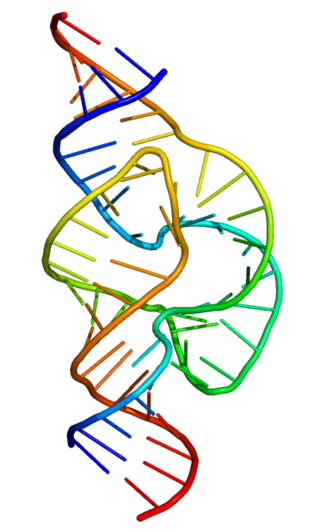
Нуклеиновая кислота
Нуклеи́новая кислота (от лат. nucleus — ядро) — высокомолекулярное органическое соединение, биополимер (полинуклеотид), образованный остатками нуклеотидов. Нуклеиновые кислоты ДНК и РНК присутствуют в клетках всех живых организмов и выполняют важнейшие функции по хранению, передаче и реализации наследственной информации.
История исследования
- В 1847 из экстракта мышц быка было выделено[1] вещество, которое получило название «инозиновая кислота». Это соединение стало первым изученным нуклеотидом. В течение последующих десятилетий были установлены детали его химического строения. В частности, было показано, что инозиновая кислота является рибозид-5'-фосфатом и содержит N-гликозидную связь.
- В 1868 году швейцарским химиком Фридрихом Мишером при изучении некоторых биологических субстанций было открыто неизвестное ранее вещество. Вещество содержало фосфор и не разлагалось под действием протеолитических ферментов. Также оно обладало выраженными кислотными свойствами. Вещество было названо «нуклеином». Соединению была приписана брутто-формула C29H49N9O22P3.
- Уилсон обратил внимание на практическую идентичность химического состава «нуклеина» и открытого незадолго до этого «хроматина» — главного компонента хромосом[2]. Было выдвинуто предположение об особой роли «нуклеина» в передаче наследственной информации.
- В 1889 г. Рихард Альтман ввел термин «нуклеиновая кислота», а также разработал удобный способ получения нуклеиновых кислот, не содержащих белковых примесей.
- Фебус Левин[англ.] и Уолтер Джейкобс[англ.], изучая продукты щелочного гидролиза нуклеиновых кислот, выделили их основные составляющие — нуклеотиды и нуклеозиды, а также предложили структурные формулы, верно описывающие их химические свойства.
- В 1921 году Левин выдвинул гипотезу «тетрануклеотидной структуры ДНК»[3], оказавшуюся впоследствии ошибочной[4].
- В 1935 году Клейн и Танхаузер [de] с помощью фермента фосфатазы провели мягкое фрагментирование ДНК, в результате чего были получены в кристаллическом состоянии четыре ДНК-образующих нуклеотида[5]. Это открыло новые возможности для установления структуры этих соединений.
- В 1940-е годы научная группа в Кембридже под руководством Александера Тодда проводит широкие синтетические исследования в области химии нуклеотидов и нуклеозидов, в результате которых были установлены детали химического строения и стереохимии нуклеотидов. За этот цикл работ Александер Тодд был награждён Нобелевской премией в области химии в 1957 году.
- В 1951 году Чаргаффом была установлена закономерность содержания в нуклеиновых кислотах нуклеотидов разных типов, получившая впоследствии название Правило Чаргаффа.
- В 1953 году Уотсоном и Криком установлена вторичная структура ДНК, двойная спираль[6].
Способы выделения

Описаны многочисленные методики выделения нуклеиновых кислот из природных источников. Основными требованиями, предъявляемыми к методу выделения, являются эффективное отделение нуклеиновых кислот от белков, а также минимальная степень фрагментации полученных препаратов. Классический метод выделения ДНК был описан в 1952 году и используется в настоящее время без значительных изменений[7]. Клеточные стенки исследуемого биологического материала разрушаются одним из стандартных методов, а затем обрабатываются анионным детергентом. При этом белки выпадают в осадок, а нуклеиновые кислоты остаются в водном растворе. ДНК может быть осаждена в виде геля осторожным добавлением этанола к её солевому раствору. Концентрацию полученной нуклеиновой кислоты, а также наличие примесей (белки, фенол) обычно определяют спектрофотометрически по поглощению на А260 нм.
Нуклеиновые кислоты легко деградируют под действием особого класса ферментов — нуклеаз. В связи с этим при их выделении важно обработать лабораторное оборудование и материалы соответствующими ингибиторами. Так, например, при выделении РНК широко используется такой ингибитор рибонуклеаз, как DEPC.
Физические свойства
Нуклеиновые кислоты хорошо растворимы в воде, практически нерастворимы в органических растворителях. Очень чувствительны к действию температуры и критическим значениям уровня pH. Молекулы ДНК с высокой молекулярной массой, выделенные из природных источников, способны фрагментироваться под действием механических сил, например, при перемешивании раствора. Нуклеиновые кислоты фрагментируются ферментами — нуклеазами.
Строение

Полимерные формы нуклеиновых кислот называют полинуклеотидами. Различают четыре уровня структурной организации нуклеиновых кислот: первичную, вторичную, третичную и четвертичную структуры. Первичная структура представляет собой цепочки из нуклеотидов, соединяющихся через остаток фосфорной кислоты (фосфодиэфирная связь). Вторичная структура — это две цепи нуклеиновых кислот, соединённые водородными связями. Цепи соединяются по типу «голова-хвост» (3' к 5'), по принципу комплементарности (азотистые основания находятся внутри этой структуры). Третичная структура, или же спираль, образуется за счет радикалов азотистых оснований (образуются водородные дополнительные связи, которые и сворачивают эту структуру, тем самым обуславливая её прочность). И, наконец, четвертичная структура — это комплексы гистонов и нитей хроматина.
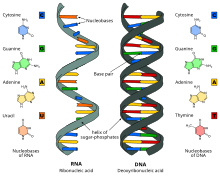
Поскольку в нуклеотидах существует только два типа гетероциклических молекул, рибоза и дезоксирибоза, то и имеется лишь два вида нуклеиновых кислот — дезоксирибонуклеиновая (ДНК) и рибонуклеиновая (РНК).
Мономерные формы также встречаются в клетках и играют важную роль в процессах передачи сигналов или запасании энергии. Наиболее известный мономер РНК — АТФ, аденозинтрифосфорная кислота, важнейший аккумулятор энергии в клетке.
ДНК и РНК
- ДНК (дезоксирибонуклеиновая кислота). Сахар — дезоксирибоза, азотистые основания: пуриновые — гуанин (G), аденин (A), пиримидиновые — тимин (T) и цитозин (C). ДНК часто состоит из двух полинуклеотидных цепей, направленных антипараллельно.
- РНК (рибонуклеиновая кислота). Сахар — рибоза, азотистые основания: пуриновые — гуанин (G), аденин (A), пиримидиновые — урацил (U) и цитозин (C). Структура полинуклеотидной цепочки аналогична таковой в ДНК. Из-за особенностей рибозы молекулы РНК часто имеют различные вторичные и третичные структуры, образуя комплементарные участки между разными цепями.
Типы РНК
Ма́тричная рибонуклеи́новая кислота́ (мРНК, синоним — информацио́нная РНК, иРНК) — РНК, содержащая информацию о первичной структуре (аминокислотной последовательности) белков[8]. мРНК синтезируется на основе ДНК в ходе транскрипции, после чего, в свою очередь, используется в ходе трансляции как матрица для синтеза белков. Тем самым мРНК играет важную роль в «проявлении» (экспрессии) генов.
Рибосо́мные рибонуклеи́новые кисло́ты (рРНК) — несколько молекул РНК, составляющих основу рибосомы. Основной функцией рРНК является осуществление процесса трансляции — считывания информации с мРНК при помощи адапторных молекул тРНК и катализ образования пептидных связей между присоединёнными к тРНК аминокислотами.

Транспортная РНК, тРНК — рибонуклеиновая кислота, функцией которой является транспортировка аминокислот к месту синтеза белка. Имеет типичную длину от 73 до 93 нуклеотидов и размеры около 5 нм. тРНК также принимают непосредственное участие в наращивании полипептидной цепи, присоединяясь — будучи в комплексе с аминокислотой — к кодону мРНК и обеспечивая необходимую для образования новой пептидной связи конформацию комплекса.
Для каждой аминокислоты существует своя тРНК.
тРНК является одноцепочечной РНК, однако в функциональной форме имеет конформацию «клеверного листа». Аминокислота ковалентно присоединяется к 3'-концу молекулы с помощью специфичного для каждого типа тРНК фермента аминоацил-тРНК-синтетазы. На участке C находится антикодон, соответствующий аминокислоте.
Некодирующие РНК (non-coding RNA, ncRNA) — это молекулы РНК, которые не транслируются в белки. Ранее использовавшийся синоним, малые РНК (smRNA, small RNA), в настоящее время не используется, так как некоторые некодирующие РНК могут быть очень большими, например, Xist.
Последовательность ДНК, на которой транскрибируются некодирующие РНК, часто называют РНК-геном.
В число некодирующих РНК входят молекулы РНК, которые выполняют очень важные функции в клетке — транспортные РНК (тРНК), рибосомные РНК (рРНК), такие малые РНК, как малые ядрышковые РНК (snoRNA), микроРНК, siRNA, piRNA, а также длинные некодирующие РНК — Xist, Evf, Air, CTN, PINK, TUG1.
Последние транскриптомные технологии (секвенирование РНК) и методы ДНК-микрочипов предполагают наличие более чем 30 000 длинных некодирующих РНК (англ. long ncRNA). Примерно такое же количество малых регуляторных РНК содержится в геноме мыши.
Примечания
- ↑ J. Liebig. ??? (неопр.) // Annalen. — 1847. — Т. 62. — С. 257.
- ↑ Edmund B. Wilson. An Atlas of the Fertilization and Karyokinesis of the Ovum. — N. Y.: Macmillan, 1895. — P. 4.
- ↑ P. A. Levene. ??? (англ.) // J. Biol. Chem. : journal. — 1921. — Vol. 48. — P. 119.
- ↑ Во время выдвижения «тетрануклеотидной структуры» химики критически относились к самой возможности существования макромолекул, вследствие чего ДНК была приписана структура с низкой молекулярной массой
- ↑ W. Klein, S. J. Thannhauser. ??? (неопр.) // Z. physiol. Chem.. — 1935. — Т. 231. — С. 96.
- ↑ J. D. Watson, F. H. C. Crick. Molecular Structure of Nucleic Acids: A Structure for Deoxyribose Nucleic Acid (англ.) // Nature. — 1953. — Vol. 171. — P. 737—738. — doi:10.1038/171737a0.
- ↑ Ernest R. M. Kay, Norman S. Simmons, Alexander L. An Improved Preparation of Sodium Desoxyribonucleate (англ.) // J. Am. Chem. Soc.[англ.] : journal. — 1952. — Vol. 74, no. 7. — P. 1724—1726. — doi:10.1021/ja01127a034.
- ↑ Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter. Molecular Biology of the Cell. — 5. — Garland Science, 2008. — 1392 с. — ISBN 0815341059.
Литература
- Бартон Д., Оллис У. Д. Общая органическая химия. — М.: Химия, 1986. — Т. 10. — С. 32—215. — 704 с.
- Франк-Каменецкий М. Д. Самая главная молекула. — М.: Наука, 1983. — 160 с.
- Аппель Б., Бенеке И., Бенсон Я., под ред. С. Мюллер. Нуклеиновые кислоты от А до Я. — М.: Бином, 2012. — 352 с. — ISBN 978-5-9963-0376-2.
Основные группы биохимических молекул | |
|---|---|
Типы нуклеиновых кислот | ||||
|---|---|---|---|---|
| Азотистые основания | ||||
| Нуклеозиды | ||||
| Нуклеотиды | ||||
| РНК | ||||
| ДНК | ||||
| Аналоги | ||||
| Типы векторов |
| |||
| ||||