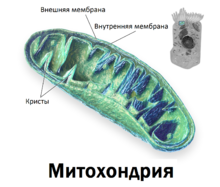
Митохо́ндрия — двумембранная сферическая или эллипсоидная органелла диаметром обычно около 1 микрометра. Характерна для большинства эукариотических клеток, как автотрофов, так и гетеротрофов. Энергетическая станция клетки; основная функция — окисление органических соединений и использование освобождающейся при их распаде энергии для генерации электрического потенциала, синтеза АТФ и термогенеза. Эти три процесса осуществляются за счёт движения электронов по электронно-транспортной цепи белков внутренней мембраны. Количество митохондрий в клетках различных организмов существенно отличается: так, одноклеточные зелёные водоросли и трипаносомы имеют лишь одну гигантскую митохондрию, тогда как ооцит и амёба Chaos chaos содержат 300 000 и 500 000 митохондрий соответственно; у кишечных анаэробных энтамёб и некоторых других паразитических простейших митохондрии отсутствуют. В специализированных клетках органов животных содержатся сотни и даже тысячи митохондрий.

Органе́ллы, раннее называвшиеся также органо́идами, — постоянные компоненты клетки. Располагаются во внутренней части клетки — цитоплазме, в которой, наряду с органеллами, могут находиться различные включения.

Цитохром — небольшой гем-содержащий белок, относится к классу цитохромов, содержит в структуре гем типа c. Выполняет в клетке две функции. С одной стороны, он является одноэлектронным переносчиком, свободно связанным с внутренней мембраной митохондрий, и необходимым компонентом дыхательной цепи. Он способен окисляться и восстанавливаться, но не связывает при этом кислород. С другой стороны, при определённых условиях он может отсоединяться от мембраны, переходить в раствор в межмембранном пространстве и активировать апоптоз. Такая двойственность связана со специфичными свойствами молекулы цитохрома c.
Активный транспорт — перенос вещества через клеточную или внутриклеточную мембрану или через слой клеток, протекающий из области низкой концентрации в область высокой, т. е. с затратой свободной энергии организма. В большинстве случаев, но не всегда, источником энергии для осуществления активного транспорта служит энергия макроэргических связей АТФ.

Аденозинтрифосфатсинта́за (АТФ-синта́за, АТФ-фосфогидролаза, H+-transporting two-sector ATPase) — группа ферментов, относящихся к классу транслоказ и синтезирующих аденозинтрифосфат (АТФ) из аденозиндифосфата (АДФ) и неорганического фосфата. Название по номенклатуре — АТФ-фосфогидролаза, однако с августа 2018 года фермент перенесён из третьего (3.6.3.14) в седьмой класс (7.1.2.2), так как катализируемая ферментом реакция протекает по пути, противоположному гидролизу, и не может быть описана с помощью других типов реакций, характеризующие прочие классы ферментов.

Клеточное, или тканевое дыхание — совокупность биохимических реакций, протекающих в клетках живых организмов, в ходе которых происходит окисление углеводов, липидов и аминокислот до углекислого газа и воды, а также образование энергии. Высвобожденная энергия запасается в химических связях макроэргических соединений и может быть использована по мере необходимости. Входит в группу процессов катаболизма. О физиологических процессах транспортировки к клеткам многоклеточных организмов кислорода и удалению от них углекислого газа см. статью Дыхание.
Дыхательная цепь переноса электронов, также электрон-транспортная цепь (сокр. ЭТЦ, англ. ETC, Electron transport chain) — система трансмембранных белков и переносчиков электронов, необходимых для поддержания энергетического баланса. ЭТЦ поддерживает баланс за счёт переноса электронов и протонов из НАД∙Н и ФАДН2 в акцептор электронов. В случае аэробного дыхания акцептором может быть молекулярный кислород (О2). В случае анаэробного дыхания акцептором могут быть NO3−, NO2−, Fe3+, фумарат, диметилсульфоксид, сера, SO42−, CO2 и т. д. ЭТЦ у прокариот локализована в ЦПМ, у эукариот — на внутренней мембране митохондрий. Переносчики электронов расположены в порядке уменьшения сродства к электрону, то есть по своему окислительно-восстановительному потенциалу, где у акцептора самое сильное сродство к электрону. Поэтому транспорт электрона на всём протяжении цепи протекает самопроизвольно с выделением энергии. Выделение энергии в межмембранное пространство при переносе электронов происходит ступенчато, в виде протона (H+). Протоны из межмембранного пространства попадают в протонную помпу, где наводят протонный потенциал. Протонный потенциал преобразуется АТФ-синтазой в энергию химических связей АТФ. Сопряжённая работа ЭТЦ и АТФ-синтазы носит название окислительного фосфорилирования.

Окисли́тельное фосфорили́рование — метаболический путь, при котором энергия, образовавшаяся при окислении питательных веществ, запасается в митохондриях клеток в виде АТФ. Хотя различные формы жизни на Земле используют разные питательные вещества, АТФ является универсальным соединением, в котором запасается энергия, необходимая для других метаболических процессов. Почти все аэробные организмы осуществляют окислительное фосфорилирование. Вероятно, широкому распространению этого метаболического пути способствовала его высокая энергетическая эффективность по сравнению с анаэробным брожением.
Кардиолипин — фосфолипид, который является важным компонентом внутренней мембраны митохондрий, липидный состав которой включает около 20 % кардиолипина. Кардиолипин во внутренней мембране митохондрий клеток млекопитающих и растительных клеток необходим для функционирования многочисленных ферментов, участвующих в энергетическом обмене. Кардиолипин также встречается в мембранах бактерий.

Цитохром-c-оксида́за (цитохромоксидаза) или цитохром-c-кислород:оксидоредуктаза, также известная как цитохром aa3 и комплекс IV — терминальная оксидаза аэробной дыхательной цепи переноса электронов, которая катализирует перенос электронов с цитохрома с на кислород с образованием воды. Цитохромоксидаза присутствует во внутренней мембране митохондрий всех эукариот, где её принято называть комплекс IV, а также в клеточной мембране многих аэробных бактерий.
Протонный насос в молекулярной биологии — механизм, создающий или использующий протонный градиент. Протонный насос является белковым комплексом, в котором имеются мембранные белки. В процессе работы протонного насоса осуществляется перенос протонов через мембрану клетки, митохондрии или другого внутриклеточного компартмента. Первичными протонными насосами называют комплексы, в которых разность концентраций используется для синтеза АТФ; вторичные протонные насосы расщепляют АТФ для создания протонного градиента.

Тилакоиды — ограниченные мембраной компартменты внутри хлоропластов и цианобактерий. В тилакоидах происходят светозависимые реакции фотосинтеза. Слово «тилакоид» происходит от греческого слова θύλακος, означающего «мешочек». Тилакоиды состоят из мембраны, окружающей просвет тилакоида. Тилакоиды хлоропластов часто имеют структуру, напоминающую стопку дисков. Эти стопки называют гранами. Граны соединены межграновыми или строматическими тилакоидами (ламеллами) в единое функциональное пространство.
Хемиосмос — биохимический механизм, с помощью которого осуществляется превращение энергии цепи переноса электронов в энергию АТФ. Включает изменение электрохимического потенциала клеточной мембраны.

Железосерные кластеры — элементоорганические соединения, группа белковых кофакторов, обладающих окислительно-восстановительным (Red/Ox) потенциалом в районе от −500 мВ до +300 мВ. Red/Ox-потенциал зависит от структуры и конформации белка, что делает эти кофакторы важнейшими участниками окислительно-восстановительных реакций в клетке. Железосерные кластеры способны принимать или отдавать электроны. Белки, содержащие железосерные кластеры, являются эволюционно древними и распространены во всех царствах, включая животных, растения, грибы, бактерии и археи. Мутации по генам метаболизма Fe—S-кластеров являются причиной многих тяжёлых заболеваний или летальны.

Внутренняя мембрана митохондрий — митохондриальная мембрана, разделяющая митохондриальный матрикс и межмембранное пространство.
Митохондриа́льный ма́трикс или просто ма́трикс — ограниченное внутренней мембраной пространство, расположенное внутри митохондрий. Слово «матрикс» происходит из того, что эта среда является намного более вязкой по сравнению с более водянистой цитоплазмой. В состав матрикса входит множество веществ, включая ферменты, митохондриальную ДНК (кольцевая), рибосомы, малые органические молекулы, нуклеотидные коферменты и неорганические ионы. Ферменты матрикса содействуют реакциям биохимических процессов, в ходе которых синтезируется АТФ, таких как цикл трикарбоновых кислот, окислительное фосфорилирование, окисление пирувата и бета-окисление жирных кислот.

Электрохими́ческий градие́нт, или градиéнт электрохимического потенциáла, — совокупность градиента концентрации и мембранного потенциала, которая определяет направление движения ионов через мембрану. Состоит из двух составляющих: химического градиента, или разницы в концентрациях растворённого вещества по обе стороны мембраны, и электрического градиента, или разницы зарядов, расположенных на противоположных сторонах мембраны. Градиент возникает вследствие неодинаковой концентрации ионов на противоположных сторонах водопроницаемой мембраны. Ионы двигаются через мембрану из области, имеющую более высокую концентрацию в область с более низкой концентрацией путём простой диффузии. Также ионы несут электрический заряд, который формирует электрический потенциал на мембране. Если существует неравномерное распределение зарядов по обе стороны мембраны, то разница в электрическом потенциале порождает силу, которая приводит к ионной диффузии, пока заряды по обе стороны не будут сбалансированы.
Митофа́гия — селективное разрушение митохондрий путём аутофагии. Впервые этот термин был введён J. J. Lemasters в 2005 году, хотя ещё с 1962 года было известно о том, что лизосомы клеток печени содержат фрагменты митохондрий, а в 1977 году при изучении метаморфоза тутового шелкопряда было отмечено, что функциональные изменения митохондрий могут вызывать аутофагию. Митофагия необходима для поддержания жизнедеятельности клетки, она способствует кругообороту митохондрий и предотвращает накопление нефункциональных митохондрий, которые могут привести к дегенерации клетки. У дрожжей митофагия опосредуется белком Atg32, а у млекопитающих регуляция митофагии осуществляется белками PINK1 и паркином. Митофагия не приурочена только к повреждённым митохондриям, в ней могут участвовать и неповреждённые митохондрии.
Ионный транспортёр — трансмембранный белок, который перемещает ионы через биологическую мембрану для выполнения множества различных биологических функций, включая клеточную связь, поддержание гомеостаза, производство энергии и т. д. Существуют разные типы транспортёров, включая насосы, унипортёры, антипортёры и симпортёры. Активные транспортёры или ионные насосы — это переносчики, которые преобразуют энергию из различных источников, включая аденозинтрифосфат (АТФ), солнечный свет и другие окислительно-восстановительные реакции, в потенциальную энергию, двигая ион вверх по градиенту его концентрации. Эта потенциальная энергия может затем использоваться вторичными транспортёрами, включая транспортёры ионов и ионные каналы, для управления жизненно важными клеточными процессами, такими как синтез АТФ.
F-АТФаза, также известная как АТФаза F-типа, представляет собой АТФазу/синтазу, обнаруженную в бактериальных клеточных мембранах, во внутренних мембранах митохондрий и в мембранных тилакоидов хлоропластов. F-АТФаза использует протонный градиент для управления синтезом АТФ, используя энергию, высвобождаемую в результате транспортной реакции, для высвобождения вновь образованного АТФ из активного центра F-АТФазы. Вместе с V-АТФазой и A-АТФазой, F-АТФаза принадлежат к суперсемейству родственных роторных ионтранслирующих АТФаз.