархеоты распространены по всему миру. В частности, они составляют важную часть планктона в полярных, умеренных и тропических водах. Описаны два симбиотических вида кренархеот: один обитает в холодноводной голотурии, а другой был обнаружен в морской губке[14].
Систематика и классификация
Согласно некоторым данным, мезофильные морские кренархеоты выделяются в самостоятельный тип Thaumarchaeota, однако на молекулярном уровне отличия между кренархеотами и Thaumarchaeota весьма немногочисленны[15]. В настоящее время близкие друг к другу типы Thaumarchaeota, Aigarchaeota, Crenarchaeota и Korarchaeota выделяются в надтип TACK[16].
На июнь 2017 года в класс Thermoprotei включают 5—6 порядков[7][8]:
- Порядок Acidilobales Prokofeva et al. 2009
- Порядок Cenarchaeales[англ.] Cavalier-Smith 2002 — согласно данным NCBI, относится к типу Thaumarchaeota[17]
- Порядок Desulfurococcales Huber and Stetter 2002
- Порядок Fervidicoccales Perevalova et al. 2010
- Порядок Sulfolobales Stetter 1989
- Порядок Thermoproteales Zillig and Stetter 1982 emend. Burggraf et al. 1997
Разделение кренархеот на порядки основано на последовательностях генов 16S рРНК. Самую древнюю эволюционную ветвь кренархеот составляют роды Thermoproteus, Thermofilum и Pyrobaculum[9].
Эоцитная гипотеза

Так называемая эоцитная гипотеза была предложена в 1980-х Джеймсом Лейком (англ. James Lake). Согласно ей, эукариоты произошли от прокариотических эоцитов (то есть кренархеот). Возможным подтверждением этой гипотезы является наличие гомолога субъединицы Rbp-8 РНК-полимеразы у кренархеот, но не у остальных архей[19].
Примечания
- ↑ Krupovic M., Quemin E. R. J., Bamford D. H., Forterre P., Prangishvili D. Unification of the Globally Distributed Spindle-Shaped Viruses of the Archaea // Journal of Virology. — 2013. — 11 декабря (т. 88, № 4). — С. 2354—2358. — ISSN 0022-538X. — doi:10.1128/JVI.02941-13.
- ↑ Hochstein Rebecca, Bollschweiler Daniel, Engelhardt Harald, Lawrence C. Martin, Young Mark. Large Tailed Spindle Viruses of Archaea: a New Way of Doing Viral Business // Journal of Virology. — 2015. — 17 июня (т. 89, № 18). — С. 9146—9149. — ISSN 0022-538X. — doi:10.1128/JVI.00612-15.
- ↑ Robb F., Antranikian G., Grogan D., Driessen A. (Editors). Thermophiles: Biology and Technology at High Temperatures. — CRC Press, 2007. — P. 231—232. — 368 p. — ISBN 978-0849392146.
- ↑ Uneven Distribution of Viruses Suggests Surprising Evolutionary Power : [англ.] // Scientific American.
- ↑ 1 2 Crenarchaeota (англ.) на сайте Национального центра биотехнологической информации (NCBI). (Дата обращения: 23 июля 2017).
- ↑ 1 2 Воробьёва, 2007, с. 317.
- ↑ 1 2 Classification of domains and phyla - Hierarchical classification of prokaryotes (bacteria) : Version 2.0 : [англ.] // LPSN. — 2016. — 2 October.
- ↑ 1 2 Taxonomy Browser : Crenarchaeota : [англ.] // NCBI. (Дата обращения: 23 июля 2017).
- ↑ 1 2 Tolweb: Crenarchaeota. Дата обращения: 10 августа 2017. Архивировано 10 августа 2017 года.
- ↑ Lundgren M., Malandrin L., Eriksson S., Huber H., Bernander R. Cell Cycle Characteristics of Crenarchaeota: Unity among Diversity // Journal of Bacteriology. — 2008. — 23 мая (т. 190, № 15). — С. 5362—5367. — ISSN 0021-9193. — doi:10.1128/JB.00330-08.
- ↑ Cubonová L., Sandman K., Hallam S. J., Delong E. F., Reeve J. N. Histones in crenarchaea. (англ.) // Journal of bacteriology. — 2005. — Vol. 187, no. 15. — P. 5482—5485. — doi:10.1128/JB.187.15.5482-5485.2005. — PMID 16030242.
- ↑ 1 2 3 Пиневич, 2006, с. 90.
- ↑ Madigan M., Martinko J. (editors). Brock Biology of Microorganisms (неопр.). — 11th. — Prentice Hall, 2005. — ISBN 0-13-144329-1.
- ↑ Joanne M. Willey, Linda M. Sherwood, Christopher J. Woolverton. Prescott's Principles of Microbiology. — 1st edition. — McGraw-Hill Higher Education, 2009. — P. 413. — 968 p. — ISBN 978-0-07-337523-6.
- ↑ Issues in Life Sciences—Muscle, Membrane, and General Microbiology: 2012 Edition / Q. Ashton Acton. — ScholarlyEditions, 2012. — P. 100. — 223 p. — ISBN 978-1-4816-4674-1.
- ↑ Zuo G., Xu Z., Hao B. Phylogeny and Taxonomy of Archaea: A Comparison of the Whole-Genome-Based CVTree Approach with 16S rRNA Sequence Analysis. (англ.) // Life (Basel, Switzerland). — 2015. — Vol. 5, no. 1. — P. 949—968. — doi:10.3390/life5010949. — PMID 25789552.
- ↑ Taxonomy Browser : Cenarchaeales : [англ.] // NCBI. (Дата обращения: 25 июля 2017).
- ↑ Cox C. J., Foster P. G., Hirt R. P., Harris S. R., Embley T. M. The archaebacterial origin of eukaryotes. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 2008. — Vol. 105, no. 51. — P. 20356—20361. — doi:10.1073/pnas.0810647105. — PMID 19073919.
- ↑ Kwapisz M., Beckouët F., Thuriaux P. Early evolution of eukaryotic DNA-dependent RNA polymerases. (англ.) // Trends in genetics : TIG. — 2008. — Vol. 24, no. 5. — P. 211—215. — doi:10.1016/j.tig.2008.02.002. — PMID 18384908.
Литература
- Воробьёва Л. И. Археи: учебное пособие для вузов. — М.: ИКЦ «Академкнига», 2007. — 447 с. — ISBN 978-5-94628-277-2.
- Пиневич А. В. Микробиология. Биология прокариотов : Учебник : в 3 т. — СПб. : Издательство С.-Петербургского университета, 2006. — Т. 1. — 352 с. — ISBN 5-288-04057-5.
| Таксономия |
|---|
Классификация архей | |
|---|---|
| Euryarchaeota |
|
| TACK[англ.] |
|
| Asgard[англ.] |
|
| DPANN[англ.] |
|