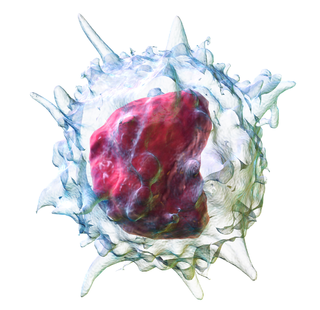
Нейтрофилы
| Нейтрофил | |
|---|---|
 | |
| Ткань | соединительная |
| История дифференцировки клетки | Зигота → Бластомер → Эмбриобласт → Эпибласт → Клетка первичной мезодермы → Прегемангиобласт → Гемангиобласт → Гемоцитобласт → Общий миелоидный прародитель → Нейтрофильный промиелоцит → Нейтрофильный миелоцит → Нейтрофильный метамиелоцит → Палочкоядерный нейтрофил → Сегментоядерный нейтрофил (нейтрофильный гранулоцит) |
Нейтрофи́лы, или нейтрофи́льные гранулоци́ты, или нейтрофи́льные сегментоя́дерные гранулоци́ты[1], или полиморфонуклеа́рные нейтрофи́лы[2], — наиболее многочисленная группа гранулоцитов, на долю которой приходится от 40 % до 70 % всех лейкоцитов у человека[3]. Нейтрофилы являются частью врождённого иммунитета[4], их основная функция — фагоцитоз патогенных микроорганизмов (бактерий, грибков, простейших) и продуктов распада тканей организма[5].
Нейтрофилы представляют собой очень подвижные клетки, которые проникают даже в те ткани, которые недоступны для других лейкоцитов. В зависимости от морфологии ядра нейтрофилы подразделяют на палочкоядерные (незрелые) и сегментоядерные (зрелые) нейтрофилы[6][7]. Развитие нейтрофилов находится под контролем цитокинов, прежде всего G-CSF[англ.], а также GM-CSF, IL-3 и IL-6. В условиях воспалительного ответа количество нейтрофилов увеличивается под действием IL-17[англ.] и IL-23[англ.][1].
Нейтрофилы являются основными фагоцитами кровотока, но в ходе острого воспаления интенсивно мигрируют в очаг воспаления[8][9][10]. Они проникают через стенки кровеносных сосудов и движутся по градиенту различных провоспалительных молекул в ходе хемотаксиса[11]. Нейтрофилы — самые многочисленные клетки, входящие в состав гноя, именно они придают ему беловатый или желтоватый цвет[12].
Структура
Нейтрофилы — наиболее многочисленный подтип лейкоцитов у человека; каждый день в организме человека образуется порядка 1011 новых нейтрофилов. В норме количество нейтрофилов в литре крови составляет от 2—2,5 до 7,5 миллиардов клеток. У людей африканского и средневосточного происхождения количество нейтрофилов может быть меньше 2,5 миллиардов на литр[13]. В кровотоке находится лишь 1—2 % зрелых нейтрофилов, все остальные локализованы в тканях. Кроме того, в крови здорового человека присутствуют незрелые палочкоядерные нейтрофилы в количестве 0,04—0,3 × 109 на литр, что соответствует 1—6 % всех нейтрофилов[1]. У человека повышение содержания палочкоядерных нейтрофилов в крови свидетельствует об остром воспалении, лейкозе или повреждении костного мозга, однако у некоторых животных существенная доля незрелых нейтрофилов в крови является нормой[14].
В мазках крови нейтрофилы имеют диаметр от 12 до 15 мкм. В суспензии человеческие нейтрофилы достигают 7—9 мкм в диаметре[15]. Неактивированные нейтрофилы, плавающие в кровотоке, имеют сферическую форму; после активации форма нейтрофилов меняется, становится амёбоидной, появляются псевдоподии, с помощью которых нейтрофилы могут захватывать антигены[16].
На поверхности нейтрофилов присутствуют молекулы CD13, служащей рецептором для некоторых вирусов, CD14 (рецептор липополисахарида), β2-интегрины (LFA-1, Mac-1 и p155/95), Fc-рецепторы (CD32[англ.] и CD16), рецепторы комплемента[англ.] (CR1[англ.], Mac-1, CR4[англ.]) и рецепторы факторов хемотаксиса. Нейтрофилы постоянно экспрессируют главный комплекс гистосовместимости I класса (MHC-I[англ.]), а под действием некоторых цитокинов, таких как GM-CSF, начинают экспрессировать MHC-II[англ.]. Нейтрофилы несут рецепторы для ключевых факторов, влияющих на их развитие, миграцию и активацию: G-CSF (главный регулятор развития нейтрофилов), IL-17 и IL-23, основного фактора хемотаксиса IL-8 (рецепторы CXCR1 и CXCR2), а также хемокина SDF-1, определяющего связь нейтрофилов с тканями (соответствующий ему рецептор известен как CXCR4)[17].
У человека выделяют пять групп антигенов нейтрофилов, которые обозначают HNA 1—5. Три антигена группы HNA-1 (a, b, c) располагаются на Fc-рецепторе CD16. Единственный антиген группы HNA-2, HNA-2a, входит в состав поверхностного гликопротеина CD177. Антигены группы HNA-3, a и b, входят в состав белка SLC44A2[англ.]. Группы HNA-4 и HNA-5 содержат по два антигена, a и b, которые располагаются на интегрине β2[18].
Морфология ядра нейтрофилов


Подобно базофилам и эозинофилам, зрелые нейтрофилы имеют дольчатое клеточное ядро из 2—5 сегментов, соединённых тонкими перемычками. У некоторых животных ядро зрелых нейтрофилов не имеет отчётливой сегментации[14]. По мере созревания нейтрофила ядрышко исчезает (исчезновение ядрышка происходит лишь у некоторых типов дифференцированных клеток, имеющих ядро). Центр ядра составляет эухроматин, а гетерохроматин сосредоточен на периферии. У женщин некоторые нейтрофилы несут инактивированную X-хромосому в виде тельца Барра, которое выглядит как придаток в виде барабанной палочки у одного из сегментов ядра[19][20]. Поскольку промоторы многих генов в ядре нейтрофила недоступны для дифференцировочных факторов, экспрессия генов и возможность к биосинтезу макромолекул у нейтрофилов ограничены. Тем не менее нейтрофилы всё же сохраняют способность к биосинтезу, хотя и ограниченному. У незрелых палочкоядерных нейтрофилов ядро не подразделено на сегменты, хотя содержит уплотнённый хроматин[21].
Аппарат Гольджи, как правило, слабо развит, митохондрии и рибосомы редки, шероховатый эндоплазматический ретикулум (ЭПР) отсутствует. В цитоплазме имеется от 50 до 200 гранул, из которых приблизительно 10—20 % (по некоторым оценкам, до трети) составляют азурофильные гранулы[22], оставшиеся 80—90 % приходится на специфические гранулы. Азурофильные гранулы окружены одинарной мембраной, они более крупные, чем специфические, и имеют диаметр около 0,4 мкм, окрашиваются в фиолетово-красный цвет. Они представляют собой первичные лизосомы, содержат электронно-плотную сердцевину и ферменты, активные при низких значениях pH: кислую фосфатазу, β-глюкуронидазу[англ.], миелопероксидазу, которая продуцирует из перекиси водорода молекулярный кислород, имеющий антибактериальные свойства, α-фукозидазу[англ.], 5’-нуклеотидазу[англ.], арилсульфатазу[англ.], β-галактозидазу, α-маннозидазу[англ.], N-ацетилглюкозаминидазу[англ.], лизоцим, нейтральные протеазы (катепсин G, эластаза, коллагеназа, азуроцидин[англ.]). Кроме того, азурофильные гранулы содержат антимикробные пептиды из числа дефензинов[20], кателицидинов[англ.], гранулофизин, лактоферрин, кислые гликозаминогликаны. Поскольку в ходе дифференцировки нейтрофилов в костном мозге азурофильные гранулы появляются раньше специфических, их также называют первичными гранулами[23]. Молекулярные маркеры[англ.] азурофильных гранул — миелопероксидаза и мембранная молекула CD63[24].
Специфические[англ.], или вторичные гранулы — более мелкие (диаметр около 20 мкм), светлые, многочисленные. Они электронно-прозрачны, но иногда содержат кристаллоид. В специфических гранулах содержатся белки с бактериостатическими[англ.] и бактерицидными свойствами — лизоцим, щелочную фосфатазу, лактоферрин[23], а также белок BPI[англ.], связывающий витамин B12. В специфических гранулах в больших количествах содержится фермент NADPH-оксидаза, которая катализирует образование активных форм кислорода, выступающих в роли главных бактерицидных факторов фагоцитов. Кроме того, в состав специфических гранул входят разнообразные ферменты, разрушающие внеклеточный матрикс, такие как коллагеназы[20]. Молекулярными маркерами специфических гранул являются лактоферрин и мембранная молекула CD66[25].
Помимо первичных и вторичных гранул в нейтрофилах присутствуют третичные желатиназные и секреторные гранулы, или везикулы. Желатиназные гранулы содержат желатиназу B[англ.], также известную как матриксная металлопротеиназа 9 (MMP9). Они мельче специфических гранул и легче подвергаются экзоцитозу. В мембранах и специфических, и желатиназных гранул имеется флавоцитохром b558, который входит в состав NADPH-оксидазы. Секреторные везикулы содержат щелочную фосфатазу[26] и несут на своей поверхности Fc-рецептор CD16, рецепторы комплемента (в том числе CD35), а также интегрины CD11b/CD18, CD11c/CD18 и молекулы CD15 и CD14. После того как в ходе экзоцитоза мембрана секреторной везикулы встроится в клеточную мембрану нейтрофила, перечисленные рецепторы могут быть моментально использованы клеткой, что может резко изменить её фенотип[27].
В таблице ниже перечислены основные компоненты мембран и матрикса гранул нейтрофилов[27][26].
| Компартмент гранулы | Азурофильные (первичные) гранулы | Специфические (вторичные) гранулы | Желатиназные (третичные) гранулы | Секреторные гранулы |
|---|---|---|---|---|
| Мембрана | CD63, CD68 | CD15, CD66, CD67, FPR[англ.], CD120, b558 | CD11b/CD18, FPR, b558 | CD11b/CD18, CD13, CD14, CD16, FPR, CD35 |
| Матрикс | Металлопротеиназы[англ.], катепсин G, эластаза, азуроцидин, лизоцим, BPI, α-дефензины | Лизоцим, кателицидин, BPI, липокалин 2[англ.], лактоферрин, фосфолипаза A2, коллагеназа | Желатиназа B | Азуроцидин, альбумин, щелочная фосфатаза |
Нейтрофилы содержат гликоген, и основную роль в получении энергии у нейтрофилов играет гликолиз. Цикл Кребса и окислительное фосфорилирование вносят меньший вклад в энергообеспечение нейтрофила, о чём свидетельствует небольшое количество митохондрий у данного типа клеток. Способность нейтрофилов выживать в анаэробных условиях очень важна для их функционирования, поскольку позволяет им убивать и фагоцитировать бактерии даже в бедных кислородом тканях, например, в повреждённых или некротизированных тканях[20].
Развитие

Нейтрофилы образуются в костном мозге из гемопоэтических стволовых клеток. Гемопоэтическая стволовая клетка даёт начало мультипотентной[англ.] клетке — родоначальнице гранулоцитарного, эритроцитарного, моноцитарного и мегакариоцитарного рядов гемопоэза, которая, в свою очередь, дает начало олигопотентной клетке-предшественнице моноцитов и нейтрофилов. От неё происходит унипотентная клетка-предшественница нейтрофилов, дающая начало миелобластам. Композиция её поверхностных маркеров описывается как CD34+CD33+, а также рецепторы для GM-CSF, G-CSF, IL-1, IL-3, IL-6, IL-11[англ.], IL-12[28]. Далее последовательность клеток-предшественников выглядит следующим образом: миелобласт → промиелоцит → нейтрофильный миелоцит → метамиелоцит → палочкоядерный нейтрофил → сегментоядерный нейтрофил. Первичные гранулы появляются на стадии промиелоцитов, вторичные — на стадии миелоцитов. До стадии метамиелоцитов предшественники делятся митозом, а метамиелоциты и последующие стадии лишены способности к делению. На стадии метамиелоцитов увеличивается количество специфических гранул в цитоплазме, дальнейшее созревание ядра приводит появлению палочкоядерных нейтрофилов. Сегментация ядра палочкоядерных нейтрофилов приводит к их преобразованию в зрелые сегментоядерные нейтрофилы. Полный период развития нейтрофилов у человека составляет около 14 суток, из которых 7,5 суток приходятся на пролиферацию, а 6,5 — на постмитотическую дифференцировку[29]. У взрослого человека за сутки из костного мозга в кровь выходит от 5 × 1010 до 1011 зрелых нейтрофилов[2].
Срок жизни неактивированного нейтрофила в кровотоке составляет, по разным оценкам, от 5 до 135 часов[30][31]. При активации нейтрофилы приобретают способность протискиваться через эндотелий сосудов и мигрируют в ткани, где они живут от одного до двух дней. Около 30 % нейтрофилов, покидающих кровоток, мигрируют в костный мозг и печень, 20 % направляются в лёгкие, 15 % — в селезёнку. Главные факторы хемотаксиса нейтрофилов, направляющие их перемещение в ткани, — лейкотриен B4 и IL-8. В процессе миграции нейтрофилов участвуют молекулы адгезии, а именно, β2-интегрины, P- и E-селектины, а также секретируемый нейтрофилами фермент эластаза[англ.]. Нейтрофилы гораздо многочисленнее долгоживущих макрофагов, и патоген, проникнувший в организм, прежде всего сталкивается именно с нейтрофилами. Через 3—5 суток пребывания в тканях нейтрофилы подвергаются апоптозу и поглощаются резидентными макрофагами. Некоторые исследователи считают, что короткий срок жизни нейтрофилов является эволюционной адаптацией. Со смертью фагоцита погибают патогены, сохраняющиеся в нём, что защищает организм. Кроме того, ввиду высокой токсичности веществ, выделяемых нейтрофилами для борьбы с инфекцией, по отношению к тканям организма, быстрая гибель нейтрофилов обеспечивает действие противомикробных веществ только в очаге воспаления и защищает остальные ткани организма[32][20]. Существуют сведения, указывающие на возможность перехода нейтрофилов в тканях в долгоживущую форму и даже в макрофаги[21].
Функции

Воспалённые или повреждённые участки соединительной ткани требуют немедленной миграции разнообразных лейкоцитов, в том числе нейтрофилов, в очаг повреждения для удаления патогенных микроорганизмов и восстановления ткани. Наиболее хорошо процесс миграции в ткани изучен для нейтрофилов, которые прибывают в очаг воспаления первыми, существенно быстрее моноцитов, и способны развивать защитные метаболические реакции (в частности, «окислительный взрыв[англ.]», сопровождающийся продукцией активных форм кислорода) в течение секунд. Активация нейтрофилов сопровождается в первую очередь высвобождением содержимого секреторных гранул. При развитии локального воспаления макрофаги, активированные бактериями или повреждениями ткани, выделяют провоспалительные цитокины[англ.], такие как IL-1 или фактор некроза опухоли α (TNF-α). Также к факторам хемотаксиса нейтрофилов относят компоненты комплемента, вещества, выделяемые тучными клетками, иммунные комплексы[англ.], эндотоксины и бактериальные пептиды, а также наполненные лизосомы, выходящие в ткань при распаде погибших нейтрофилов и макрофагов[33]. Под действием провоспалительных цитокинов в эндотелиальных клетках, выстилающих ближайшие к очагу воспаления посткапиллярные венулы, увеличивается количество селектинов на поверхности, обращённой в просвет сосуда. Нейтрофилы, циркулирующие в посткапиллярных венулах и имеющие подходящий набор поверхностных гликопротеинов, связываются с селектинами на поверхности эндотелиальных клеток. На данном этапе связывание с эндотелием непрочное, и нейтрофилы продолжают «катиться» по поверхности эндотелия. Провоспалительные цитокины запускают экспрессию интегринов в катящихся нейтрофилах и лигандов интегринов, известных как ICAM-1, на поверхностях эндотелиальных клеток. В то же время межклеточные контакты между клетками эндотелия ослабляются, и эндотелий становится более проницаемым. За счёт интегринов нейтрофилы прочно связываются с эндотелием и прекращают качение, и в конечном счёте приступают к прохождению через эндотелий с помощью псевдоподий (этот процесс известен как диапедез). Проникновение нейтрофилов через эндотелий облегчается благодаря длинному и сегментированному ядру. Прохождение нейтрофилов через эндотелий и дальнейшая миграция к очагу воспаления стимулируются хемокинами[34]. Базальные мембраны нейтрофилы преодолевают благодаря выделению содержимого желатиназных гранул[35].
Координированное перемещение нейтрофилов в очаг острого воспаления называют роением нейтрофилов[англ.][36]. Масштаб роения и его длительность определяются многими факторами, среди которых размер повреждённой области ткани и наличие патогенов[37]. Явление роения нейтрофилов было изучено преимущественно у мышей на ткани уха[38] и у рыбок данио-рерио[39].

Активированные нейтрофилы при встрече с микроорганизмами поглощают их с помощью псевдоподий, и микроорганизм оказывается внутри фагосомы. В течение нескольких секунд после активации нейтрофила изменяется его мембранный потенциал, в клетку входят ионы натрия и кальция, изменяется текучесть мембраны[англ.][33]. Примерно через 30 секунд после поглощения частицы специфические гранулы сливаются с фагосомой, изливая в неё своё содержимое, после чего фагосома дополнительно закисляется с помощью мембранных протонных помп. С закисленными фагосомами сливаются азурофильные гранулы (примерно через 1—3 минуты после поглощения частицы). В ходе фагоцитоза в нейтрофиле образуются активные формы кислорода, такие как супероксид-анион и перекись водорода, и другие компоненты гранул с бактерицидными свойствами. Активные формы кислорода используются для уничтожения бактериальной клетки, наряду с содержимым специфических и азурофильных гранул. Белок лактоферрин, входящий в состав специфических гранул, не убивает бактерию напрямую, а прочно связывает ионы железа, делая их недоступными для бактерии и, в результате, приводя к её смерти. Содержимое гранул (в частности, азурофильных) может высвобождаться в ходе дегрануляции[англ.], после которой восстановления гранул не происходит. Комбинация описанных механизмов способна убить практически любую бактерию, и погибшие бактериальные клетки впоследствии расщепляются лизосомными ферментами. Наиболее чувствительны к компонентам гранул нейтрофилов дрожжи (Candida) и бактерии из числа стрептококков и стафилококков. Нейтрофилы, погибшие апоптозом, бактерии, полуразрушенные останки клеток и тканевая жидкость формируют густой гной белого или жёлтого цвета[40][35].

Ещё одна форма противомикробной активности нейтрофилов заключается в особом типе программируемой клеточной гибели, характерном для нейтрофилов, — нетозе[41]. При нетозе погибающий нейтрофил выбрасывает свою ДНК наружу в виде так называемых внеклеточных ловушек нейтрофилов (от англ. neutrophil extracellular traps, NETs). Внеклеточные ловушки нейтрофилов состоят из хроматина и сериновых протеаз и способны иммобилизовать и убивать микробные клетки[42]. Таким образом, внеклеточные ловушки нейтрофилов также противодействуют распространению клеток патогенов по тканям. При сепсисе массовый нетоз происходит непосредственно в кровеносных сосудах[43]. Образование внеклеточных ловушек нейтрофилов может вносить вклад в развитие ряда воспалительных заболеваний, таких как преэклампсия[44], а их образование в кровеносных сосудов может приводить к формированию тромбов, в том числе в коронарных артериях[45][46]. В 2018 году было показано, что один и тот же нейтрофил может подвергаться и апоптозу, и нетозу одновременно; этот вид программируемой клеточной гибели нейтрофилов получил название апонетоз[47].
Многие исследователи выделяют две функционально различные субпопуляции нейтрофилов на основании различных интенсивности образования активных форм кислорода, проницаемости мембраны, активности ферментов гранул и способности к инактивации. Нейтрофилы с повышенной проницаемостью мембран называют нейтрофилами-киллерами. Нейтрофилы-киллеры интенсивно генерируют активные формы кислорода и инактивируются после взаимодействия с субстратом. Нейтрофилы второй субпопуляции образуют активные формы кислорода менее интенсивно, не прикрепляются к субстрату и не инактивируются[48][49][50][51].
Активность нейтрофилов зависит от возраста организма человека. У новорождённого нет адекватной продукции нейтрофилов, а в старческом возрасте способность нейтрофилов к фагоцитозу ограничена[52].
Нейтрофилы экспрессируют и продуцируют широкий спектр цитокинов, среди которых хемокины, колониестимулирующие факторы[англ.], провоспалительные цитокины (IL-1α, IL-1β, IL-6, IL-7, IL-18, MIF[англ.] и другие), иммунорегуляторные цитокины (IL-12, IL-21[англ.], IL-23[англ.], IL-27[англ.], TSLP[англ.] и другие), противовспалительные цитокины (IL-1ra, TGFβ1, TGFβ2[англ.]), факторы ангиогенеза и фиброгенеза (VEGF, BV8, HBEGF[англ.], FGF2[англ.], TGFα[англ.], HGF[англ.], ангиопоэтин), цитокины суперсемейства фактора некроза опухоли (TNF) и некоторые другие цитокины, такие как PBEF, амфирегулин[англ.], мидкин, онкостатин M[англ.], активин A[англ.], эндотелин. За счёт выделения разнообразных цитокинов нейтрофилы могут быть вовлечены в процессы, не связанные с иммунной защитой, такие как гемопоэз, ангиогенез и заживление ран. Кроме того, нейтрофилы могут участвовать в развитии некоторых аутоиммунных и злокачественных заболеваний[53].
Иммуносупрессорные популяции нейтрофилов задействованы в поддержании иммунологической толерантности при беременности. В частности, иммуносупрессорные нейтрофилы низкой плотности могут подавлять пролиферацию CD4+ и CD8+ T-лимфоцитов, подвергая их воздействию активных форм кислорода. Кроме того, они подавляют цитотоксическую активность естественных киллеров и экспрессию Toll-подобных рецепторов на моноцитах. Нейтрофилы, располагающиеся в децидуальной оболочке[англ.] во время второго триместра беременности, стимулируют в ней дополнительный ангиогенез[54].
Клиническое значение

Состояние, при котором количество нейтрофилов снижено, называется нейтропения. Нейтропения может быть наследственной или приобретённой, как при некоторых видах апластической анемии или лейкозов. Нейтропения может развиться вследствие приёма определённых препаратов, в частности, противораковой химиотерапии. Пациенты с нейтропенией отличаются повышенной уязвимостью к инфекционным заболеваниям бактериальной и грибковой природы. Чаще всего пациенты с нейтропенией страдают от таких заболеваний, как воспаление подкожной жировой клетчатки, фурункулёз, пневмония и септицемия[55]. Степень нейтропении определяют с помощью параметра, известного как абсолютное количество нейтрофилов[англ.] (англ. Absolute neutrophil count, ANC). Состояние с ANC < 1500 клеток / мм³ считается нейтропенией, а с ANC < 500 клеток / мм³ — тяжёлой нейтропенией[56].

Состояние, противоположное нейтропении, при котором количество нейтрофилов в крови повышено, называют нейтрофилией. Наиболее частой причиной нейтрофилии являются бактериальные инфекции, особенно сопровождающиеся гнойным воспалением[57]. Повышение уровня нейтрофилов происходит при любом остром воспалении, поэтому нейтрофилия может появиться в результате инфаркта миокарда или ожога[57], а также аппендицита и спленэктомии. Уровень нейтрофилов может повышаться при некоторых злокачественных заболеваниях крови, таких как хронический миелоидный лейкоз, при котором происходит неконтролируемая пролиферация лейкоцитов[58].
Нейтрофилы, будучи иммунными клетками, принимают участие во взаимодействии с клетками опухолей. Нейтрофилы, инфильтрованные в опухоль, имеют повышенную, по сравнению с нейтрофилами в обычных тканях, продолжительность жизни. Нейтрофилы могут играть ведущую роль в онкогенезе за счёт взаимодействия с другими иммунными клетками, реагирующими на появление и рост опухоли. Нейтрофилы могут стимулировать пролиферацию опухолевых клеток и ангиогенез в опухоли, активировать метастазирование и подавлять иммунный ответ на злокачественные клетки. В то же время нейтрофилы могут обладать и противоопухолевой активностью[2].
Известно несколько наследственных расстройств, связанных с дисфункцией нейтрофилов. В некоторых случаях подвижность нейтрофилов снижена за счёт нарушений в полимеризации актина, а при недостаточной экспрессии гена, кодирующего NADPH-оксидазу, нейтрофилы теряют способность вырабатывать перекись водорода и гипохлорит, что уменьшает их противомикробные свойства. Дети, страдающие от таких наследственных нарушений, сильнее подвержены бактериальным инфекциям[34]. Наследственная миелопероксидазная недостаточность[англ.], связанная с мутациями в гене, кодирующем миелопероксидазу, рассматривается как первичный иммунодефицит[59].
Недостаточная или избыточная активность ряда белков, входящих в состав гранул нейтрофилов, приводит к болезненным состояниям. При наследственной болезни, известной как альфа-1 антитрипсиновая недостаточность[англ.], эластаза в составе гранул нейтрофилов недостаточно ингибируется альфа-1 антитрипсином[англ.], что приводит к сильнейшим повреждениям тканей при воспалительных процессах, в частности, при хронической обструктивной болезни лёгких. Повышенная активность эластазы нейтрофилов может привести к разрушению лёгочного барьера и острому респираторному дистресс-синдрому[60]. Эластаза нейтрофилов влияет на активность макрофагов, разрушая их Toll-подобные рецепторы и подавляя синтез цитокинов через ингибирование перемещения в ядро транскрипционного фактора NF-κB[61]. При периодической болезни мутации в гене MEFV[англ.], кодирующем белок пирин, экспрессирующийся преимущественно в нейтрофилах, могут привести к хроническому острому воспалению, что выражается в лихорадке, артралгии, перитоните и, в конце концов, может стать причиной амилоидоза[62]. Имеются данные о связи сниженной активности нейтрофилов, связанной с нарушениями активности миелопероксидазы и дегрануляции, с гипергликемией[63].
Нейтрофилы связаны с развитием многих хронических воспалительных заболеваний, в том числе аутоиммунных. Они поддерживают хроническое воспаление при системной красной волчанке, псориазе, хроническом гранулёматозе[англ.], АНЦА-ассоциированном системном васкулите, связанном с образованием аутоантител к компонентам цитоплазмы нейтрофилов, ревматоидном артрите, идиопатическом юношеском артрите и PAPA-синдроме[англ.] (от англ. pyogenic arthritis-pyoderma gangrenosum-acne syndrome)[54].
История изучения

Впервые нейтрофилы описал французский исследователь Жан-Батист Сенак, который в 1749 году обнаружил белые клетки в гное. В 1843 году английский врач Уильям Аддисон[англ.] заметил, что белые клетки в составе гноя происходят от лейкоцитов, которые просочились через кровеносные сосуды. В 1845 году английский врач и физиолог Джон Хьюз Беннет[англ.] сообщил, что лейкоз обусловлен накоплением белого гноя в кровеносных сосудах, но уже в 1847 году это представление было опровергнуто Рудольфом Вирховым, который показал, что патологические клетки при лейкозе происходят не из гноя, а из самой крови. Первое точное морфологическое описание различных гранулоцитов, в том числе и нейтрофилов, было выполнено Максом Шульце в 1865 году. Кроме морфологических описаний гранулоцитов, Шульце описал и фагоцитоз[64]. Кроме того, в 1876 году фагоцитоз был отмечен Уильямом Ослером[65]
Фагоцитоз был детально изучен и назван Ильёй Ильичом Мечниковым в 1882 году, когда им же была установлена роль фагоцитов в защите от бактерий[66]. В 1879—1880 годах Пауль Эрлих опубликовал несколько работ, в которых детально описал морфологию белых клеток крови. Окрашивание кислыми и щелочными красителями позволило ему выявить соответственно эозинофилы и базофилы, а также тучные клетки. Впоследствии с помощью окрашивания нейтральными красителями он визуализировал сегментоядерные нейтрофилы и их гранулы (за что нейтрофилы и получили своё название). Первоначально Эрлих назвал нейтрофилы «клетками с полиморфными ядрами». Также он описал некоторые аспекты появления, функционирования и дальнейшей судьбы эозинофилов и нейтрофилов[64]. В 1908 году Пауль Эрлих и Илья Мечников разделили Нобелевскую премию по физиологии и медицине[67].
Примечания
- ↑ 1 2 3 Ярилин, 2010, с. 52.
- ↑ 1 2 3 Andryukov B. G., Bogdanova V. D., Lyapun I. N. Phenotypic heterogeneity of neutrophils: new antimicrobic charasteristics and diagnostic technologies (англ.) // Russian journal of hematology and transfusiology. — 2019. — 4 October (vol. 64, no. 2). — P. 211—221. — ISSN 2411-3042. — doi:10.35754/0234-5730-2019-64-2-211-221.
- ↑ Actor Jeffrey K. Cells and Organs of the Immune System (англ.) // Elsevier's Integrated Review Immunology and Microbiology. — 2012. — P. 7—16. — ISBN 9780323074476. — doi:10.1016/B978-0-323-07447-6.00002-8.
- ↑ Ermert D., Niemiec M. J., Röhm M., Glenthøj A., Borregaard N., Urban C. F. Candida albicans escapes from mouse neutrophils. (англ.) // Journal Of Leukocyte Biology. — 2013. — August (vol. 94, no. 2). — P. 223—236. — doi:10.1189/jlb.0213063. — PMID 23650619.
- ↑ Хаитов, 2019, с. 73.
- ↑ Witko-Sarsat V., Rieu P., Descamps-Latscha B., Lesavre P., Halbwachs-Mecarelli L. Neutrophils: molecules, functions and pathophysiological aspects. (англ.) // Laboratory Investigation; A Journal Of Technical Methods And Pathology. — 2000. — May (vol. 80, no. 5). — P. 617—653. — doi:10.1038/labinvest.3780067. — PMID 10830774.
- ↑ Nathan C. Neutrophils and immunity: challenges and opportunities. (англ.) // Nature Reviews. Immunology. — 2006. — March (vol. 6, no. 3). — P. 173—182. — doi:10.1038/nri1785. — PMID 16498448.
- ↑ Jacobs L., Nawrot T. S., de Geus B., Meeusen R., Degraeuwe B., Bernard A., Sughis M., Nemery B., Panis L. I. Subclinical responses in healthy cyclists briefly exposed to traffic-related air pollution: an intervention study. (англ.) // Environmental Health : A Global Access Science Source. — 2010. — 25 October (vol. 9). — P. 64—64. — doi:10.1186/1476-069X-9-64. — PMID 20973949.
- ↑ Waugh D. J., Wilson C. The interleukin-8 pathway in cancer. (англ.) // Clinical Cancer Research : An Official Journal Of The American Association For Cancer Research. — 2008. — 1 November (vol. 14, no. 21). — P. 6735—6741. — doi:10.1158/1078-0432.CCR-07-4843. — PMID 18980965.
- ↑ De Larco J. E., Wuertz B. R., Furcht L. T. The potential role of neutrophils in promoting the metastatic phenotype of tumors releasing interleukin-8. (англ.) // Clinical Cancer Research : An Official Journal Of The American Association For Cancer Research. — 2004. — 1 August (vol. 10, no. 15). — P. 4895—4900. — doi:10.1158/1078-0432.CCR-03-0760. — PMID 15297389.
- ↑ Yoo S. K., Starnes T. W., Deng Q., Huttenlocher A. Lyn is a redox sensor that mediates leukocyte wound attraction in vivo. (англ.) // Nature. — 2011. — 20 November (vol. 480, no. 7375). — P. 109—112. — doi:10.1038/nature10632. — PMID 22101434.
- ↑ Barer M.R. The natural history of infection (англ.) // Medical Microbiology. — 2012. — P. 168—173. — ISBN 9780702040894. — doi:10.1016/b978-0-7020-4089-4.00029-9.
- ↑ Reich D., Nalls M. A., Kao W. H., Akylbekova E. L., Tandon A., Patterson N., Mullikin J., Hsueh W. C., Cheng C. Y., Coresh J., Boerwinkle E., Li M., Waliszewska A., Neubauer J., Li R., Leak T. S., Ekunwe L., Files J. C., Hardy C. L., Zmuda J. M., Taylor H. A., Ziv E., Harris T. B., Wilson J. G. Reduced neutrophil count in people of African descent is due to a regulatory variant in the Duffy antigen receptor for chemokines gene. (англ.) // PLoS Genetics. — 2009. — January (vol. 5, no. 1). — P. e1000360—1000360. — doi:10.1371/journal.pgen.1000360. — PMID 19180233.
- ↑ 1 2 Eclinpath: normal leukocytes. Дата обращения: 20 сентября 2020. Архивировано 1 октября 2020 года.
- ↑ Niemiec M. J., De Samber B., Garrevoet J., Vergucht E., Vekemans B., De Rycke R., Björn E., Sandblad L., Wellenreuther G., Falkenberg G., Cloetens P., Vincze L., Urban C. F. Trace element landscape of resting and activated human neutrophils on the sub-micrometer level. (англ.) // Metallomics : Integrated Biometal Science. — 2015. — June (vol. 7, no. 6). — P. 996—1010. — doi:10.1039/c4mt00346b. — PMID 25832493.
- ↑ Edwards Steven W. Biochemistry and physiology of the neutrophil. — Cambridge University Press, 1994. — P. 6. — ISBN 978-0-521-41698-6.
- ↑ Ярилин, 2010, с. 53.
- ↑ Chu H. T., Lin H., Tsao T. T., Chang C. F., Hsiao W. W., Yeh T. J., Chang C. M., Liu Y. W., Wang T. Y., Yang K. C., Chen T. J., Chen J. C., Chen K. C., Kao C. Y. Genotyping of human neutrophil antigens (HNA) from whole genome sequencing data. (англ.) // BMC Medical Genomics. — 2013. — 12 September (vol. 6). — P. 31—31. — doi:10.1186/1755-8794-6-31. — PMID 24028078.
- ↑ Афанасьев и др., 2004, с. 176.
- ↑ 1 2 3 4 5 Mescher, 2016, p. 243.
- ↑ 1 2 Ярилин, 2010, с. 52—53.
- ↑ Zucker-Franklin D, Greaves MF, Grossi CE, Marmont AM. Neutrophils // Atlas of Blood Cells: Function and Pathology. — 2nd. — Philadelphia : Lea & Ferbiger, 1988. — Vol. 1. — P. 168—170. — ISBN 978-0-8121-1094-4.
- ↑ 1 2 Афанасьев и др., 2004, с. 177—178.
- ↑ Ярилин, 2010, с. 54—55.
- ↑ Ярилин, 2010, с. 54.
- ↑ 1 2 Ярилин, 2010, с. 55.
- ↑ 1 2 Хаитов, 2019, с. 75.
- ↑ Руководство по гематологии, 2002, с. 89.
- ↑ Афанасьев и др., 2004, с. 201—204.
- ↑ Tak T., Tesselaar K., Pillay J., Borghans J. A., Koenderman L. What's your age again? Determination of human neutrophil half-lives revisited. (англ.) // Journal Of Leukocyte Biology. — 2013. — October (vol. 94, no. 4). — P. 595—601. — doi:10.1189/jlb.1112571. — PMID 23625199.
- ↑ Pillay J., den Braber I., Vrisekoop N., Kwast L. M., de Boer R. J., Borghans J. A., Tesselaar K., Koenderman L. In vivo labeling with 2H2O reveals a human neutrophil lifespan of 5.4 days. (англ.) // Blood. — 2010. — 29 July (vol. 116, no. 4). — P. 625—627. — doi:10.1182/blood-2010-01-259028. — PMID 20410504.
- ↑ Wheater Paul R., Stevens Alan. Wheater's basic histopathology: a colour atlas and text. — Edinburgh : Churchill Livingstone, 2002. — ISBN 978-0-443-07001-3.
- ↑ 1 2 Руководство по гематологии, 2002, с. 91.
- ↑ 1 2 Mescher, 2016, p. 244.
- ↑ 1 2 Ярилин, 2010, с. 56.
- ↑ Tan S. Y., Weninger W. Neutrophil migration in inflammation: intercellular signal relay and crosstalk. (англ.) // Current Opinion In Immunology. — 2017. — February (vol. 44). — P. 34—42. — doi:10.1016/j.coi.2016.11.002. — PMID 27951434.
- ↑ Lämmermann T. In the eye of the neutrophil swarm-navigation signals that bring neutrophils together in inflamed and infected tissues. (англ.) // Journal Of Leukocyte Biology. — 2016. — July (vol. 100, no. 1). — P. 55—63. — doi:10.1189/jlb.1MR0915-403. — PMID 26416718.
- ↑ Chtanova T., Schaeffer M., Han S. J., van Dooren G. G., Nollmann M., Herzmark P., Chan S. W., Satija H., Camfield K., Aaron H., Striepen B., Robey E. A. Dynamics of neutrophil migration in lymph nodes during infection. (англ.) // Immunity. — 2008. — 19 September (vol. 29, no. 3). — P. 487—496. — doi:10.1016/j.immuni.2008.07.012. — PMID 18718768.
- ↑ Henry K. M., Loynes C. A., Whyte M. K., Renshaw S. A. Zebrafish as a model for the study of neutrophil biology. (англ.) // Journal Of Leukocyte Biology. — 2013. — October (vol. 94, no. 4). — P. 633—642. — doi:10.1189/jlb.1112594. — PMID 23463724.
- ↑ Mescher, 2016, p. 243—244.
- ↑ Brinkmann V., Reichard U., Goosmann C., Fauler B., Uhlemann Y., Weiss D. S., Weinrauch Y., Zychlinsky A. Neutrophil extracellular traps kill bacteria. (англ.) // Science (New York, N.Y.). — 2004. — Vol. 303, no. 5663. — P. 1532—1535. — doi:10.1126/science.1092385. — PMID 15001782.
- ↑ Urban C. F., Ermert D., Schmid M., Abu-Abed U., Goosmann C., Nacken W., Brinkmann V., Jungblut P. R., Zychlinsky A. Neutrophil extracellular traps contain calprotectin, a cytosolic protein complex involved in host defense against Candida albicans. (англ.) // PLoS Pathogens. — 2009. — October (vol. 5, no. 10). — P. e1000639—1000639. — doi:10.1371/journal.ppat.1000639. — PMID 19876394.
- ↑ Clark S. R., Ma A. C., Tavener S. A., McDonald B., Goodarzi Z., Kelly M. M., Patel K. D., Chakrabarti S., McAvoy E., Sinclair G. D., Keys E. M., Allen-Vercoe E., Devinney R., Doig C. J., Green F. H., Kubes P. Platelet TLR4 activates neutrophil extracellular traps to ensnare bacteria in septic blood. (англ.) // Nature Medicine. — 2007. — April (vol. 13, no. 4). — P. 463—469. — doi:10.1038/nm1565. — PMID 17384648.
- ↑ Gupta A. K., Hasler P., Holzgreve W., Hahn S. Neutrophil NETs: a novel contributor to preeclampsia-associated placental hypoxia? (англ.) // Seminars In Immunopathology. — 2007. — June (vol. 29, no. 2). — P. 163—167. — doi:10.1007/s00281-007-0073-4. — PMID 17621701.
- ↑ Hoyer F. F., Nahrendorf M. Neutrophil contributions to ischaemic heart disease. (англ.) // European Heart Journal. — 2017. — 14 February (vol. 38, no. 7). — P. 465—472. — doi:10.1093/eurheartj/ehx017. — PMID 28363210.
- ↑ Mangold A., Alias S., Scherz T., Hofbauer T., Jakowitsch J., Panzenböck A., Simon D., Laimer D., Bangert C., Kammerlander A., Mascherbauer J., Winter M. P., Distelmaier K., Adlbrecht C., Preissner K. T., Lang I. M. Coronary neutrophil extracellular trap burden and deoxyribonuclease activity in ST-elevation acute coronary syndrome are predictors of ST-segment resolution and infarct size. (англ.) // Circulation Research. — 2015. — 27 March (vol. 116, no. 7). — P. 1182—1192. — doi:10.1161/CIRCRESAHA.116.304944. — PMID 25547404.
- ↑ Azzouz Dhia, Palaniyar Nades. ApoNETosis: discovery of a novel form of neutrophil death with concomitant apoptosis and NETosis (англ.) // Cell Death & Disease. — 2018. — August (vol. 9, no. 8). — ISSN 2041-4889. — doi:10.1038/s41419-018-0846-9.
- ↑ Gerasimov I. G., Ignatov D. Iu. Functional heterogenicity of human blood neutrophils: generation of oxygen active species. // Tsitologiia. — 2001. — Т. 43, № 5. — С. 432—436. — PMID 11517658.
- ↑ Gerasimov I. G., Ignatov D. Iu. Neutrophil activation in vitro. // Tsitologiia. — 2004. — Т. 46, № 2. — С. 155—158. — PMID 15174354.
- ↑ Gerasimov I. G., Ignatov D. Iu., Kotel'nitskiĭ M. A. Nitroblue tetrazolium reduction by human blood neutrophils. I. The influence of pH. // Tsitologiia. — 2005. — Т. 47, № 6. — С. 549—553. — PMID 16708848.
- ↑ Gerasimov I. G., Ignatov D. Iu. Nitroblue tetrazolium reduction by human blood neutrophils. II. The influence of sodium and potassium ions. // Tsitologiia. — 2005. — Т. 47, № 6. — С. 554—558. — PMID 16708849.
- ↑ Руководство по гематологии, 2002, с. 90.
- ↑ Tecchio C., Micheletti A., Cassatella M. A. Neutrophil-derived cytokines: facts beyond expression. (англ.) // Frontiers In Immunology. — 2014. — Vol. 5. — P. 508—508. — doi:10.3389/fimmu.2014.00508. — PMID 25374568.
- ↑ 1 2 Silvestre-Roig C., Fridlender Z. G., Glogauer M., Scapini P. Neutrophil Diversity in Health and Disease. (англ.) // Trends In Immunology. — 2019. — July (vol. 40, no. 7). — P. 565—583. — doi:10.1016/j.it.2019.04.012. — PMID 31160207.
- ↑ Справочник MSD: Нейтропения. Дата обращения: 20 сентября 2020. Архивировано 25 ноября 2020 года.
- ↑ Al-Gwaiz L. A., Babay H. H. The diagnostic value of absolute neutrophil count, band count and morphologic changes of neutrophils in predicting bacterial infections. (англ.) // Medical Principles And Practice : International Journal Of The Kuwait University, Health Science Centre. — 2007. — Vol. 16, no. 5. — P. 344—347. — doi:10.1159/000104806. — PMID 17709921.
- ↑ 1 2 Mitchell, Richard Sheppard, Kumar, Vinay, Abbas, Abul K., Fausto, Nelson. Robbins Basic Pathology. — Philadelphia : Saunders. — ISBN 1-4160-2973-7.
- ↑ bloodandcancerclinic. Дата обращения: 10 апреля 2013. Архивировано из оригинала 21 мая 2013 года.
- ↑ OMIM Entry - # 254600 - MYELOPEROXIDASE DEFICIENCY; MPOD. omim.org. Дата обращения: 17 марта 2020. Архивировано 13 марта 2020 года.
- ↑ Kawabata K., Hagio T., Matsuoka S. The role of neutrophil elastase in acute lung injury. (англ.) // European Journal Of Pharmacology. — 2002. — 6 September (vol. 451, no. 1). — P. 1—10. — doi:10.1016/s0014-2999(02)02182-9. — PMID 12223222.
- ↑ Domon H., Nagai K., Maekawa T., Oda M., Yonezawa D., Takeda W., Hiyoshi T., Tamura H., Yamaguchi M., Kawabata S., Terao Y. Neutrophil Elastase Subverts the Immune Response by Cleaving Toll-Like Receptors and Cytokines in Pneumococcal Pneumonia. (англ.) // Frontiers In Immunology. — 2018. — Vol. 9. — P. 732—732. — doi:10.3389/fimmu.2018.00732. — PMID 29922273.
- ↑ Ozen S. Familial mediterranean fever: revisiting an ancient disease. (англ.) // European Journal Of Pediatrics. — 2003. — July (vol. 162, no. 7-8). — P. 449—454. — doi:10.1007/s00431-003-1223-x. — PMID 12751000.
- ↑ Xiu F., Stanojcic M., Diao L., Jeschke M. G. Stress hyperglycemia, insulin treatment, and innate immune cells. (англ.) // International Journal Of Endocrinology. — 2014. — Vol. 2014. — P. 486403—486403. — doi:10.1155/2014/486403. — PMID 24899891.
- ↑ 1 2 Kay A. B. Paul Ehrlich and the Early History of Granulocytes. (англ.) // Microbiology Spectrum. — 2016. — August (vol. 4, no. 4). — doi:10.1128/microbiolspec.MCHD-0032-2016. — PMID 27726791.
- ↑ Ambrose C. T. The Osler slide, a demonstration of phagocytosis from 1876 Reports of phagocytosis before Metchnikoff's 1880 paper. (англ.) // Cellular Immunology. — 2006. — March (vol. 240, no. 1). — P. 1—4. — doi:10.1016/j.cellimm.2006.05.008. — PMID 16876776.
- ↑ Gordon S. Phagocytosis: An Immunobiologic Process. (англ.) // Immunity. — 2016. — 15 March (vol. 44, no. 3). — P. 463—475. — doi:10.1016/j.immuni.2016.02.026. — PMID 26982354.
- ↑ The Nobel Prize in Physiology or Medicine 1908, Paul Erlich - Biography. Дата обращения: 20 сентября 2020. Архивировано 14 августа 2018 года.
Литература
- Афанасьев Ю. И., Кузнецов С. Л., Юрина Н. А., Котовский Е. Ф. и др. Гистология, цитология и эмбриология. — 6-е изд., перераб. и доп.. — М.: Медицина, 2004. — 768 с. — ISBN 5-225-04858-7.
- А. Н. Маянский, А. Н. Галиуллин. Реактивность нейтрофила. — Казань: Изд-во Казан. ун-та, 1984. — 160 с.
- Руководство по гематологии: в 3 т. / Под ред. А. И. Воробьёва. — М.: Ньюдиамед, 2002. — Т. 1. — 280 с. — ISBN 5-88107-038-0.
- Хаитов Р. М. Иммунология: структура и функции иммунной системы. — М.: ГЭОТАР-Медиа, 2019. — 328 с. — ISBN 978-5-9704-4962-2.
- Ярилин А. А. Иммунология. — М.: ГЭОТАР-Медиа, 2010. — 752 с. — ISBN 978-5-9704-1319-7.
- Anthony L. Mescher. Junqueira's Basic Histology. — McGraw-Hill Education, 2016. — ISBN 978-0-07-184270-9.
| Словари и энциклопедии | |
|---|---|
| В библиографических каталогах |