Лактозный оперон

Лактозный оперон (англ. lac operon) — полицистронный оперон бактерий, кодирующий гены метаболизма лактозы.
Регуляция экспрессии генов метаболизма лактозы у кишечной палочки (Escherichia coli) была впервые описана в 1961 году учёными Ф. Жакобом и Ж. Моно[1] (получившими в 1965 году Нобелевскую премию совместно с А. Львовым). Бактериальная клетка синтезирует ферменты, принимающие участие в метаболизме лактозы, лишь в том случае, когда лактоза присутствует в окружающей среде и клетка испытывает недостаток глюкозы.
Структура
Лактозный оперон (lac operon) состоит из трёх структурных генов, промотора, оператора и терминатора. Иногда принимается, что в состав оперона входит также ген-регулятор, который кодирует белок-репрессор (хотя он находится в другом участке генома и не имеет общего с лактозным опероном промотора).
Структурные гены лактозного оперона — lacZ, lacY и lacA:
- lacZ кодирует фермент β-галактозидазу, которая расщепляет дисахарид лактозу на глюкозу и галактозу,
- lacY кодирует β-галактозидпермеазу, мембранный транспортный белок, который переносит лактозу внутрь клетки.
- lacA кодирует β-галактозидтрансацетилазу, фермент, переносящий ацетильную группу от ацетил-КoA на бета-галактозиды.
Для катаболизма лактозы необходимы только продукты генов lacZ и lacY; роль продукта гена lacA не ясна. Возможно, что реакция ацетилирования даёт бактериям преимущество при росте в присутствии определённых неметаболизируемых аналогов бета-галактозидов, поскольку эта модификация ведёт к их детоксикации и выведению из клетки.
Регуляция
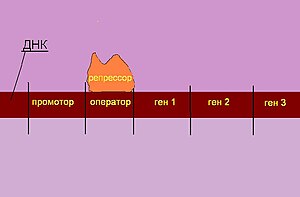



РНК-полимераза начинает транскрипцию с промоторного района, который перекрывается с операторным районом. В отсутствие или при низкой концентрации лактозы в клетке белок-репрессор, который является продуктом моноцистронного оперона LacI, обратимо соединяется с операторным районом и препятствует транскрипции. Таким образом, в отсутствие лактозы в клетке ферменты для метаболизма лактозы не синтезируются.
Даже в случае, когда в плазматической мембране клетки отсутствует фермент β-галактозидпермеаза, лактоза из окружающей среды может попадать в клетку в небольших количествах. В клетке две молекулы лактозы связываются с белком-репрессором, что приводит к изменению его конформации и далее к диссоциации белка-репрессора от операторного участка. Может осуществляться транскрипция генов лактозного оперона. При снижении концентрации лактозы новые порции белка-репрессора взаимодействуют с операторными последовательностями и препятствуют транскрипции. Данный механизм регуляции активности лактозного оперона называют негативной индукцией. Веществом-индуктором служит лактоза; при её связывании с белком-репрессором происходит его диссоциация от операторного участка.
Если в клетке концентрация глюкозы достаточна для поддержания метаболизма, активация лактозного оперона не происходит. Промоторная последовательность лактозного оперона «слабая», поэтому даже при отсутствии белка-репрессора на операторном участке транскрипция практически не инициируется. Когда концентрация глюкозы в клетке снижается, происходит активация фермента аденилатциклазы, которая катализирует превращение АТФ в циклическую форму — цАМФ (циклическую форму АМФ в данном случае также называют «сигналом клеточного голода»). Глюкоза является ингибитором фермента аденилатциклазы и активирует фосфодиэстеразу — фермент, катализирующий превращение молекулы цАМФ в АМФ. цАМФ соединяется с белком, активирующим катаболизм (англ. САР, catabolism activating protein), при этом образуется комплекс, который взаимодействует с промотором лактозного оперона, изменяет его конформацию и приводит к повышению сродства РНК-полимеразы к данному участку. В присутствии лактозы происходит экспрессия генов оперона. Белок CAP оказывает положительный контроль на лактозный оперон.
Итак, ферменты для усвоения лактозы синтезируются в клетке кишечной палочки при двух условиях: 1) наличие лактозы; 2) отсутствие глюкозы. Регуляция работы лактозного оперона в зависимости от концентрации лактозы происходит по принципу отрицательной обратной связи: чем больше лактозы — тем больше ферментов для её катаболизма (положительная прямая связь); чем больше ферментов — тем меньше лактозы, чем меньше лактозы — тем меньше производится ферментов (двойная отрицательная обратная связь).
Биологический смысл
Благодаря описанному механизму регуляции транскрипции генов, входящих в состав лактозного оперона, бактерии оптимизируют энергетические затраты, синтезируя ферменты метаболизма лактозы не постоянно, а лишь тогда, когда клетке это необходимо. Сходный механизм регуляции имеется у большинства прокариот; у эукариот он устроен значительно сложнее.
См. также
Примечания
Ссылки
| Регуляция транскрипции |
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Активация | |||||||||||||
| Инициация | Сайт начала транскрипции | ||||||||||||
| Элонгация |
| ||||||||||||
| Терминация | |||||||||||||











