Митохондриальная ДНК
Митохондриа́льная ДНК (мтДНК) — ДНК, находящаяся (в отличие от ядерной ДНК) в митохондриях, органеллах эукариотических клеток.
Гены, закодированные в митохондриальной ДНК, относятся к группе плазмогенов, расположенных вне ядра (вне хромосомы). Совокупность этих факторов наследственности, сосредоточенных в цитоплазме клетки, составляет плазмон данного вида организмов (в отличие от генома)[1].
История открытия
Митохондриальная ДНК была открыта Маргит Насс и Сильвен Насс в 1963 году в Стокгольмском университете при помощи электронной микроскопии[2] и, независимо, учёными Эллен Харлсбруннер, Хансом Туппи и Готтфридом Шацем при биохимическом анализе фракций митохондрий дрожжей в Венском университете в 1964 году.[3]
Теории возникновения митохондриальной ДНК
Согласно эндосимбиотической теории, митохондриальная ДНК произошла от кольцевых молекул ДНК бактерий и поэтому имеет иное происхождение, чем ядерный геном. Сейчас преобладает точка зрения, согласно которой митохондрии имеют монофилетическое происхождение, то есть были приобретены предками эукариот лишь однажды.
На основании сходства в последовательностях нуклеотидов ДНК ближайшими родственниками митохондрий среди ныне живущих прокариот считают альфа-протеобактерий (в частности, выдвигалась гипотеза, что к митохондриям близки риккетсии). Сравнительный анализ геномов митохондрий показывает, что в ходе эволюции происходило постепенное перемещение генов предков современных митохондрий в ядро клетки. Необъяснимыми с эволюционной точки зрения остаются некоторые особенности митохондриальной ДНК (например, довольно большое число интронов, нетрадиционное использование триплетов и другие). Ввиду ограниченного размера митохондриального генома бо́льшая часть митохондриальных белков кодируется в ядре. При этом бо́льшая часть митохондриальных тРНК кодируются митохондриальным геномом.
Формы и число молекул митохондриальной ДНК
У большинства изученных организмов митохондрии содержат только кольцевые молекулы ДНК, у некоторых растений одновременно присутствуют и кольцевые, и линейные молекулы, а у ряда протистов (например, инфузорий) имеются только линейные молекулы.[5]
Митохондрии млекопитающих обычно содержат от двух до десяти идентичных копий кольцевых молекул ДНК.[6]
У растений каждая митохондрия содержит несколько молекул ДНК разного размера, которые способны к рекомбинации.
У протистов из отряда кинетопластид (например, у трипаносом) в особом участке митохондрии (кинетопласте) содержится два типа молекул ДНК — идентичные макси-кольца (20-50 штук) длиной около 21 т. п. о. и мини-кольца (20 000-55 000 штук, около 300 разновидностей, средняя длина около 1000 п. о.). Все кольца соединены в единую сеть (катенаны), которая разрушается и восстанавливается при каждом цикле репликации. Макси-кольца гомологичны митохондриальной ДНК других организмов. Каждое мини-кольцо содержит четыре сходных консервативных участка и четыре уникальных гипервариабельных участка.[7] В мини-кольцах закодированы короткие молекулы направляющих РНК (guideRNA), которые осуществляют редактирование РНК, транскрибируемых с генов макси-колец.
Устойчивость митохондриальной ДНК
Митохондриальная ДНК особенно чувствительна к активным формам кислорода, генерируемым дыхательной цепью, в связи с непосредственной их близостью. Хотя митохондриальная ДНК связана с белками, их защитная роль менее выражена, чем в случае ядерной ДНК. Мутации в ДНК митохондрий могут вызывать передаваемые по материнской линии наследственные заболевания. Также имеются данные, указывающие на возможный вклад мутаций митохондриальной ДНК в процесс старения и развитие возрастных патологий.[8] У человека митохондриальная ДНК обычно присутствует в количестве 100—10000 копий на клетку (сперматозоиды и яйцеклетки являются исключением). С множественностью митохондриальных геномов связаны особенности проявления митохондриальных болезней — обычно позднее их начало и очень изменчивые симптомы.
Митохондриальная наследственность
Наследование по материнской линии
У большинства многоклеточных организмов митохондриальная ДНК наследуется по материнской линии. Яйцеклетка содержит на несколько порядков больше копий митохондриальной ДНК, чем сперматозоид. В сперматозоиде обычно не больше десятка митохондрий (у человека — одна спирально закрученная митохондрия), в небольших яйцеклетках морского ежа — несколько сотен тысяч, а в крупных ооцитах лягушки — десятки миллионов. Кроме того, обычно происходит деградация митохондрий сперматозоида после оплодотворения[9].
При половом размножении митохондрии, как правило, наследуются исключительно по материнской линии, митохондрии сперматозоида обычно разрушаются после оплодотворения. Кроме того, большая часть митохондрий сперматозоида находятся в основании жгутика, которое при оплодотворении иногда теряется. В 1999 году было обнаружено, что митохондрии сперматозоидов помечены убиквитином (белком-меткой, которая приводит к разрушению отцовских митохондрий в зиготе)[10].
Так как митохондриальная ДНК не является высококонсервативной и имеет высокую скорость мутирования[англ.], она является хорошим объектом для изучения филогении (эволюционного родства) живых организмов. Для этого определяют последовательности митохондриальной ДНК у разных видов и сравнивают их при помощи специальных компьютерных программ и получают эволюционное древо для изученных видов. Исследование митохондриальных ДНК собак позволило проследить происхождение собак от диких волков[11]. Исследование митохондриальной ДНК в популяциях человека позволило вычислить «митохондриальную Еву», гипотетическую прародительницу всех живущих в настоящее время людей.
Наследование по отцовской линии
Для некоторых видов показана передача митохондриальной ДНК по мужской линии, например, у мидий[12][13]. Наследование митохондрий по отцовской линии также описано для некоторых насекомых, например, для дрозофилы,[14] медоносных пчел[15] и цикад.[16]
Существуют также данные о митохондриальном наследовании по мужской линии у млекопитающих. Описаны случаи такого наследования для мышей,[17][18] при этом митохондрии, полученные от самца, впоследствии отторгаются. Такое явление показано для овец [19] и клонированного крупного рогатого скота.[20]
Наследование по отцовской линии у людей
До недавнего времени считалось, что митохондрии человека наследуются только по материнской линии. Был известен лишь один-единственный случай пациента, у которого в 2002 году достоверно обнаружили отцовскую митохондриальную ДНК[21].
Лишь недавнее исследование 2018 года показало, что митохондриальная ДНК человека иногда всё же может передаваться и по отцовской линии. Небольшое количество митохондрий отца может попасть в яйцеклетку матери вместе с цитоплазмой сперматозоида, но, как правило, отцовские митохондрии после этого из зиготы исчезают. Однако, было обнаружено, что у некоторых людей существует «мутация, которая помогает выживать митохондриям отца»[22].
Геном митохондрий
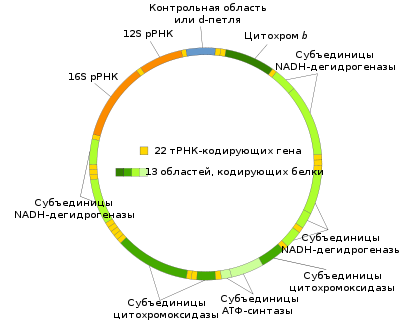
У млекопитающих каждая молекула мтДНК содержит 15000-17000 пар оснований (у человека 16565 пар нуклеотидов — исследование закончено в 1981 году[23], по другому источнику 16569 пар[24]) и содержит 37 генов — 13 кодируют белки[25], 22 — гены тРНК, 2 — рРНК (по одному гену для 12S и 16S рРНК). Другие многоклеточные животные имеют схожий набор митохондриальных генов, хотя некоторые гены могут иногда отсутствовать. Генный состав мтДНК разных видов растений, грибов и особенно протистов [26] различается более значительно. Так, у жгутиконосца-якобиды Reclinomonas americana найден наиболее полный из известных митохондриальных геномов: он содержит 97 генов, в том числе 62 гена, кодирующих белки (27 рибосомных белков, 23 белка, участвующих в работе электрон-транспортной цепи и в окислительном фосфорилировании, а также субъединицы РНК-полимеразы).
Один из наиболее маленьких митохондриальных геномов имеет малярийный плазмодий (около 6.000 п.о., содержит два гена рРНК и три гена, кодирующих белки).
Недавно открытые рудиментарные митохондрии (митосомы) некоторых протистов (дизентерийной амёбы, микроспоридий и лямблий) не содержат ДНК.
Митохондриальные геномы различных видов грибов содержат от 19 431 (делящиеся дрожжи Schizosaccharomyces pombe) до 100 314 (сордариомицет Podospora anserina) пар нуклеотидов[27].
Некоторые растения имеют огромные молекулы митохондриальной ДНК (до 25 миллионов пар оснований), при этом содержащие примерно те же гены и в том же количестве, что и меньшие мтДНК. Длина митохондриальной ДНК может широко варьировать даже у растений одного семейства. В митохондриальной ДНК растений имеются некодирующие повторяющиеся последовательности.
Геном человека содержит только по одному промотору на каждую комплементарную цепь ДНК[23].
Геном митохондрий человека кодирует следующие белки и РНК:
| Белки или РНК | Гены |
| NADH-дегидрогеназа (комплекс I) | MT-ND1, MT-ND2, MT-ND3, MT-ND4, MT-ND4L, MT-ND5, MT-ND6 |
| Кофермент Q - цитохром c редуктаза/Цитохром b (комплекс III) | MT-CYB |
| цитохром c оксидаза (комплекс IV) | MT-CO1, MT-CO2, MT-CO3 |
| АТФ-синтаза | MT-ATP6, MT-ATP8 |
| рРНК | MT-RNR1 (12S), MT-RNR2 (16S) |
| тРНК | MT-TA, MT-TC, MT-TD, MT-TE, MT-TF, MT-TG, MT-TH, MT-TI, MT-TK, MT-TL1, MT-TL2, MT-TM, MT-TN, MT-TP, MT-TQ, MT-TR, MT-TS1, MT-TS2, MT-TT, MT-TV, MT-TW, MT-TY, MT1X |
Особенности митохондриальной ДНК
Кодирующие последовательности (кодоны) митохондриального генома имеют некоторые отличия от кодирующих последовательностей универсальной ядерной ДНК.
Так, кодон AUA кодирует в митохондриальном геноме метионин (вместо изолейцина в ядерной ДНК), кодоны AGA и AGG — терминаторные кодоны (в ядерной ДНК кодируют аргинин), кодон UGA в митохондриальном геноме кодирует триптофан[23].
Если говорить точнее, то речь идёт не о митохондриальной ДНК, а о мРНК, которая списывается (транскрибируется) с этой ДНК перед началом синтеза белка. Буква U в обозначении кодона обозначает урацил, который при транскрипции гена в РНК заменяет тимин.
Количество генов тРНК (22 гена) меньше, чем в ядерном геноме с его 32 генами тРНК[23].
В человеческом митохондриальном геноме информация настолько сконцентрирована, что в последовательностях, кодирующих мРНК, как правило, частично удалены нуклеотиды, соответствующие 3'-концевым терминаторным кодонам[23].
Применение
Кроме использования при построении различных филогенетических теорий, изучение митохондриального генома — основной инструмент при проведении идентификации. Возможность идентификации связана с существующими в митохондриальном геноме человека групповыми и даже индивидуальными различиями.
Последовательность участка гена субъединицы I цитохром с-оксидазы, кодируемого в митохондриальной ДНК, широко используется в проектах, связанных с ДНК-баркодированием животных — определением принадлежности организма к тому или иному таксону на основе коротких маркеров в его ДНК[28][29]. Для баркодирования растений используется преимущественно комбинация двух маркёров в пластидной ДНК[30].
Группа Шухрата Миталипова из центра эмбриональных клеток и генной терапии Орегонского университета разработала метод замены митохондриальной ДНК для лечения наследственных митохондриальных заболеваний. Сейчас в Великобритании начаты клинические испытания этого метода, получившего неофициальное название «3-parent baby technique» — «ребёнок от трёх родителей». Известно также о рождении в результате этой процедуры ребёнка в Мексике[31].
Примечания
- ↑ Джинкс Д., Нехромосомная наследственность, пер. с англ., М., 1966; Сэджер Р., Гены вне хромосом, в кн.: Молекулы и клетки, пер. с англ., М., 1966.
- ↑ Nass, M.M. & Nass, S. (1963 at the Wenner-Gren Institute for Experimental Biology, Stockholm University, Stockholm, Sweden): Intramitochondrial Fibers with DNA characteristics Архивная копия от 10 сентября 2008 на Wayback Machine (PDF). In: J. Cell. Biol. Bd. 19, S. 593—629. PMID 14086138
- ↑ Ellen Haslbrunner, Hans Tuppy and Gottfried Schatz (1964 at the Institut for Biochemistry at the Medical Faculty of the University of Vienna in Vienna, Австрия): «Deoxyribonucleic Acid Associated with Yeast Mitochondria» (PDF) Biochem. Biophys. Res. Commun. 15, 127—132.
- ↑ Iborra F. J., Kimura H., Cook P. R. The functional organization of mitochondrial genomes in human cells (англ.) // BMC Biol.[англ.] : journal. — 2004. — Vol. 2. — P. 9. — doi:10.1186/1741-7007-2-9. — PMID 15157274. — PMC 425603. Архивировано 30 сентября 2015 года.
- ↑ Дымшиц Г. М. Сюрпризы митохондриального генома. Природа, 2002, N 6. Дата обращения: 18 мая 2014. Архивировано 28 июля 2012 года.
- ↑ Wiesner R. J., Ruegg J. C., Morano I. Counting target molecules by exponential polymerase chain reaction, copy number of mitochondrial DNA in rat tissues (англ.) // Biochim Biophys Acta.[англ.] : journal. — 1992. — Vol. 183. — P. 553—559. — PMID 1550563.
- ↑ doi:10.1016/j.exppara.2006.04.005. Дата обращения: 11 мая 2009. Архивировано из оригинала 31 марта 2011 года.
- ↑ Alexeyev, Mikhail F.; LeDoux, Susan P.; Wilson, Glenn L. Mitochondrial DNA and aging (неопр.) // Clinical Science. — 2004. — July (т. 107, № 4). — С. 355—364. — doi:10.1042/CS20040148. — PMID 15279618. Архивировано 25 сентября 2019 года.
- ↑ Ченцов Ю. С. Общая цитология. — 3-е изд. — МГУ, 1995. — 384 с. — ISBN 5-211-03055-9.
- ↑ Sutovsky, P., et. al. Ubiquitin tag for sperm mitochondria (англ.) // Nature. — Nov. 25, 1999. — Vol. 402. — P. 371—372. — doi:10.1038/46466. — PMID 10586873. Discussed in [1] Архивная копия от 19 декабря 2007 на Wayback Machine
- ↑ Vilà C., Savolainen P., Maldonado J. E., and Amorin I. R. Multiple and Ancient Origins of the Domestic Dog (англ.) // Science : journal. — 1997. — 13 June (vol. 276). — P. 1687—1689. — ISSN 0036-8075. — doi:10.1126/science.276.5319.1687. — PMID 9180076.
- ↑ Hoeh W. R., Blakley K. H., Brown W. M. Heteroplasmy suggests limited biparental inheritance of Mytilus mitochondrial DNA (англ.) // Science : journal. — 1991. — Vol. 251. — P. 1488—1490. — doi:10.1126/science.1672472. — PMID 1672472.
- ↑ Penman, Danny (2002-08-23). "Mitochondria can be inherited from both parents". NewScientist.com. Архивировано 11 октября 2008. Дата обращения: 5 февраля 2008.
- ↑ Kondo R., Matsuura E. T., Chigusa S. I. Further observation of paternal transmission of Drosophila mitochondrial DNA by PCR selective amplification method (англ.) // Genet. Res.[англ.] : journal. — 1992. — Vol. 59, no. 2. — P. 81—4. — PMID 1628820.
- ↑ Meusel M. S., Moritz R. F. Transfer of paternal mitochondrial DNA during fertilization of honeybee (Apis mellifera L.) eggs (англ.) // Curr. Genet. : journal. — 1993. — Vol. 24, no. 6. — P. 539—543. — doi:10.1007/BF00351719. — PMID 8299176.
- ↑ Fontaine, K. M., Cooley, J. R., Simon, C. Evidence for paternal leakage in hybrid periodical cicadas (Hemiptera: Magicicada spp.) (исп.) // PLoS One. : diario. — 2007. — V. 9. — P. e892. — doi:10.1371/journal.pone.0000892.
- ↑ Gyllensten U., Wharton D., Josefsson A., Wilson A. C. Paternal inheritance of mitochondrial DNA in mice (англ.) // Nature. — 1991. — Vol. 352, no. 6332. — P. 255—257. — doi:10.1038/352255a0. — PMID 1857422.
- ↑ Shitara H., Hayashi J. I., Takahama S., Kaneda H., Yonekawa H. Maternal inheritance of mouse mtDNA in interspecific hybrids: segregation of the leaked paternal mtDNA followed by the prevention of subsequent paternal leakage (англ.) // Genetics : journal. — 1998. — Vol. 148, no. 2. — P. 851—857. — PMID 9504930.
- ↑ Zhao X., Li N., Guo W., et al. Further evidence for paternal inheritance of mitochondrial DNA in the sheep (Ovis aries) (англ.) // Heredity : journal. — 2004. — Vol. 93, no. 4. — P. 399—403. — doi:10.1038/sj.hdy.6800516. — PMID 15266295.
- ↑ Steinborn R., Zakhartchenko V., Jelyazkov J., et al. Composition of parental mitochondrial DNA in cloned bovine embryos (англ.) // FEBS Lett.[англ.] : journal. — 1998. — Vol. 426, no. 3. — P. 352—356. — doi:10.1016/S0014-5793(98)00350-0. — PMID 9600265.
- ↑ Schwartz M., Vissing J. Paternal inheritance of mitochondrial DNA (англ.) // N. Engl. J. Med. : journal. — 2002. — Vol. 347, no. 8. — P. 576—580. — doi:10.1056/NEJMoa020350. — PMID 12192017.
- ↑ Митохондриальная ДНК может передаваться по отцовской линии • Полина Лосева • Новости науки на «Элементах» • Генетика, Микробиология. Дата обращения: 24 декабря 2018. Архивировано 24 декабря 2018 года.
- ↑ 1 2 3 4 5 Айала Ф. Д. Современная генетика. 1987.
- ↑ Архивированная копия. Дата обращения: 10 октября 2009. Архивировано из оригинала 13 августа 2011 года.
- ↑ Даниленко Н. Г., Давыденко О.Г — Миры геномов органелл
- ↑ MW Gray, BF Lang, R Cedergren, GB Golding, C Lemieux, D Sankoff, M Turmel, N Brossard, E Delage, TG Littlejohn, I Plante, P Rioux, D Saint-Louis, Y Zhu and G Burger. Genome structure and gene content in protist mitochondrial DNAs (англ.) // Nucleic Acids Research : journal. — 1998. — Vol. 26. — P. 865—878.http://nar.oxfordjournals.org/cgi/content/abstract/26/4/865 Архивная копия от 29 августа 2008 на Wayback Machine
- ↑ Дьяков Ю. Т., Шнырева А. В., Сергеев А. Ю. Введение в генетику грибов. — М.: изд. центр «Академия», 2005. — С. 52. — ISBN 5-7695-2174-0.
- ↑ Paul D. N. Hebert, Alina Cywinska, Shelley L. Ball, Jeremy R. deWaard. Biological identifications through DNA barcodes (англ.) // Proceedings of the Royal Society of London B: Biological Sciences. — 2003-02-07. — Vol. 270, iss. 1512. — P. 313—321. — ISSN 0962-8452. — doi:10.1098/rspb.2002.2218. Архивировано 19 апреля 2016 года.
- ↑ Živa Fišer Pečnikar, Elena V. Buzan. 20 years since the introduction of DNA barcoding: from theory to application // Journal of Applied Genetics. — 2014-02-01. — Т. 55, вып. 1. — С. 43—52. — ISSN 2190-3883. — doi:10.1007/s13353-013-0180-y.
- ↑ CBOL Plant Working Group1, Peter M. Hollingsworth, Laura L. Forrest, John L. Spouge, Mehrdad Hajibabaei. A DNA barcode for land plants (англ.) // Proceedings of the National Academy of Sciences. — National Academy of Sciences, 2009-08-04. — Vol. 106, iss. 31. — P. 12794—12797. — ISSN 0027-8424. — doi:10.1073/pnas.0905845106. Архивировано 24 мая 2016 года.
- ↑ Алла Астахова. Тонкая работа - 2. Блог о здравоохранении (22 августа 2017). Дата обращения: 23 августа 2017. Архивировано 23 августа 2017 года.
Ссылки
- Mitomap — база данных по митохондриальному геному человека
- Human mtDNA — база данных полных сиквенсов мтДНК человека
- Phylotree М.ван Овена и М.Кайзера
- Митохондриальная ДНК: пособие для начинающего молекулярного генеалога
См. также
- Митохондриальная Ева
- Гаплогруппы
- Митохондриальные заболевания
- Цитоплазматическая мужская стерильность
- Пластидная ДНК
| В библиографических каталогах |
|---|
Типы нуклеиновых кислот | ||||
|---|---|---|---|---|
| Азотистые основания | ||||
| Нуклеозиды | ||||
| Нуклеотиды | ||||
| РНК | ||||
| ДНК | ||||
| Аналоги | ||||
| Типы векторов |
| |||
| ||||