Мезопропитеки
| † Мезопропитеки | |
|---|---|
 | |
| Научная классификация | |
Домен: Эукариоты Царство: Животные Подцарство: Эуметазои Без ранга: Двусторонне-симметричные Без ранга: Вторичноротые Тип: Хордовые Подтип: Позвоночные Инфратип: Челюстноротые Надкласс: Четвероногие Клада: Амниоты Клада: Синапсиды Класс: Млекопитающие Подкласс: Звери Клада: Эутерии Инфракласс: Плацентарные Магнотряд: Бореоэутерии Надотряд: Грандотряд: Эуархонты Миротряд: Приматообразные Отряд: Приматы Подотряд: Мокроносые приматы Инфраотряд: Лемурообразные Семейство: † Палеопропитековые Род: † Мезопропитеки | |
| Международное научное название | |
| Mesopropithecus Standing, 1905 | |
| Виды | |
| |
| Ареал | |
 M. dolichobrachion M. globiceps M. pithecoides[1] | |
Мезопропитеки[2] (лат. Mesopropithecus) — вымерший род ленивцевых лемуров, обитавших на Мадагаскаре в плейстоцене и голоцене. К настоящему времени к роду относят три вида: M. dolichobrachion, M. globiceps и M. pithecoides. По размерам мезопропитеки были мельче остальных ленивцевых лемуров, но крупней современных мадагаскарских лемуров, достигая по весу (в зависимости от вида) 10—14 килограммов. Все мезопропитеки были растительноядными, их диета состояла из листьев, плодов и семян. Мезопропитеки известны по субфоссильным останкам и вымерли недавно: возраст подвергнутых радиоуглеродному анализу останков определяется VI—VII веком новой эры, уже после прихода на Мадагаскар человека, и возможно, что охота и сведение лесов стали причиной вымирания этого рода.
Классификация и происхождение
Род мезопропитеков с его тремя видами, M. dolichobrachion, M. globiceps и M. pithecoides, относится к полностью вымершему семейству палеопропитековых, или ленивцевых лемуров. В это семейство входили также археоиндри (один вид), бабакотии (также один вид) и собственно палеопропитеки (три вида). Семейство палеопропитековых является частью инфраотряда лемурообразных, куда входят все лемуры Мадагаскара. Ближайшими родичами палеопропитековых, как установлено в последние десятилетия XX века, являются современные индриевые.
Род мезопропитеков был впервые описан в 1905 году Гербертом Стэндингом по четырём черепам, найденным в Ампасамбазимбе и принадлежащими представителям вида M. pithecoides. Давая такое родовое имя, Стэндинг отметил сходство черепов как с известными к тому моменту палеопропитеками (лат. Palaeopropithecus), так и с современными сифаками (лат. Propithecus). В 1971 году описанный в середине 1930-х годов вид Neopropithecus globiceps был заново классифицирован, как ещё один вид мезопропитеков.
Поскольку почти до конца XX века мезопропитеки были известны только по черепам и зубам, найденным на Южном и Центральном Мадагаскаре и похожим на черепа сифак, этот род часто относили к семейству индриевых, к которому принадлежат сифаки. В середине 1930-х годов M. pithecoides были ошибочно приписаны части скелета белолобого индри. В итоге сложилось ошибочное представление об облике этого вида, которому придали сходство с обезьянами. Только в 1948 году эта ошибка была исправлена, но такие важные детали скелета мезопропитеков, как лучевые и локтевые кости, кости таза, позвонки, долго не удавалось найти[3]. Только после того, как на плато Анкарана на севере острова был обнаружен практически полный скелет M. dolichobrachion, стало ясно, что многочисленные черты роднят мезопропитеков с ленивцевыми лемурами. Длинные передние конечности и другие признаки адаптации к висячему образу жизни, характерные для палеопропитековых, но не для индриевых, позволили более точно установить место мезопропитеков на эволюционной лестнице, как рода, относящегося к семейству палеопропитековых, но раньше других отколовшегося от основной ветви развития.
Известные виды
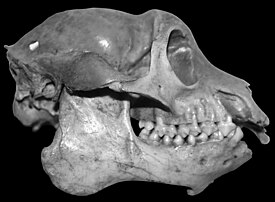
- M. pithecoides — описан в 1905 году, первым из мезопропитеков включён в классификацию. Видовое имя pithecoides (от греч. πιθηκος — обезьяна) подчёркивает сходство внешнего вида с обезьянами. M. pithecoides был лемуром средних размеров (но мелким относительно других родов палеопропитековых), весом около 10 килограммов со средней длиной черепа 9,8 см (от 9,4 до 10,3 см). Интермембральный индекс для M. pithecoides составляет 99 (передние и задние конечности примерно одной длины), гумерофеморальный индекс — 91, брахиальный индекс — 101, круральный индекс — 85. Череп M. pithecoides напоминает черепа второго открытого вида мезопропитеков, M. globiceps, но имеет более широкую лицевую часть и в целом более массивен, особенно в районе сагиттального и затылочного гребня и височных дуг. Он обитал в центральной возвышенной части Мадагаскара (местонахождения Ампасамбазимба, Анцирабе, Масинандраина), деля ареал с другими, более крупными видами ленивцевых лемуров, такими, как Archaeoindris fontoynonti и Palaeopropithecus maximus. Радиоуглеродный анализ останков одной из известных особей вида позволяет датировать их концом VII — концом VIII века новой эры[4]. Основу диеты вида составляли листья, однако она включала также плоды и реже твёрдые семена.
- M. globiceps описан в 1936 году как вид отдельного рода Neopropithecus. Видовое имя, связанное с выпуклостью лба, образовано от латинского слова globus (шар) с добавлением новолатинского суффикса -ceps, обозначающего голову. Подобно M. pithecoides, это был средних размеров лемур весом около 11 килограммов[5], со средней длиной черепа 9,4 сантиметра и интермембральным индексом 97 (гумерофеморальный индекс — 90, брахиальный — 101, круральный — 85), однако черепа и скелеты M. globiceps более изящные, приближаясь по пропорциям к скелетам современных сифак. Зубы глобицепсов напоминают зубы сифак, но в основном крупней, за исключением более коротких нижних премоляров и третьего верхнего коренного зуба. Основные отличия от современных индриевых заключаются в строении задних конечностей, позвоночного столба и таза, лучше приспособленных для висячего образа жизни, подобно таким родам ленивцевых лемуров, как палеопропитеки и бабакотии. M. globiceps обитал в центральной и западной части острова (останки обнаружены в местонахождениях Анавуха, Белу, Манумбу, Тауламбиби, Циандруина, Анказуабу, Ампасамбазимба) и питался плодами и листьями, реже разнообразя диету семенами, как современные индри. Три из найденных экземпляров M. globiceps были подвергнуты радиоуглеродному анализу и датированы периодом с середины IV века до н. э. до первой половины V века н. э.[4]
| Место мезопропитеков в систематике лемурообразных[6][7][8] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
- M. dolichobrachion впервые обнаружен в 1986 году в пещерах на плато Анкарана в северной части острова (в одно время с бабакотией) и классифицирован в 1995 году. Видовое название в переводе с греческого означает «длиннорукий», указывая на тот факт, что, в отличие от других мезопропитеков, у этого вида передние конечности были длинней задних (интермембральный индекс 113) благодаря длинной и массивной плечевой кости. Этот вид был несколько крупней двух остальных: средняя длина черепа составляет 10,2 сантиметра, предположительный вес тела — порядка 14 килограммов[5]. Помимо более длинных передних конечностей, M. dolichobrachion характеризовали более искривлённые фаланги пальцев, что позволило сделать предположение, что этот вид был лучше, чем другие, приспособлен для висячего образа жизни, подобного тому, который ведут современные ленивцы (искривлённость фаланг пальцев этого вида, известных по находкам в пещерах Анджоибе, составляет 73°, что выше, чем у любых других известных приматов, за исключением орангутанов[9]). Это предположение получило дополнительное подтверждение после анализа обнаруженного позвонка M. dolichobrachion, относящегося к поясничному отделу и схожего по форме и размерам с аналогичными позвонками другого рода палеопропитековых, приспособленного к висячему образу жизни, — бабакотий. У позвонка уменьшены остистый и поперечные отростки, относительно общих размеров меньшие, чем у палеопропитеков. Зубы M. dolichobrachion похожи на зубы двух остальных видов мезопропитеков, за исключением разницы в форме третьего верхнего коренного зуба, имеющего более широкий тригонид и более узкую дистальную поверхность. Видимо, в рацион этого вида входило больше семян, чем у двух других. M. dolichobrachion, известный по немногочисленным находкам в пределах небольшого ареала на крайнем севере Мадагаскара, делил его ещё с двумя видами ленивцевых лемуров — Babakotia radofilai и Palaeopropithecus maximus.
Анатомия и физиология
Мезопропитеки — самый мелкий из известных родов палеопропитековых, но всё же заметно крупнее живущих в наше время лемурообразных. В зависимости от вида, средний вес мезопропитеков варьировался от 10 до 14 килограммов, а длина черепа — от 9,3 до 10,6 сантиметра. Они ближе, чем остальные палеопропитековые, стоят к индриевым, напоминая тех строением черепа и скелета. Зубная формула мезопропитеков не отличается от других ленивцевых лемуров или индриевых: 2.1.2.31.1.2.3 или 2.1.2.32.0.2.3, в зависимости от того, считать ли один из нижних постоянных зубов резцом или клыком. Нижняя зубная щётка у мезопропитеков, как у индриевых и бабакотий, состояла из четырёх зубов вместо шести, характерных для многих мокроносых приматов. Постоянные зубы у этого рода, как и у прочих палеопропитековых и индриевых, развивались рано. Другие черты, роднящие мезопропитеков и бабакотий с индриевыми и отличающие их от более крупных палеопропитековых, включают увеличенные слуховую буллу (костную структуру, в которую частично заключены органы внутреннего и среднего уха) и внутреннее хрящевое кольцо, удерживающее барабанную перепонку.
Ряд важных характеристик, однако, отличает мезопропитеков от индриевых. Эти различия включают более массивный череп, в особенности в области височных дуг, сагиттального гребня и затылочного бугра. Черепная коробка более круглая, глазницы относительно меньшего размера, окружены более массивными костными дугами и ближе посажены, заглазничное сужение черепа выражено сильней, а лицевой отдел шире. Более массивная нижняя челюсть и прочный подбородочный симфиз (спайка двух половин челюсти) свидетельствуют о том, что в диете мезопропитеков листья, требующие наиболее тщательного перетирания, составляли большую часть, чем у индриевых. Относительный размер глазниц совпадает с размером глазниц у наиболее мелких из современных индриевых и свидетельствует о низкой резкости зрения. Основные различия между мезопропитеками и индриевыми касаются устройства скелета. Конечности мезопропитековых приспособлены не для скачков, как у индриевых, а для лазания, висения и передвижения на четырёх ногах: у представителей этого рода удлинены не столько нижние, сколько верхние конечности. По сравнению с другими ленивцевыми лемурами, однако, интермембральный индекс у мезопропитеков не был особо высоким, составляя в зависимости от вида от 97 до 113, что свидетельствует о их лучшей приспособленности к ходьбе на четырёх лапах и меньшей специализации к висению. Обнаруженные в 1999 году кистевые кости указывают на способность к вертикальному лазанию и больше похожи на кисть современных лориевых. Для конечностей мезопропитеков характерны удлинённые искривлённые фаланги пальцев, приспособленные для надёжного захвата и указывающие на висячий образ жизни (это характерно для всех палеопропитековых и не характерно как для современных индриевых, так и для другого семейства ископаемых лемуров — археолемуровых, чьи фаланги искривлены под вдвое меньшими углами, чем у бабакотии)[9].
Распространение и экология
Как биологический род мезопропитеки были распространены в разных областях Мадагаскара, хотя ареалы отдельных видов, возможно, не пересекались (см. Известные виды). При этом эти виды обитали в центральных и северных областях Мадагаскара одновременно с другими видами ленивцевых лемуров. Известны семь местонахождений останков M. globiceps на юге острова и по одному местонахождению останков M. pithecoides (в центральной части) и M. dolichobrachion (на севере)[4].
Если обитавший на центральном плато M. pithecoides был, по-видимому, почти исключительно листоядным, рацион двух других видов был более разнообразным, включая плоды, а временами твёрдые семена. При этом строение зубов M. dolichobrachion указывает на то, что семена в его диете занимали пропорционально большее место, чем у других видов[10].
Исчезновение
Мезопропитеки, вымершие недавно, известны по субфоссильным останкам и поэтому считаются современной формой лемурообразных. Возможно, они были одними из последних вымерших лемуров и исчезли окончательно около 500 лет назад; наиболее недавние радиоуглеродные датировки останков мезопропитеков относят к концу VI концу VII века новой эры, то есть уже после появления на Мадагаскаре людей, произошедшего около 2000 лет назад. Вероятно, охота и уничтожение человеком привычной среды обитания мезопропитеков были среди причин, приведших к их исчезновению[11]. Среди других возможных причин вымирания называется иссушение климата, приведшее к отступлению тропических лесов на Мадагаскаре. Вероятно, мезопропитеки вымерли окончательно ещё до появления на острове первых европейцев.
Примечания
- ↑ Godfrey, L. R., Jungers, W. L. Quaternary fossil lemurs // The Primate Fossil Record / Hartwig, W. C. (Ed.). — Cambridge University Press, 2002. — P. 97—121. — ISBN 978-0-521-66315-1.
- ↑ Каландадзе Н. Н., Шаповалов А. В. Судьба мегафауны неземных экосистем Мадагаскарской зоогеографической области в позднем антропогене. Проблемы эволюции. Дата обращения: 6 марта 2017. Архивировано 21 июня 2018 года.
- ↑ Godfrey, L. R., Jungers, W. L. The Extinct Sloth Lemurs of Madagascar // Evolutionary Anthropology. — 2003. — Vol. 12. — P. 252—263. — doi:10.1002/evan.10123. Архивировано 17 июля 2011 года.
- ↑ 1 2 3 Godfrey, L. R., Jungers, W. L., Burney, D. A. Subfossil Lemurs of Madagascar // Cenozoic Mammals of Africa / Werdelin, L., Sanders, W. J. (Eds.). — California Press, 2010. — ISBN 978-0-520-25721-4.
- ↑ 1 2 Jungers, W. L., Demes, B., and Godfrey, L. R. How big were the "giant" extinct lemurs of Madagascar? // Elwyn Simons: A search for origins / John G. Fleagle, Christopher C. Gilbert (Eds.). — , New York, NY: Springer, 2008. — P. 350. — ISBN 978-0-387-73895-6.
- ↑ Godfrey, L. R., Jungers, W. L. Subfossil Lemurs // The Natural History of Madagascar / Goodman, S. M., Benstead, J. P. (Eds.). — University of Chicago Press, 2003. — P. 1247—1252. — ISBN 0-226-30306-3.
- ↑ Horvath, J. E. et al. Development and Application of a Phylogenomic Toolkit: Resolving the Evolutionary History of Madagascar's Lemurs // Genome Research. — 2008. — Vol. 18, № 3. — P. 489—499. — doi:10.1101/gr.7265208. — PMID 18245770. Архивировано 24 августа 2013 года.
- ↑ Orlando, L., Calvignac, S., Schnebelen, C., Douady, C. J., Godfrey, L. R., Hänni, C. DNA from extinct giant lemurs links archaeolemurids to extant indriids // BMC Evolutionary Biology. — 2008. — Vol. 8, № 121. — doi:10.1186/1471-2148-8-121. — PMID 18442367. Архивировано 18 января 2012 года.
- ↑ 1 2 Jungers, W. L., Godfrey, L. R., Simons, E. L., Chatrath, P. S. Phalangeal curvature and positional behavior in extinct sloth lemurs (Primates, Palaeopropithecidae) // Proceedings of the National Academy of Sciences. — 1997. — Vol. 94, № 22. — P. 11998—12001. — doi:10.1073/pnas.94.22.11998. — PMID 11038588. Архивировано 24 сентября 2015 года.
- ↑ Godfrey, L. R., Semprebon, G. M., Jungers, W. L., Sutherland, M. R., Simons, E. L., Solounias, N. Dental use wear in extinct lemurs: evidence of diet and niche differentiation // Journal of Human Evolution. — 2004. — Vol. 47, № 3. — P. 145—169. — doi:10.1016/j.jhevol.2004.06.003. — PMID 15337413. Архивировано 30 марта 2012 года.
- ↑ Nowak, R. M. Family Palaeopropithecidae: Sloth Lemurs // Walker's Mammals of the World. — 6th ed. — Johns Hopkins University Press, 1999. — Vol. 2. — P. 529—531. — ISBN 0-8018-5789-9.
Ссылки
- Мезопропитек на сайте «Век млекопитающих»
Вымершие приматы | |||||||
|---|---|---|---|---|---|---|---|
| Базальные таксоны | |||||||
| Вымершие полуобезьяны |
| ||||||
| Вымершие обезьяны | |||||||
| Гоминиды | |||||||













