Метаболизм лекарств
Метаболизм лекарств — это метаболическое расщепление лекарств живыми организмами, обычно с помощью специализированных ферментных систем. В более общем смысле метаболизм ксенобиотиков (от греч. xenos — ‘чужой’ и biotic — ‘относящийся к живым существам’) — это набор метаболических путей, изменяющих химическую структуру ксенобиотиков, которые являются соединениями, чуждыми нормальной биохимии организма, например любой наркотик или яд. Эти пути, считающиеся древними, представляют собой форму биотрансформации, присутствующую во всех основных группах организмов. Эти реакции часто служат для детоксикации ядовитых соединений (хотя в некоторых случаях промежуточные продукты метаболизма ксенобиотиков могут сами вызывать токсические эффекты). Изучение метаболизма лекарств называется фармакокинетикой.
Метаболизм фармацевтических препаратов является важным аспектом фармакологии и медицины. Например, скорость метаболизма определяет продолжительность и интенсивность фармакологического действия лекарства. Метаболизм лекарств также влияет на множественную лекарственную устойчивость при инфекционных заболеваниях и при химиотерапии рака, а действие некоторых лекарств как субстратов или ингибиторов ферментов, участвующих в метаболизме ксенобиотиков, является распространенной причиной опасных лекарственных взаимодействий. Эти пути также важны для экологии, поскольку метаболизм ксенобиотиков в микроорганизмах определяет, будет ли загрязняющее вещество разрушено в процессе биоремедиации или сохранится в окружающей среде. Ферменты метаболизма ксенобиотиков, особенно глутатион S-трансферазы, также важны в сельском хозяйстве, поскольку они могут вызывать устойчивость к пестицидам и гербицидам.
Метаболизм лекарств делится на три фазы. В первой фазе ферменты, такие как оксидазы цитохрома Р450, вводят реактивные или полярные группы в ксенобиотики. Эти модифицированные соединения затем конъюгируются с полярными соединениями в реакциях фазы II. Эти реакции катализируются ферментами-трансферазами, такими как глутатион S-трансферазы. В фазе III конъюгированные ксенобиотики могут подвергаться дальнейшей обработке, после чего распознаются эффлюксными транспортерами и выводятся из клеток. Метаболизм лекарств часто преобразует липофильные соединения в гидрофильные продукты, которые легче выводятся из организма.
Барьеры проницаемости и детоксикация
Точные соединения, которым подвергается организм, в значительной степени непредсказуемы и могут сильно меняться со временем; это основные характеристики ксенобиотического токсического стресса [1]. Основная проблема, с которой сталкиваются системы детоксикации ксенобиотиков, заключается в том, что они должны быть способны удалить практически неограниченное количество ксенобиотических соединений из сложной смеси химических веществ, участвующих в нормальном метаболизме. Решение этой проблемы представляет собой элегантную комбинацию физических барьеров и низкоспецифичных ферментных систем.
Все организмы используют клеточные мембраны как гидрофобные барьеры проницаемости для контроля доступа к своей внутренней среде. Полярные соединения не могут диффундировать через эти клеточные мембраны, и поглощение полезных молекул происходит через транспортные белки, которые специально выбирают субстраты из внеклеточной смеси. Такое избирательное поглощение означает, что большинство гидрофильных молекул не могут попасть в клетки, поскольку они не распознаются никакими специфическими переносчиками [2]. Диффузия гидрофобных соединений через эти барьеры, напротив, не может контролироваться, и поэтому организмы не в состоянии исключить жирорастворимые ксенобиотики с помощью мембранных барьеров.
Однако существование барьера проницаемости означает, что организмы смогли развить системы детоксикации, которые используют гидрофобность, характерную для мембранопроницаемых ксенобиотиков. Таким образом, эти системы решают проблему специфичности, обладая настолько широкой субстратной специфичностью, что метаболизируют практически любое неполярное соединение [1]. Полезные метаболиты исключены, поскольку они полярны и, как правило, содержат одну или несколько заряженных групп.
Детоксикация реактивных побочных продуктов нормального метаболизма не может быть достигнута с помощью систем, описанных выше, поскольку эти виды получены из нормальных клеточных компонентов и обычно имеют общие полярные характеристики. Однако, поскольку этих соединений немного, специфические ферменты могут распознавать и удалять их. Примерами таких специфических систем детоксикации являются система глиоксалазы, которая удаляет реактивный альдегид метилглиоксаль [3], а также различные антиоксидантные системы, которые удаляют реактивные виды кислорода [4].
Фазы детоксикации
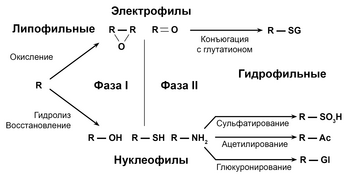
Метаболизм ксенобиотиков часто разделяют на три фазы: модификация, конъюгация и выведение. Эти реакции действуют согласованно для детоксикации ксенобиотиков и удаления их из клеток
Фаза I — модификация
В фазе I различные ферменты вступают в реактивные и полярные группы на своих субстратах. Одной из наиболее распространенных модификаций является гидроксилирование, катализируемое цитохром Р-450-зависимой оксидазной системой со смешанной функцией. Эти ферментные комплексы включают атом кислорода в неактивированные углеводороды, что может привести либо к присоединению гидроксильных групп, либо к N-, O- и S-деалкилированию субстратов [5]. Механизм реакции оксидаз P-450 протекает через восстановление связанного с цитохромом кислорода и образование высокореактивного оксиферрильного вида по следующей схеме [6]:
- O2 + НАДФH + H + + RH → НАДФ + + H2O + ROH
Реакции фазы I (также называемые несинтетическими реакциями) могут происходить путем окисления, восстановления, гидролиза, циклизации, дециклизации, присоединения кислорода или удаления водорода, осуществляемых оксидазами смешанной функции, часто в печени. В этих окислительных реакциях обычно участвуют монооксигеназа цитохрома P450 (часто сокращенно CYP), NADPH и кислород. К классам фармацевтических препаратов, которые используют этот метод для метаболизма, относятся фенотиазины, парацетамол и стероиды. Если метаболиты реакции фазы I достаточно полярны, они могут быть легко выведены из организма на этом этапе. Однако многие продукты I фазы не выводятся быстро и подвергаются последующей реакции, в которой эндогенный субстрат соединяется с вновь присоединенной функциональной группой, образуя высокополярный конъюгат.
Обычное окисление фазы I включает преобразование связи C-H в C-OH. Эта реакция иногда превращает фармакологически неактивное соединение (пролекарство) в фармакологически активное. Таким же образом фаза I может превратить нетоксичную молекулу в ядовитую (токсификация). Простой гидролиз в желудке обычно является безобидной реакцией, однако бывают и исключения. Например, метаболизм I фазы превращает ацетонитрил в HOCH2CN, который быстро диссоциирует на формальдегид и цианистый водород [7].
Метаболизм I фазы кандидатов в лекарственные препараты может быть смоделирован в лаборатории с использованием неферментных катализаторов [8]. Этот пример биомиметической реакции, как правило, дает продукты, которые часто содержат метаболиты I фазы. Например, основной метаболит фармацевтического препарата тримебутин, десметилтримебутин (нор-тримебутин), может быть эффективно получен путем окисления in vitro коммерчески доступного препарата. Гидроксилирование N-метильной группы приводит к выделению молекулы формальдегида, в то время как окисление O-метильных групп происходит в меньшей степени.
Окисление
- Цитохром P450 монооксигеназная система
- Флавинсодержащая монооксигеназная система
- Алкогольдегидрогеназа и альдегиддегидрогеназа
- Моноаминоксидаза
- Коокисление с помощью пероксидаз
Восстановление
- НАДФ-цитохром-P450-редуктаза
Цитохром-P450-редуктаза, также известная как НАДФH-ферригемопротеиновая оксидоредуктаза, НАДФH-гемопротеиновая оксидоредуктаза, НАДФH-P450-оксидоредуктаза, P450-редуктаза, POR, CPR, CYPOR, является мембраносвязанным ферментом, необходимым для переноса электронов на цитохром P450 в микросоме эукариотической клетки от ФАД- и ФМН-содержащего фермента НАДФH-цитохром-P450-редуктазы Общая схема потока электронов в системе POR/P450 такова: НАДФH → ФАД → ФМН → P450 → O2.
- Восстановленный (железистый) цитохром Р450.
Во время восстановительных реакций химическое вещество может вступать в бесполезную цикличность, при которой оно приобретает свободнорадикальный электрон, а затем быстро теряет его в пользу кислорода (с образованием супероксид-аниона).
Гидролиз
- Эстеразы и амидазы
- Эпоксид гидролаза
Фаза II — конъюгация
В последующих реакциях фазы II эти активированные метаболиты ксенобиотиков конъюгируются с заряженными видами, такими как глутатион (GSH), сульфат, глицин или глюкуроновая кислота. Места на препаратах, где происходят реакции конъюгации, включают карбокси- (-COOH), гидрокси- (-OH), амино- (NH2 ) и тиоловые (-SH) группы. Продукты реакций конъюгации имеют повышенную молекулярную массу и, как правило, менее активны, чем их субстраты. В отличие от реакций фазы I, которые часто приводят к образованию активных метаболитов. Добавление больших анионных групп (таких как GSH) детоксифицирует реактивные электрофилы и производит более полярные метаболиты, которые не могут диффундировать через мембраны и, следовательно, могут активно транспортироваться.
Эти реакции катализируются большой группой трансфераз с широкой специфичностью, которые в совокупности могут метаболизировать практически любое гидрофобное соединение, содержащее нуклеофильные или электрофильные группы [1].Одним из наиболее важных классов этой группы являются глутатион S-трансферазы (GSTs).
| Механизм | Вовлеченный фермент | Кофактор | Расположение | Источники |
|---|---|---|---|---|
| Метилирование | Метилтрансфераза | S-аденозил-L-метионин | Печень, почки, легкие, ЦНС | [9] |
| Сульфатация | Сульфотрансферазы | 3'-фосфоаденозин-5'-фосфосульфат | Печень, почки, кишечник | [9] |
| Ацетилирование |
| Ацетил кофермент А | Печень, легкие, селезенка, слизистая желудка, эритроциты, лимфоциты | [9] |
| Глюкуронирование | UDP-глюкуронозилтрансферазы | UDP-глюкуроновая кислота | Печень, почки, кишечник, легкие, кожа, простата, головной мозг | [9] |
| Конъюгация глутатиона | Глутатион S-трансферазы | Глутатион | Печень, почки | [9] |
| Конъюгация с глицином | Двухэтапный процесс:
| Глицин | Печень, почки | [10] |
Фаза III — дальнейшая модификация и выведение
После реакции фазы II конъюгаты ксенобиотиков могут подвергаться дальнейшему метаболизму. Обычным примером является переработка коньюгатов глутатиона в коньюгаты ацетилцистеина (меркаптуровой кислоты) [11]. Здесь остатки γ-глутамата и глицина в молекуле глутатиона удаляются гамма-глутамилтранспептидазой и дипептидазами. На последнем этапе остаток цистеина в конъюгате ацетилируется.
Конъюгаты и их метаболиты могут выводиться из клеток в III фазе метаболизма, при этом анионные группы действуют как аффинные метки для различных мембранных транспортеров семейства белков множественной лекарственной устойчивости (MRP) [12]. Эти белки являются членами семейства АТФ-связывающих кассетных транспортеров и могут катализировать АТФ-зависимый транспорт огромного разнообразия гидрофобных анионов [13], таким образом действуя для удаления продуктов II фазы во внеклеточную среду, где они могут быть подвергнуты дальнейшему метаболизму или выведены из организма [14].
Эндогенные токсины
Детоксикация эндогенных реактивных метаболитов, таких как пероксиды и реактивные альдегиды, часто не может быть достигнута системой, описанной выше. Это результат того, что эти виды являются производными нормальных клеточных компонентов и обычно имеют общие полярные характеристики. Однако, поскольку этих соединений немного, ферментативные системы могут использовать специфическое молекулярное распознавание для их распознавания и удаления. Сходство этих молекул с полезными метаболитами означает, что различные ферменты детоксикации обычно требуются для метаболизма каждой группы эндогенных токсинов. Примерами таких специфических систем детоксикации являются система глиоксалазы, которая утилизирует реактивный альдегид метилглиоксаль, и различные антиоксидантные системы, удаляющие реактивные виды кислорода.
Места метаболизма
В количественном отношении гладкий эндоплазматический ретикулум клеток печени является главным органом метаболизма лекарств, хотя любая биологическая ткань обладает определенной способностью к метаболизму лекарств. Вклад печени в метаболизм обусловлен тем, что это большой орган, что это первый орган, через который проходят химические вещества, поглощенные в кишечнике, и что здесь очень высокая концентрация большинства ферментных систем, метаболизирующих лекарства, по сравнению с другими органами. Если лекарство попадает в ЖКТ, где оно поступает в печеночную циркуляцию через воротную вену, оно хорошо метаболизируется и проявляет эффект первого прохождения.
Другие места метаболизма лекарств включают эпителиальные клетки желудочно-кишечного тракта, легких, почек и кожи. Эти участки обычно ответственны за локализованные реакции токсичности.
Факторы, влияющие на метаболизм лекарств
Продолжительность и интенсивность фармакологического действия большинства липофильных препаратов определяется скоростью их метаболизма до неактивных продуктов. Система монооксигеназ цитохрома Р450 является наиболее важным путем в этом отношении. В целом все, что увеличивает скорость метаболизма (например, индукция ферментов) фармакологически активного метаболита, уменьшает продолжительность и интенсивность действия препарата. Обратное также верно (например, ингибирование фермента). Однако в тех случаях, когда фермент отвечает за метаболизм пролекарства в лекарство, индукция фермента может ускорить это превращение и повысить уровень метаболизма препарата, потенциально вызывая токсичность.
Различные физиологические и патологические факторы также могут влиять на метаболизм лекарств. Физиологические факторы, которые могут влиять на метаболизм лекарств, включают возраст, индивидуальные различия (например, фармакогенетика), энтерогепатическую циркуляцию, питание, кишечную флору или половые различия.
В целом у плода, новорожденного, пожилых людей и животных лекарства метаболизируются медленнее, чем у взрослых.
Генетические вариации (полиморфизм) объясняют некоторую изменчивость в действии лекарств. В случае N-ацетилтрансфераз, участвующих в реакциях фазы II, индивидуальные различия создают группу людей, которые ацетилируют медленно (медленные ацетиляторы), и тех, кто ацетилирует быстро. Соотношение быстрых и медленных метаболизаторов в разных регионах может быть разным. Эта вариация может иметь драматические последствия, поскольку медленные ацетиляторы более склонны к дозозависимой токсичности.
Ферменты монооксигеназной системы цитохрома P450 также могут отличаться у разных людей, их дефицит встречается у 1–30 % людей в зависимости от их этнического происхождения.
Доза, частота, способ введения, распределение в тканях и связывание с белками влияют на метаболизм препарата.
Патологические факторы также могут влиять на метаболизм лекарств, включая заболевания печени, почек или сердца.
Методы моделирования и симуляции in silico позволяют прогнозировать метаболизм лекарств в виртуальных популяциях пациентов до проведения клинических исследований на людях [15]. Это можно использовать для выявления лиц, наиболее подверженных риску возникновения побочных реакций.
История
Изучение того, как люди преобразуют вещества, принимаемые внутрь, началось в середине XIX века, когда химики обнаружили, что органические химические вещества, такие как бензальдегид, могут окисляться и конъюгироваться с аминокислотами в организме человека [16]. В течение оставшейся части XIX века было открыто несколько других основных реакций детоксикации, таких как метилирование, ацетилирование и сульфонирование.
В начале XX века работа перешла к исследованию ферментов и путей, которые отвечают за производство этих метаболитов. Эта область выделилась в отдельную область исследований после публикации Ричардом Уильямсом книги «Механизмы детоксикации» в 1947 году [17]. Эти современные биохимические исследования привели к идентификации глутатион S-трансфераз в 1961 году [18], затем к открытию цитохрома P450 в 1962 году [19] и осознанию их центральной роли в метаболизме ксенобиотиков в 1963 году [20][21][21].
Смотрите также
- Антиоксидант
- Биодеградация
- Биоремедиация
- Микробная биодеградация
Ссылки
- ↑ 1 2 3 "The enzymes of detoxication". J. Biol. Chem. 265 (34): 20715—8. December 1990. doi:10.1016/S0021-9258(17)45272-0. PMID 2249981. Архивировано из оригинала 21 июня 2009. Дата обращения: 29 декабря 2012.
- ↑ "Impact of drug transporter studies on drug discovery and development". Pharmacol. Rev. 55 (3): 425—61. September 2003. doi:10.1124/pr.55.3.1. PMID 12869659.
- ↑ Thornalley PJ (July 1990). "The glyoxalase system: new developments towards functional characterization of a metabolic pathway fundamental to biological life". Biochem. J. 269 (1): 1—11. doi:10.1042/bj2690001. PMID 2198020.
- ↑ Sies H (March 1997). "Oxidative stress: oxidants and antioxidants". Exp. Physiol. 82 (2): 291—5. doi:10.1113/expphysiol.1997.sp004024. PMID 9129943.
- ↑ Guengerich FP (June 2001). "Common and uncommon cytochrome P450 reactions related to metabolism and chemical toxicity". Chem. Res. Toxicol. 14 (6): 611—50. doi:10.1021/tx0002583. PMID 11409933.
- ↑ "The catalytic pathway of cytochrome p450cam at atomic resolution". Science. 287 (5458): 1615—22. March 2000. Bibcode:2000Sci...287.1615S. doi:10.1126/science.287.5458.1615. PMID 10698731.
- ↑ Acetonitrile (EHC 154, 1993). www.inchem.org. Дата обращения: 3 мая 2017. Архивировано 22 мая 2017 года.
- ↑ "Oxidation of antiparasitic 2-substituted quinolines using metalloporphyrin catalysts: scale-up of a biomimetic reaction for metabolite production of drug candidates". Org. Biomol. Chem. 6 (24): 4494—7. December 2008. doi:10.1039/b815963g. PMID 19039354.
- ↑ 1 2 3 4 5 "Drug glucuronidation in clinical psychopharmacology". J Clin Psychopharmacol. 21 (5): 500—15. October 2001. doi:10.1097/00004714-200110000-00008. PMID 11593076.
- ↑ "Glycine conjugation: importance in metabolism, the role of glycine N-acyltransferase, and factors that influence interindividual variation". Expert Opinion on Drug Metabolism & Toxicology. 9 (9): 1139—1153. September 2013. doi:10.1517/17425255.2013.796929. PMID 23650932.
Glycine conjugation of mitochondrial acyl-CoAs, catalyzed by glycine N-acyltransferase (GLYAT, E.C. 2.3.1.13), is an important metabolic pathway responsible for maintaining adequate levels of free coenzyme A (CoASH). However, because of the small number of pharmaceutical drugs that are conjugated to glycine, the pathway has not yet been characterized in detail. Here, we review the causes and possible consequences of interindividual variation in the glycine conjugation pathway. ...
Figure 1. Glycine conjugation of benzoic acid. The glycine conjugation pathway consists of two steps. First benzoate is ligated to CoASH to form the high-energy benzoyl-CoA thioester. This reaction is catalyzed by the HXM-A and HXM-B medium-chain acid:CoA ligases and requires energy in the form of ATP. ... The benzoyl-CoA is then conjugated to glycine by GLYAT to form hippuric acid, releasing CoASH. In addition to the factors listed in the boxes, the levels of ATP, CoASH, and glycine may influence the overall rate of the glycine conjugation pathway. - ↑ "The role of glutathione and glutathione S-transferases in mercapturic acid biosynthesis". Adv. Enzymol. Relat. Areas Mol. Biol. 32: 173—219. 1969. doi:10.1002/9780470122778.ch5. ISBN 9780470122778. PMID 4892500.
- ↑ "Multidrug resistance-associated proteins: Export pumps for conjugates with glutathione, glucuronate or sulfate". BioFactors. 17 (1—4): 103—14. 2003. doi:10.1002/biof.5520170111. PMID 12897433.
- ↑ "Conjugate export pumps of the multidrug resistance protein (MRP) family: localization, substrate specificity, and MRP2-mediated drug resistance". Biochim. Biophys. Acta. 1461 (2): 377—94. December 1999. doi:10.1016/S0005-2736(99)00169-8. PMID 10581368.
- ↑ "Enzymes and transport systems involved in the formation and disposition of glutathione S-conjugates. Role in bioactivation and detoxication mechanisms of xenobiotics". Pharmacol. Rev. 47 (2): 271—330. June 1995. PMID 7568330.
- ↑ "Simulation and prediction of in vivo drug metabolism in human populations from in vitro data". Nat Rev Drug Discov. 6 (2): 140—8. February 2007. doi:10.1038/nrd2173. PMID 17268485.
- ↑ Murphy PJ (June 2001). "Xenobiotic metabolism: a look from the past to the future". Drug Metab. Dispos. 29 (6): 779—80. PMID 11353742. Архивировано из оригинала 21 июня 2009. Дата обращения: 29 декабря 2012.
- ↑ "Richard Tecwyn Williams: the man, his work, his impact". Drug Metab. Rev. 14 (3): 559—607. 1983. doi:10.3109/03602538308991399. PMID 6347595.
- ↑ "An enzyme from rat liver catalysing conjugations with glutathione". Biochem. J. 79 (3): 516—24. June 1961. doi:10.1042/bj0790516. PMID 16748905.
- ↑ "A new cytochrome in liver microsomes". J. Biol. Chem. 237 (4): 1375—6. April 1962. doi:10.1016/S0021-9258(18)60338-2. PMID 14482007. Архивировано из оригинала 21 июня 2009. Дата обращения: 29 декабря 2012.
- ↑ Estabrook RW (December 2003). "A passion for P450s (remembrances of the early history of research on cytochrome P450)". Drug Metab. Dispos. 31 (12): 1461—73. doi:10.1124/dmd.31.12.1461. PMID 14625342.
- ↑ 1 2 "The light reversible carbon monoxide inhibition of steroid C-21 hydroxylase system in adrenal cortex". Biochem Z. 338: 741—55. 1963. PMID 14087340.