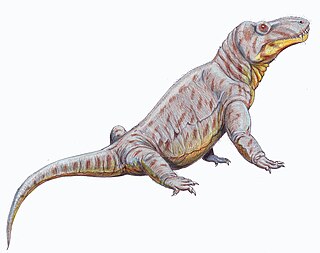
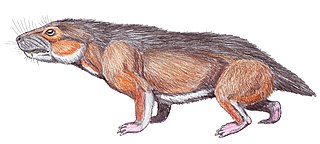
Маммализация териодонтов
Маммализа́ция териодо́нтов — процесс постепенного накопления в ветвях синапсид, терапсид и териодонтов признаков, характерных для млекопитающих. Концепцию с таким названием выдвинул в 1976 году Л. П. Татаринов, обративший внимание на параллельное и независимое появление отдельных признаков млекопитающих в различных группах териодонтов.
Появление первых млекопитающих датируется триасовым периодом, предположительно около 225 млн лет назад[1].
Прогрессивная эволюция млекопитающих была связана прежде всего с приобретением ими таких приспособительных черт, как высокая температура тела, способность к терморегуляции, высокий аэробный уровень метаболизма. Этому способствовали изменения в дыхательной и кровеносной системах: морфологически это выражалось в разделении сердца на четыре камеры и в сохранении одной дуги аорты, обусловивших несмешиваемость артериальной и венозной крови, в появлении вторичного костного нёба, обеспечившего дыхание во время еды, в изменении эффективности кормовой стратегии, обеспечившей ускоренное переваривание пищи. Она оказалась возможной на основе изменения строения челюстей (что привело и к развитию слуховых косточек, чьё наличие усилило возможности слухового анализатора млекопитающих, особенно в отношении звуков высокой частоты), дифференцировки зубов, развития челюстной мускулатуры.
Такие черты млекопитающих, как крупный мозг и живорождение, сложились в эволюции значительно позднее.
Синапсиды как предки млекопитающих

Одна из первых обособившихся групп древних амниот во второй половине палеозоя — синапсиды. Они легли в основу становления подкласса (другие исследователи выделяют их в отдельный класс) звероподобных (лат. Theromorpha), именуемых также терапсидами (лат. Therapsida). В пермском периоде среди них сформировалась группа зверозубые (лат. Theriodontia). Они оказались близкими по уровню организации млекопитающим.
Однако условия жизни, сложившиеся в мезозойскую эру, благоприятствовали эволюционному развитию и увеличению разнообразия завропсидов, и в мезозое доминирующие позиции заняли архозавры. Размеры тела зверозубых уменьшились, сократились их численность и распространение, они были вынуждены довольствоваться второстепенными нишами. Реализация их возможностей осуществилась позднее, что, очевидно, связано с массовым вымиранием архозавров и изменением климата в конце мезозоя.
Что любопытно, появление первых динозавров ознаменовалось массовым, но не полным вымиранием терапсид, высшие формы которых по своей организации были очень близки к однопроходным млекопитающим и, по некоторым предположениям, имели молочные железы и шерсть[2]. Многочисленные копролиты, найденные в верхнепермских отложениях и принадлежащие, вероятно, тероцефалам и базальным архозаврам (протерозухидам), содержат вытянутые полые структуры, которые могут быть остатками волос. Это означает, что терапсиды обладали волосяным покровом уже 252 миллиона лет назад[3].

Наиболее близкими к млекопитающим среди зверозубых были цинодонты (лат. Cynodontia). Наиболее ярко черты скелетных изменений обнаружились среди них у тринаксодона (лат. Thrinaxodon) из раннего триаса.
Ранние млекопитающие
Молочные железы возникли у древних млекопитающих, вероятно, из видоизмененных потовых желез. Возможно, первоначальной функцией млечных желез было не столько выкармливание, сколько «выпаивание» детенышей, то есть снабжение их необходимой влагой и солями.
В последующем становлении млекопитающих палеонтологи подчеркивают изменения в зубной системе. Это привело к вычленению двух групп — морганукодонтид (лат. Morganucodontidae) и Kuehneotheriidae.
К потомкам ранних млекопитающих, найденных в отложениях верхнего триаса, относят своеобразных многобугорчатых, получивших своё название в связи с наличием на коренных зубах многочисленных бугорков. Это была специализированная группа животных с очень сильно развитыми резцами и без клыков. Многобугорчатые представляли специфических растительноядных зверей, и их нельзя считать предками последующих групп млекопитающих. Выдвигались, правда, гипотезы, что их ранние формы дали начало однопроходным, но эти гипотезы устарели.
Вторая группа оказалась более успешной в последующей адаптивной радиации. Их главную линию составили эупантотерии (лат. Eupantotheria); этот термин, впрочем, почти вышел из употребления, поскольку обозначаемая им группа млекопитающих является парафилетической и включает довольно-таки разнообразных мезозойских млекопитающих). Вероятно, они были маленькими зверьками, питавшимися насекомыми, может быть, и другими мелкими животными и яйцами рептилий. Головной мозг их был мал, но больше, чем у немаммальных синапсид. В конце мезозоя в этой группе наметилось разделение на два самостоятельные ствола — низших, сумчатых (Metatheria, Marsupialia), и высших, плацентарных (лат. Eutheria).
Плацентарные млекопитающие, как и сумчатые, возникли в меловом периоде. Как показали исследования, в меловом периоде они уже эволюционировали в весьма различных направлениях.
Во время мел-палеогенового вымирания (около 65 миллионов лет назад) все существовавшие в мезозойской эре экосистемы были полностью разрушены, что впоследствии резко подстегнуло эволюцию млекопитающих, давших в начале палеогена огромное многообразие форм благодаря освобождению большинства экологических ниш[4]. Через 100 тыс. лет после падения метеорита таксономическое разнообразие млекопитающих удвоилось, а максимальная масса тела млекопитающих увеличилась почти до уровней, предшествующих мел-палеогеновому вымиранию. Приблизительно трёхкратное увеличение максимальной массы тела млекопитающих произошло через 300 тыс. лет после мел-палеогенового вымирания, первые крупные млекопитающие появились через 700 тысяч лет после мел-палеогенового вымирания, что совпадает с первым появлением растений семейства бобовых[5].
Кладограмма синапсид
Происхождение млекопитающих прослеживается из следующей цепочки (кладограммы), отражающей филогению синапсид:
Synapsida (Синапсиды) (пеликозавры) |-Caseasauria (эотириды и казеиды) `-Eupelycosauria |-Varanopseidae `-+-Ophiacodontidae `-+-Edaphosauridae `-Sphenacodontia |-Sphenacodontidae `-Therapsida (Терапсиды) |-Biarmosuchia | `-Eotitanosuchus `-Eutherapsida |-Dinocephalia `-Neotherapsida |-Anomodontia `-Theriodontia (Териодонты) |-Gorgonopsia `-Eutheriodontia |-Therocephalia `-Cynodontia (Цинодонты) |- + -Dvinia | `--Procynosuchus `- Epicynodontia |-Thrinaxodon `-Eucynodontia |- + -Cynognathus | `- + -Tritylodontidae | `- Traversodontidae `-Probainognathia |- + - Trithelodontidae | `--Chiniquodontidae `- + - Prozostrodon `- Mammaliaformes `-Mammalia (Млекопитающие)
См. также
- Аделобазилевс
- Borealestes
- Megazostrodon
- Probainognathus
- Pseudotherium
Примечания
- ↑ David M. Hunt, Mark W. Hankins, Shaun P. Collin, N. Justin Marshall. Evolution of Visual and Non-visual Pigments. — Springer, 2014-10-04. — 278 с. — ISBN 9781461443551. Архивировано 22 июня 2018 года.
- ↑ Еськов К. Ю. История Земли и жизни на ней: От хаоса до человека. — М.: ЭНАС, 2008. — 312 с. — ISBN 978-5-93196-711-0. Архивировано 4 мая 2014 года. Архивированная копия. Дата обращения: 21 мая 2014. Архивировано 4 мая 2014 года. — С. 170, 178.
- ↑ А. Г. Сенников, В. К. Голубев, G. Niedzwiedzki, P. Bajdek, K. Owocki, T. Sulej. Древнейшая находка остатков волос в копролитах тетрапод из терминальной перми Владимирской области. — М.: ПИН РАН, 2016. — P. 71-72. Архивная копия от 3 ноября 2023 на Wayback Machine
- ↑ Most comprehensive tree of life shows placental mammal diversity exploded after age of dinosaurs Архивная копия от 26 октября 2019 на Wayback Machine, February 7, 2013
- ↑ Lyson T. R. et al.Exceptional continental record of biotic recovery after the Cretaceous-Paleogene mass extinction Архивная копия от 1 ноября 2019 на Wayback Machine
Литература
- Кэрролл Р. Палеонтология и эволюция позвоночных: В 3-х тт. Т. 2. — М.: Мир, 1993. — 283 с. — ISBN 5-03-001819-0.
- Кэрролл Р. Палеонтология и эволюция позвоночных: В 3-х тт. Т. 3. — М.: Мир, 1993. — 312 с. — ISBN 5-03-001819-0.
Ссылки
| Эволюционные процессы |  | |
|---|---|---|
| Факторы эволюции | ||
| Генетика популяций | ||
| Происхождение жизни | ||
| Исторические концепции | ||
| Современные теории | ||
| Эволюция таксонов | ||