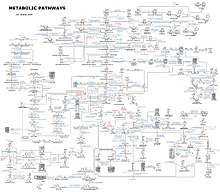
Запрос «Метаболизм» перенаправляется сюда; об архитектурной концепции см. Метаболизм (архитектура).
Упрощённая схема клеточного метаболизма
Метаболи́зм, или обме́н веще́ств, — это химические реакции, поддерживающие жизнь в живом организме. Эти процессы позволяют организмам расти и размножаться, сохранять свои структуры и отвечать на воздействия окружающей среды.
Метаболизм обычно делят на 2 стадии: катаболизм и анаболизм. В ходе катаболизма сложные органические вещества деградируют до более простых, обычно выделяя энергию, а в процессах анаболизма — более сложные вещества синтезируются из более простых с затратами энергии.
Серии химических реакций обмена веществ называют метаболическими путями. В них, при участии ферментов, одни биологически значимые молекулы последовательно превращаются в другие.
Ферменты играют важную роль в метаболических процессах, потому что:
позволяют регулировать метаболические пути в ответ на изменения среды клетки или сигналы от других клеток.
Особенности метаболизма влияют на то, будет ли пригодна определённая молекула для использования организмом в качестве источника энергии. Так, например, некоторые прокариоты используют сероводород в качестве источника энергии, однако этот газ ядовит для животных[1]. Скорость обмена веществ также влияет на количество пищи, необходимой для организма.
Основные метаболические пути и их компоненты одинаковы для многих видов, что свидетельствует о единстве происхождения всех живых существ[2]. Например, некоторые карбоновые кислоты, являющиеся интермедиатамицикла трикарбоновых кислот, присутствуют во всех организмах, начиная от бактерий и заканчивая многоклеточными организмами эукариот[3]. Сходства в обмене веществ, вероятно, связаны с высокой эффективностью метаболических путей, а также с их ранним появлением в истории эволюции[4][5].
Биологические молекулы
Структура липида триглицеридаДиаграмма, изображающая большой набор метаболических путей человека.
Органические вещества, входящие в состав всех живых существ (животных, растений, грибов и микроорганизмов), представлены в основном аминокислотами, углеводами, липидами (часто называемые жирами) и нуклеиновыми кислотами. Так как эти молекулы имеют важное значение для жизни, метаболические реакции сосредоточены на создании этих молекул при строительстве клеток и тканей или разрушении их с целью использования в качестве источника энергии. Многие важные биохимические реакции объединяются вместе для синтеза ДНК и белков.
Белки являются биополимерами и состоят из остатков аминокислот, соединённых пептидными связями. Некоторые белки являются ферментами и катализируют химические реакции. Другие белки выполняют структурную или механическую функцию (например образуют цитоскелет)[6]. Белки также играют важную роль в передаче сигнала в клетках, иммунных реакциях, агрегации клеток, активном транспорте через мембраны и регуляции клеточного цикла[7]. Аминокислоты также способствуют клеточному энергетическому метаболизму, обеспечивая источник углерода для входа в цикл лимонной кислоты (цикл трикарбоновых кислот)[8], особенно когда основного источника энергии, такого как глюкоза, недостаточно, или когда клетки подвергаются метаболическому стрессу[9].
Липиды — это самая разнообразная группа биохимических веществ. Они входят в состав биологических мембран, например плазматических мембран, являются компонентами коферментов и источниками энергии[7]. Липиды представляют собой полимеры жирных кислот, которые содержат длинную неполярную углеводородную цепь с небольшой полярной областью, содержащей кислород. Липиды являются гидрофобными или амфифильными биологическими молекулами, растворимыми в органических растворителях, таких как бензол или хлороформ[10]. Жиры — большая группа соединений, в состав которых входят жирные кислоты и глицерин. Молекула трёхатомного спирта глицерина, образующая три сложные эфирные связи с тремя молекулами жирных кислот, называется триглицеридом[11]. Наряду с остатками жирных кислот, в состав сложных липидов может входить, например, сфингозин (сфинголипиды), гидрофильные группы фосфатов (в фосфолипидах). Стероиды, например холестерол, представляют собой ещё один большой класс липидов[12].
Глюкоза может существовать как в форме прямой цепи, так и в форме кольца.
Сахара могут существовать в кольцевой или линейной форме в виде альдегидов или кетонов, имеют несколько гидроксильных групп. Углеводы являются наиболее распространёнными биологическими молекулами. Углеводы выполняют следующие функции: хранение и транспортировка энергии (крахмал, гликоген), структурная (целлюлоза растений, хитин у грибов и животных)[7]. Наиболее распространёнными мономерами сахаров являются гексозы — глюкоза, фруктоза и галактоза. Моносахариды входят в состав более сложных линейных или разветвлённых полисахаридов[13].
Полимерные молекулы ДНК и РНК представляют собой длинные неразветвлённые цепочки нуклеотидов. Нуклеиновые кислоты выполняют функцию хранения и реализации генетической информации, которые осуществляются в ходе процессов репликации, транскрипции, трансляции и биосинтеза белка[7]. Информация, закодированная в нуклеиновых кислотах, защищается от изменений системами репарации и мультиплицируется при помощи репликации ДНК.
Некоторые вирусы имеют РНК-содержащий геном. Например, вирус иммунодефицита человека использует обратную транскрипцию для создания матрицы ДНК из собственного РНК-содержащего генома[14]. Некоторые молекулы РНК обладают каталитическими свойствами (рибозимы) и входят в состав сплайсосом и рибосом.
Метаболизм включает широкий спектр химических реакций, большинство из которых относится к нескольким основным типам реакций переноса функциональных групп[16]. Для переноса функциональных групп между ферментами, катализирующими химические реакции, используются коферменты[15]. Каждый класс химических реакций переноса функциональных групп катализируется отдельными ферментами и их кофакторами[17].
Аденозинтрифосфат (АТФ) — один из центральных коферментов, универсальный источник энергии клеток. Этот нуклеотид используется для передачи химической энергии, запасённой в макроэргических связях, между различными химическими реакциями. В клетках существует небольшое количество АТФ, который постоянно регенерируется из AДФ и AМФ. Организм человека за сутки расходует массу АТФ, равную массе собственного тела[17]. АТФ выступает в качестве связующего звена между катаболизмом и анаболизмом: при катаболических реакциях образуется АТФ, при анаболических — энергия потребляется. АТФ также выступает донором фосфатной группы в реакциях фосфорилирования.
Витамины — низкомолекулярные органические вещества, необходимые в небольших количествах, причём, например, у человека большинство витаминов не синтезируется, а получается с пищей или через микрофлору ЖКТ. В организме человека большинство витаминов являются кофакторами ферментов. Большинство витаминов приобретает биологическую активность в изменённом виде, например, все водорастворимые витамины в клетках фосфорилируются или соединяются с нуклеотидами[18]. Никотинамидадениндинуклеотид (NADH) является производным витамина B3 (ниацина) и представляет собой важный кофермент — акцептора водорода. Сотни различных ферментов дегидрогеназ отнимают электроны из молекул субстратов и переносят их на молекулы NAD+, восстанавливая его до NADH. Окисленная форма кофермента является субстратом для различных редуктаз в клетке[19]. NAD в клетке существует в двух связанных формах — NADH и NADPH. NAD+/NADH больше важен для протекания катаболических реакций, а NADP+/NADPH чаще используется в анаболических реакциях.
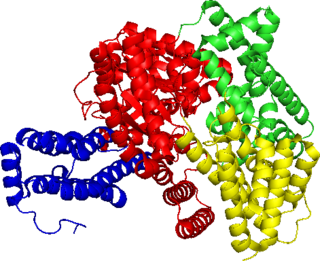
Структура гемоглобина. Белковые субъединицы окрашены красным и синим, а железосодержащий гем — зелёным. Из PDB1GZX
Неорганические элементы играют важнейшую роль в обмене веществ. Около 99 % массы млекопитающего состоит из углерода, азота, кальция, натрия, магния, хлора, калия, водорода, фосфора, кислорода и серы[20]. Биологически значимые органические соединения (белки, жиры, углеводы и нуклеиновые кислоты) содержат большое количество углерода, водорода, кислорода, азота и фосфора[20].
Многие неорганические соединения являются ионнымиэлектролитами. Наиболее важны для организма ионы натрия, калия, кальция, магния, хлоридов, фосфатов и гидрокарбонатов. Баланс этих ионов внутри клетки и во внеклеточной среде определяет осмотическое давление и pH[21]. Концентрации ионов также играют важную роль для функционирования нервных и мышечных клеток. Потенциал действия в возбудимых тканях возникает при обмене ионами между внеклеточной жидкостью и цитоплазмой[22]. Электролиты входят и выходят из клетки через ионные каналы в плазматической мембране. Например, в ходе мышечного сокращения в плазматической мембране, цитоплазме и Т-трубочках перемещаются ионы кальция, натрия и калия[23].
Переходные металлы в организме являются микроэлементами, наиболее распространены цинк и железо[24][25]. Эти металлы используются некоторыми белками (например ферментами в качестве кофакторов) и имеют важное значение для регуляции активности ферментов и транспортных белков[26]. Кофакторы ферментов обычно прочно связаны со специфическим белком, однако могут модифицироваться в процессе катализа, при этом после окончания катализа всегда возвращаются к своему первоначальному состоянию (не расходуются). Металлы-микроэлементы усваиваются организмом при помощи специальных транспортных белков и не встречаются в организме в свободном состоянии, так как связаны со специфическими белками-переносчиками (например ферритином или металлотионеинами)[27][28].
Классификация организмов по типу метаболизма
Все живые организмы можно разделить на восемь основных групп в зависимости от используемого: источника энергии, источника углерода и донора электронов (окисляемого субстрата)[29].
В качестве источника энергии живые организмы могут использовать: энергию света (фото-) или энергию химических связей (хемо-). Дополнительно для описания паразитических организмов, использующих энергетические ресурсы хозяйской клетки, применяют термин паратроф.
Помимо источника энергии живым организмам также требуется донор электронов, окисляемое вещество, от которого отрывается электрон, который используется для синтеза органики. В качестве донора электронов (восстановителя) живые организмы могут использовать: неорганические вещества (лито-) или органические вещества (органо-).
В качестве источника углерода живые организмы используют: углекислый газ (авто-) или органические вещества (гетеро-). Иногда термины авто- и гетеротроф используют в отношении других элементов, которые входят в состав биологических молекул в восстановленной форме (например азота, серы). В таком случае «автотрофными по азоту» организмами являются виды, использующие в качестве источника азота окисленные неорганические соединения (например растения; могут осуществлять восстановление нитратов). А «гетеротрофными по азоту» являются организмы, не способные осуществлять восстановление окисленных форм азота и использующие в качестве его источника органические соединения (например животные, для которых источником азота служат аминокислоты).
Название типа метаболизма формируется путём сложения соответствующих корней и добавлением в конце корня -троф-. В таблице представлены возможные типы метаболизма с примерами[30]:
Некоторые авторы используют -гидро-, когда в качестве донора электронов выступает вода
CO2, CO, HCHO, CH3OH, CH4 HCOO- и «неорганическая» метильная группа -СH3, присоединённая через атом кислорода, азота или серы к другим метильным группам (от одной до трёх) или к многоуглеродному скелету[31].
Классификация была разработана группой авторов (А. Львов, К. ван Ниль, F. J. Ryan, Э. Тейтем) и утверждена на 11-м симпозиуме в лаборатории Колд-Спринг-Харбор и изначально служила для описания типов питания микроорганизмов. Однако в настоящее время применяется и для описания метаболизма других организмов[32].
Из таблицы очевидно, что метаболические возможности прокариот значительно разнообразнее по сравнению с эукариотами, которые характеризуются фотолитоавтотрофным и хемоорганогетеротрофным типом метаболизма.
Следует отметить, что некоторые виды микроорганизмов могут в зависимости от условий среды (освещение, доступность органических веществ и т. д.) и физиологического состояния осуществлять метаболизм разного типа. Такое сочетание нескольких типов метаболизма описывается как миксотрофия.
При применении данной классификации к многоклеточным организмам важно понимать, что в рамках одного организма могут быть клетки, отличающиеся типом обмена веществ. Так клетки надземных, фотосинтезирующих органов многоклеточных растений характеризуются фотолитоавтотрофным типом метаболизма, в то время как клетки подземных органов описываются как хемоорганогетеротрофные. Так же как и в случае с микроорганизмами, при изменении условий среды, стадии развития и физиологического состояния тип метаболизма клеток многоклеточного организма может изменяться. Так, например, в темноте и на стадии прорастания семени клетки высших растений осуществляют метаболизм хемоорганогетеротрофного типа.
Катаболизмом называют метаболические процессы, при которых расщепляются относительно крупные органические молекулы сахаров, жиров, аминокислот. В ходе катаболизма образуются более простые органические молекулы, необходимые для реакций анаболизма (биосинтеза). Часто именно в ходе реакций катаболизма организм мобилизует энергию, переводя энергию химических связей органических молекул, полученных в процессе переваривания пищи, в доступные формы: в виде АТФ, восстановленных коферментов и трансмембранного электрохимического потенциала. Термин катаболизм не является синонимом «энергетического обмена»: у многих организмов (например у фототрофов) основные процессы запасания энергии не связаны напрямую с расщеплением органических молекул. Классификация организмов по типу метаболизма может быть основана на источнике получения энергии, что отражено в предыдущем разделе. Энергию химических связей используют хемотрофы, а фототрофы потребляют энергию солнечного света. Однако все эти различные формы обмена веществ зависят от окислительно-восстановительных реакций, которые связаны с передачей электронов от восстановленных доноров молекул, таких как органические молекулы, вода, аммиак, сероводород, на акцепторные молекулы, такие как кислород, нитраты или сульфат[33]. У животных эти реакции сопряжены с расщеплением сложных органических молекул до более простых, таких как двуокись углерода и воду. В фотосинтезирующих организмах — растениях и цианобактериях — реакции переноса электрона не высвобождают энергию, но они используются как способ запасания энергии, поглощаемой из солнечного света[34].
Катаболизм у животных может быть разделён на три основных этапа. Во-первых, крупные органические молекулы, такие как белки, полисахариды и липиды, расщепляются до более мелких компонентов вне клеток. Далее эти небольшие молекулы попадают в клетки и превращается в ещё более мелкие молекулы, например ацетил-КоА. В свою очередь, ацетильная группа кофермента А окисляется до воды и углекислого газа в цикле Кребса и дыхательной цепи, высвобождая при этом энергию, которая запасается в форме АТР.
Такие макромолекулы, как крахмал, целлюлоза или белки, должны расщепляться до более мелких единиц прежде, чем они могут быть использованы клетками. Несколько классов ферментов принимают участие в деградации: протеазы, которые расщепляют белки до пептидов и аминокислот, гликозидазы, которые расщепляют полисахариды до олиго- и моносахаридов.
Микроорганизмы выделяют гидролитические ферменты в пространство вокруг себя[35][36], чем отличаются от животных, которые выделяют такие ферменты только из специализированных железистых клеток[37]. Аминокислоты и моносахариды, образующиеся в результате активности внеклеточных ферментов, затем поступают в клетки с помощью активного транспорта[38][39].
Упрощённая схема катаболизма белков, сахаров и липидов
В ходе катаболизма углеводов сложные сахара расщепляются до моносахаридов, которые усваиваются клетками[40]. Попав внутрь, сахара (например глюкоза и фруктоза) в процессе гликолиза превращаются в пируват, при этом вырабатывается некоторое количество АТФ[41]. Пировиноградная кислота (пируват) является промежуточным продуктом в нескольких метаболических путях. Основной путь метаболизма пирувата — превращение в ацетил-КоА и далее поступление в цикл трикарбоновых кислот. При этом в цикле Кребса в форме АТР запасается часть энергии, а также восстанавливаются молекулы NADH и FAD. В процессе гликолиза и цикла трикарбоновых кислот образуется диоксид углерода, который является побочным продуктом жизнедеятельности. В анаэробных условиях в результате гликолиза из пирувата при участии фермента лактатдегидрогеназы образуется лактат и происходит окисление NADH до NAD+, который повторно используется в реакциях гликолиза. Существует также альтернативный путь метаболизма моносахаридов — пентозофосфатный путь, в ходе реакций которого энергия запасается в форме восстановленного кофермента NADPH и образуются пентозы, например рибоза, необходимая для синтеза нуклеиновых кислот.
Жиры на первом этапе катаболизма гидролизуются в свободные жирные кислоты и глицерин. Жирные кислоты расщепляются в процессе бета-окисления с образованием ацетил-КоА, который в свою очередь далее катаболизируется в цикле Кребса, либо идёт на синтез новых жирных кислот. Жирные кислоты выделяют больше энергии, чем углеводы, так как жиры содержат удельно больше атомов водорода в своей структуре.
Аминокислоты либо используются для синтеза белков и других биомолекул, либо окисляются до мочевины, диоксида углерода и служат источником энергии[42]. Окислительный путь катаболизма аминокислот начинается с удаления аминогруппы ферментами трансаминазами. Аминогруппы утилизируются в цикле мочевины; аминокислоты, лишённые аминогрупп, называют кетокислотами. Некоторые кетокислоты — промежуточные продукты цикла Кребса. Например, при дезаминировании глутамата образуется альфа-кетоглутаровая кислота[43]. Гликогенные аминокислоты также могут быть преобразованы в глюкозу в реакциях глюконеогенеза[44].
При окислительном фосфорилировании электроны, удалённые из пищевых молекул в метаболических путях (например в цикле Кребса), переносятся на кислород, а выделяющаяся энергия используется для синтеза АТФ. У эукариот данный процесс осуществляется при участии ряда белков, закреплённых в мембранах митохондрий, называемых дыхательной цепью переноса электронов. У прокариот эти белки присутствуют во внутренней мембране клеточной стенки[45]. Белки цепи переноса электронов используют энергию, полученную при передаче электронов от восстановленных молекул (например NADH) на кислород, для перекачки протонов через мембрану[46].
Механизм действия АТФ-синтазы. АТФ показан красным цветом, АДФ и фосфат — розовым, а вращающаяся стеблевая субъединица — чёрным.
При перекачке протонов создаётся разница концентраций ионов водорода и возникает электрохимический градиент[47]. Эта сила возвращает протоны обратно в митохондрии через основание АТФ-синтазы. Поток протонов заставляет вращаться кольцо из c-субъединиц фермента, в результате чего активный центр синтазы изменяет форму и фосфорилирует аденозиндифосфат, превращая его в АТФ[17].
Энергия из неорганических соединений
Хемолитотрофами называют прокариот, имеющих особый тип обмена веществ, при котором энергия образуется в результате окисления неорганических соединений. Хемолитотрофы могут окислять молекулярный водород[48], соединения серы (например сульфиды, сероводород и неорганические тиосульфаты)[1], оксид железа(II)[49] или аммиак[50]. При этом энергия от окисления этих соединений образуется с помощью акцепторов электронов, таких как кислород или нитриты[51]. Процессы получения энергии из неорганических веществ играют важную роль в таких биогеохимических циклах, как ацетогенез, нитрификация и денитрификация[52][53].
Энергия из солнечного света
Энергия солнечного света поглощается растениями, цианобактериями, пурпурными бактериями, зелёными серными бактериями и некоторыми простейшими. Этот процесс часто сочетается с превращением диоксида углерода в органические соединения, как часть процесса фотосинтеза (см. ниже). Системы захвата энергии и фиксации углерода у некоторых прокариот могут работать раздельно (например у пурпурных и зелёных серных бактерий)[54][55].
У многих организмов поглощение солнечной энергии в принципе аналогично окислительному фосфорилированию, так как при этом энергия запасается в форме градиента концентрации протонов и движущая сила протонов приводит к синтезу АТФ[17]. Электроны, необходимые для этой цепи переноса, поступают от светособирающих белков, называемых центрами фотосинтетических реакций (примером являются родопсины). В зависимости от вида фотосинтетических пигментов классифицируют два типа центров реакций; в настоящее время большинство фотосинтезирующих бактерий имеют только один тип, в то время как растения и цианобактерии два[56].
У растений, водорослей и цианобактерий фотосистема II использует энергию света для удаления электронов из воды, при этом молекулярный кислород выделяется как побочный продукт реакции. Электроны затем поступают в комплекс цитохрома b6f, который использует энергию для перекачки протонов через тилакоидную мембрану в хлоропластах[7]. Под действием электрохимического градиента протоны движутся обратно через мембрану и запускают АТР-синтазу. Электроны затем проходят через фотосистему I и могут быть использованы для восстановления кофермента NADP+, для использования в цикле Кальвина или рециркуляции для образования дополнительных молекул АТР[57].
Анаболизм — совокупность метаболических процессов биосинтеза сложных молекул с затратой энергии. Сложные молекулы, входящие в состав клеточных структур, синтезируются последовательно из более простых предшественников. Анаболизм включает три основных этапа, каждый из которых катализируется специализированным ферментом. На первом этапе синтезируются молекулы-предшественники, например аминокислоты, моносахариды, терпеноиды и нуклеотиды. На втором этапе предшественники с затратой энергии АТФ преобразуются в активированные формы. На третьем этапе активированные мономеры объединяются в более сложные молекулы, например белки, полисахариды, липиды и нуклеиновые кислоты.
Не все живые организмы могут синтезировать все биологически активные молекулы. Автотрофы (например растения) могут синтезировать сложные органические молекулы из таких простых неорганических низкомолекулярных веществ, как углекислый газ и вода. Гетеротрофам необходим источник более сложных веществ, таких как моносахариды и аминокислоты, для создания более сложных молекул. Организмы классифицируют по их основным источникам энергии: фотоавтотрофы и фотогетеротрофы получают энергию из солнечного света, в то время как хемоавтотрофы и хемогетеротрофы получают энергию из неорганических реакций окисления.
Растительные клетки содержат хлоропласты (зелёного цвета), в тилакоидах которых происходят процессы фотосинтеза. Plagiomnium affine из семейства Mniaceae отдела Настоящие мхи (Bryophyta)
Фотосинтезом называют процесс биосинтеза сахаров из углекислого газа, при котором необходимая энергия поглощается из солнечного света. У растений, цианобактерий и водорослей при кислородном фотосинтезе происходит фотолиз воды, при этом как побочный продукт выделяется кислород. Для преобразования CO2 в 3-фосфоглицерат используется энергия АТФ и НАДФ, запасённая в фотосистемах. Реакция связывания углерода осуществляется с помощью фермента рибулозобисфосфаткарбоксилазы и является частью цикла Кальвина[58]. У растений классифицируют три типа фотосинтеза — по пути трёхуглеродных молекул, по пути четырёхуглеродых молекул (С4), и CAM фотосинтез. Три типа фотосинтеза отличаются по пути связывания углекислого газа и его вхождения в цикл Кальвина; у C3 растений связывание CO2 происходит непосредственно в цикле Кальвина, а при С4 и CAM CO2 предварительно включается в состав других соединений. Разные формы фотосинтеза являются приспособлениями к интенсивному потоку солнечных лучей и к сухим условиям[59].
У фотосинтезирующих прокариот механизмы связывания углерода более разнообразны. Углекислый газ может быть фиксирован в цикле Кальвина, в обратном цикле Кребса[60] или в реакциях карбоксилирования ацетил-КоА[61][62]. Прокариоты — хемоавтотрофы также связывают CO2 через цикл Кальвина, но для протекания реакции используют энергию из неорганических соединений[63].
В процессе анаболизма сахаров простые органические кислоты могут быть преобразованы в моносахариды, например в глюкозу, и затем использованы для синтеза полисахаридов, таких как крахмал. Образование глюкозы из соединений, таких как пируват, лактат, глицерин, 3-фосфоглицерат и аминокислот, называют глюконеогенезом. В процессе глюконеогенеза пируват превращается в глюкозо-6-фосфат через ряд промежуточных соединений, многие из которых образуются и при гликолизе[41]. Однако глюконеогенез не просто является гликолизом в обратном направлении, так как несколько химических реакций катализируют специальные ферменты, что даёт возможность независимо регулировать процессы образования и распада глюкозы[64][65].
Многие организмы запасают питательные вещества в форме липидов и жиров, однако позвоночные не имеют ферментов, катализирующих превращение ацетил-КоА (продукта метаболизма жирных кислот) в пируват (субстрат глюконеогенеза)[66]. После длительного голодания позвоночные начинают синтезировать кетоновые тела из жирных кислот, которые могут заменять глюкозу в таких тканях, как головной мозг[67]. У растений и бактерий данная метаболическая проблема решается использованием глиоксилатного цикла, который обходит этап декарбоксилирования в цикле лимонной кислоты и позволяет превращать ацетил-КоА в оксалоацетат и далее использовать для синтеза глюкозы[66][68]. Помимо жира, глюкоза хранится в большинстве тканей в качестве энергетического ресурса, доступного в тканях посредством гликогенеза, который обычно используется для поддержания уровня глюкозы в крови[69].
Полисахариды и гликаны образуются путём последовательного добавления моносахаридов гликозилтрансферазой из реактивного донора сахара-фосфата, такого как уридиндифосфат глюкозы (UDP-Glc), к акцепторной гидроксильной группе на растущем полисахариде. Поскольку любая из гидроксильных групп в кольце субстрата может быть акцепторами, полученные полисахариды могут иметь прямые или разветвлённые структуры[70]. Полисахариды выполняют структурные и метаболические функции, а также могут быть соединены с липидами (гликолипиды) и белками (гликопротеиды) при помощи ферментов олигосахаридтрансфераз[71][72].
Синтез стероидов из изопентилпирофосфата, диметилаллилпирофосфата, геранилпирофосфата и сквалена. Некоторые промежуточные продукты не показаны
Жирные кислоты образуются синтазами жирных кислот из ацетил-КоА. Углеродный скелет жирных кислот удлиняется в цикле реакций, в которых сначала присоединяется ацетильная группа, далее карбонильная группа восстанавливается до гидроксильной, затем происходит дегидратация и последующее восстановление. Ферменты биосинтеза жирных кислот классифицируют на две группы: у животных и грибов все реакции синтеза жирных кислот осуществляются одним многофункциональным белком I типа[73], в пластидах растений и у бактерий каждый этап катализируют отдельные ферменты II типа[74][75].
Терпены и терпеноиды являются представителями самого многочисленного класса растительных натуральных продуктов[76]. Представители данной группы веществ являются производными изопрена и образуются из активированных предшественников изопентилпирофосфата и диметилаллилпирофосфата, которые, в свою очередь, образуются в разных реакциях обмена веществ[77]. У животных и архей изопентилпирофосфат и диметилаллилпирофосфат синтезируются из ацетил-КоА в мевалонатном пути[78], в то время как у растений и бактерий субстратами не-мевалонатного пути являются пируват и глицеральдегид-3-фосфат[77][79]. В реакциях биосинтеза стероидов молекулы изопрена объединяются и образуют сквалены, которые далее формируют циклические структуры с образованием ланостерола[80]. Ланостерол может быть преобразован в другие стероиды, например холестерин и эргостерин[80][81].
Организмы различаются по способности к синтезу 20 общих аминокислот. Большинство бактерий и растений могут синтезировать все 20, но млекопитающие способны синтезировать лишь 10 заменимых аминокислот[7]. Таким образом, в случае млекопитающих 9 незаменимых аминокислот должны быть получены из пищи. Некоторые простые паразиты, такие как бактерии Mycoplasma pneumoniae, не синтезируют все аминокислоты и получают их непосредственно от своих хозяев[82]. Все аминокислоты синтезируются из промежуточных продуктов гликолиза, цикла лимонной кислоты или пентозомонофосфатного пути. Перенос аминогрупп с аминокислот на альфа-кетокислоты называется трансаминированием. Донорами аминогрупп являются глутамат и глутамин[83]. Второстепенный синтез аминокислот зависит от образования соответствующей альфа-кетокислоты, которая затем подвергается трансаминированию с образованием аминокислоты[84].
Аминокислоты, соединёнными пептидными связями, образуют белки. Каждый белок имеет уникальную последовательность аминокислотных остатков (первичная структура белка). Подобно тому, как буквы алфавита могут комбинироваться с образованием почти бесконечных вариаций слов, аминокислоты могут связываться в той или иной последовательности и формировать разнообразные белки. Белки состоят из аминокислот, которые были активированы путём присоединения к молекуле переносной РНК через эфирную связь. Фермент Аминоацил-тРНК-синтетаза катализирует АТР-зависимое присоединение аминокислот к тРНК сложноэфирными связями, при этом образуются аминоацил-тРНК[85]. Аминоацил-тРНК являются субстратами для рибосом, которая объединяют аминокислоты в длинные полипептидные цепочки, используя матрицу мРНК[86].
Нуклеотиды образуются из аминокислот, углекислого газа и муравьиной кислоты в цепи реакций, для протекания которых требуется большое количество энергии[87][88]. Именно поэтому большинство организмов имеют эффективные системы сохранения ранее синтезированных нуклеотидов и азотистых оснований[87][89]. Пурины синтезируются как нуклеозиды (в основном связанные с рибозой). Аденин и гуанин образуются из инозин-монофосфата, который синтезируется из глицина, глутамина и аспартата при участии метенил-тетрагидрофолата. Пиримидины синтезируются из оротата, который образуется из глутамина и аспартата[90].
Все организмы постоянно подвергаются воздействию соединений, накопление которых может быть вредно для клеток. Такие потенциально опасные чужеродные соединения называются ксенобиотиками[91]. Ксенобиотики, например синтетические лекарства и яды природного происхождения, детоксифицируются специализированными ферментами. У человека такие ферменты представлены, например, цитохром-оксидазами[92], глюкуронилтрансферазой[93], и глутатион S-трансферазой[94]. Эта система ферментов действует в три этапа: на первой стадии ксенобиотики окисляются, затем происходит конъюгирование водорастворимых групп в молекулы, далее модифицированные водорастворимые ксенобиотики могут быть удалены из клеток и метаболизированы перед их экскрецией. Описанные реакции играют важную роль в разложении микробами загрязняющих веществ и биоремедиации загрязнённых земель и разливов нефти[95]. Многие подобные реакции протекают при участии многоклеточных организмов, однако, ввиду невероятного разнообразия, микроорганизмы справляются с гораздо более широким спектром ксенобиотиков, чем многоклеточные организмы, и способны даже разрушать стойкие органические загрязнители, например хлорорганические соединения[96].
Живые организмы подчиняются началам термодинамики, которые описывают превращения тепла и работы. Второе начало термодинамики гласит, что в любой изолированной системе энтропия не уменьшается. Хотя может показаться, что невероятная сложность живых организмов противоречит этому закону, жизнь возможна, так как все организмы являются открытыми системами, которые обмениваются веществом и энергией с окружающей средой. Таким образом, живые системы не находятся в термодинамическом равновесии, но, вместо этого, выступают диссипативной системой, которая поддерживает своё состояние сложной организованности, вызывая большее увеличение энтропии окружающей средой[101]. В метаболизме клеток это достигается путём сочетания спонтанных процессов катаболизма и неспонтанных процессов анаболизма. В термодинамических условиях метаболизм поддерживает порядок за счёт создания беспорядка[102].
Регуляция и контроль
Подробное рассмотрение темы: Гормоны и Передача сигнала в клетке
Гомеостазом называют постоянство внутренней среды организма. Так как внешняя среда, окружающая большинство организмов, постоянно меняется, для поддержания постоянных условий внутри клеток реакции обмена веществ должны точно регулироваться[103][104]. Регуляция метаболизма позволяет организмам отвечать на сигналы и активно взаимодействовать с окружающей средой[105]. В случае фермента регуляция заключается в повышении и снижении его активности в ответ на сигналы. С другой стороны, фермент оказывает некоторый контроль над метаболическим путём, который определяется как эффект от изменения активности фермента на данный метаболический путь[106].
Влияние инсулина на поглощение глюкозы и обмен веществ. Инсулин связывается со своим рецептором (1), который в свою очередь запускает каскад реакций активации множества белков (2). К ним относятся: транслокация переносчика GLUT4 к плазматической мембране и поступление глюкозы в клетку (3), синтез гликогена (4), гликолиз (5) и синтез жирных кислот (6)
Выделяют несколько уровней регуляции метаболизма. В метаболическом пути происходит саморегуляция на уровне субстрата или продукта; например, уменьшение количества продукта может компенсированно увеличить поток субстрата реакции по данному пути[107]. Этот тип регулирования часто включает аллостерическое регулирование активности некоторых ферментов в метаболических путях[108]. Внешний контроль включает клетку многоклеточного организма, изменяющую свой метаболизм в ответ на сигналы от других клеток. Эти сигналы, как правило, в виде растворимых мессенджеров, например гормоны и факторы роста, определяются специфическими рецепторами на поверхности клеток[109]. Затем эти сигналы передаются внутрь клетки системой вторичных мессенджеров, которые зачастую связаны с фосфорилированием белков[110].
Хорошо изученный пример внешнего контроля — регуляция метаболизма глюкозы инсулином[111]. Инсулин вырабатывается в ответ на повышение уровня глюкозы в крови. Гормон связывается с инсулиновым рецептором на поверхности клетки, затем активируется каскад протеинкиназ, которые обеспечивают поглощение молекул глюкозы клетками и преобразовывают их в молекулы жирных кислот и гликогена[112]. Метаболизм гликогена контролируется активностью фосфорилазы (фермента, который расщепляет гликоген) и гликогенсинтазы (фермента, который образует его). Эти ферменты взаимосвязаны; фосфорилирование ингибируется гликогенсинтазой, но активируется фосфорилазой. Инсулин вызывает синтез гликогена путём активации белковых фосфатаз и уменьшает фосфорилирование этих ферментов[113].
Главные пути метаболизма, описанные выше, например гликолиза и цикла Кребса, присутствуют у всех трёх доменах живых существ и обнаруживаются у последнего универсального общего предка[3][114]. Этот универсальный предок был прокариотом и, вероятно, метаногеном с аминокислотным, нуклеотидным, углеводным метаболизмом[115][116]. Сохранение этих древних метаболических путей в эволюции может быть результатом того, что эти реакции оптимальны для решения конкретных проблем с метаболизмом. Так, конечные продукты гликолиза и цикла Кребса образуются с высокой эффективностью и с минимальным количеством стадий[4][5]. Первые метаболические пути на основе ферментов могли быть частями пуринового метаболизма нуклеотидов с предыдущим метаболических путей и были частью древнего мира РНК[117].
Многие модели были предложены для описания механизмов, посредством которых новые метаболические пути эволюционировали. К ним относятся последовательное добавление новых ферментов на короткий предковый путь, дупликация, а затем дивергенция всех путей, а также набор уже существующих ферментов и их сборка в новый путь реакций[118]. Относительная важность этих механизмов неясна, однако геномные исследования показали, что ферменты в метаболическом пути, скорее всего, имеют общее происхождение, если предполагать, что многие пути эволюционировали шаг за шагом с новыми функциями, созданными из уже существующих этапов пути[119]. Альтернативная модель основана на исследованиях, в которых прослеживается эволюция структуры белков в метаболических связях; предполагают, что ферменты собирались для выполнения схожих функций в различных метаболических путях[120]. Эти процессы сборки привели к эволюционированию ферментативной мозаики[121]. Некоторые части обмена веществ, возможно, существовали в качестве «модулей», которые могли быть повторно использованы в различных путях для выполнения схожих функций[122].
Эволюция также может приводить к потере метаболических функций. Например, у некоторых паразитов метаболические процессы, которые не важны для выживания, утрачены и готовые аминокислоты, нуклеотиды и углеводы получаются от хозяина[123]. Подобные упрощения метаболических возможностей наблюдают у эндосимбиотических организмов[124].
Методы исследования
Подробное рассмотрение темы: Протеомика и Метабономика
Классически, метаболизм изучается упрощённым подходом, который фокусируется на одном метаболическом пути. Особенно ценно использование меченых атомов на организменном, тканевом и клеточном уровнях, которые определяют пути от предшественников до конечных продуктов путём выявления радиоактивно меченых промежуточных продуктов[125]. Ферменты, которые катализируют эти химические реакции, могут затем быть выделены для исследования их кинетики и ответа на ингибиторы. Параллельный подход заключается в выявлении небольших молекул в клетки или ткани; полный набор этих молекул называется метаболом. В целом, эти исследования дают хорошее представление о структуре и функциях простых путей метаболизма, но недостаточны в применении к более сложных системам, например полной метаболизм клетки[126].
Идея сложности метаболических сетей в клетках, которые содержат тысячи различных ферментов, отражена на изображении справа, показывающем взаимодействия только между 43 белками и 40 метаболитами, которые регулируются 45 000 генов[127]. Тем не менее сейчас можно использовать такие данные о геномах для воссоздания полной сети биохимических реакций и образовывать более целостные математические модели, которые могут объяснить и предсказать их поведение[128]. Эти модели особенно сильны, когда используются для интеграции данных о путях и метаболитах, полученных на основе классических методов, с данными по экспрессии генов из протеомных и ДНК-микрочиповых исследований[129]. С помощью этих методов в настоящее время создаётся модель человеческого метаболизма, которая будет служить ориентиром для будущих исследований лекарств и биохимических исследований[130]. Эти модели в настоящее время используются в анализах сети для классификации болезней человека по группам, которые различаются по общим белкам или метаболитам[131][132].
Яркий пример бактериальных метаболических сетей — устройство галстук-бабочки[133][134][135], структура которой позволяет вводить широкий спектр питательных веществ и производить большое разнообразие продуктов и сложных макромолекул, используя сравнительно немного общих промежуточных веществ.
Основная технологическая основа этой информации — метаболическая инженерия. Здесь организмы, например дрожжи, растения или бактерии, генетически модифицируются, чтобы сделать их более эффективными в биотехнологии и помочь в производстве лекарств, например антибиотиков или промышленных химических веществ, таких как 1,3-пропандиола и шикимовой кислоты[136]. Эти генетические модификации обычно направлены на уменьшение количества энергии, используемой для производства продукции, повышения урожайности и снижения производственных отходов[137].
Санторио взвешивает сам себя до и после принятия пищи, из Ars de statica medicina, впервые опубликованной в 1614 году
Термин «метаболизм» впервые введён был в биологию Теодором Шванном в 1840-х годах, однако не получил широкого использования. Термин устоялся в физиологии и проник в большинство языков после издания и перевода руководства по физиологии Фостера в 1870-х годах[138].
История изучения метаболизма охватывает несколько столетий. Исследования начинались с изучения организмов животных, в современной биохимии изучают отдельные метаболические реакции. Понятие обмена веществ впервые встречается в работах Ибн аль-Нафиса (1213—1288), который писал, что «тело и его части находятся в постоянном состоянии распада и питания, так что оно неизбежно претерпевает постоянные изменения»[139]. Первые контролируемые эксперименты по метаболизму у человека были опубликованы Санторио Санторио в 1614 году в книге итал.Ars de statica medicina[140]. Он рассказал, как он сам взвесил себя до и после приёма пищи, сна, работы, секса, натощак, после питья и выделения мочи. Он обнаружил, что большая часть пищи, которую он принял, была утрачена в результате процесса, названного «незаметным испарением».
В ранних исследованиях механизмы метаболических реакций не были обнаружены и считалось, что живой тканью управляет живая сила[141]. В XIX веке при исследовании ферментации сахара спирта дрожжами Луи Пастер сделал вывод, что брожение катализируется веществами из дрожжевых клеток, которые он назвал ферментами. Пастер писал, что «алкогольное брожение — действие, связанное с жизнью и организуется дрожжевыми клетками, не связано со смертью или разложением клеток»[142]. Это открытие, вместе с публикацией Фридриха Вёлера в 1828 году о химическом синтезе мочевины[143], доказало, что органические соединения и химические реакции, обнаруженные в клетках, не имеют различий в принципе, как и любые другие разделы химии.
Открытие ферментов в начале XX века Эдуардом Бухнером разделило изучение метаболических реакций от изучения клеток и дало начало развитию биохимии как науки[144]. Одним из успешных биохимиков начала двадцатого века был Ханс Адольф Кребс, который внёс огромный вклад в изучение метаболизма[145]. Кребс описал цикл мочевины и позднее, работая вместе с Хансом Корнбергом, цикл лимонной кислоты и глиоксилатный цикл[68][146]. В современных биохимических исследованиях широко используют новые методы, такие как хроматография, рентгеноструктурный анализ, ЯМР-спектроскопия, электронная микроскопия и метод классической молекулярной динамики. Эти методы позволяют открывать и подробно изучать множество молекул и метаболических путей в клетках.
↑ 12Ebenhöh O., Heinrich R. Evolutionary optimization of metabolic pathways. Theoretical reconstruction of the stoichiometry of ATP and NADH producing systems (англ.) // Bull Math Biol : journal. — 2001. — Vol. 63, no. 1. — P. 21—55. — doi:10.1006/bulm.2000.0197. — PMID11146883.
↑ 12Meléndez-Hevia E., Waddell T., Cascante M. The puzzle of the Krebs citric acid cycle: assembling the pieces of chemically feasible reactions, and opportunism in the design of metabolic pathways during evolution (англ.) // J Mol Evol[англ.] : journal. — 1996. — Vol. 43, no. 3. — P. 293—303. — doi:10.1007/BF02338838. — PMID8703096.
↑Nomenclature of Lipids (неопр.). IUPAC-IUB Commission on Biochemical Nomenclature (CBN). Дата обращения: 8 марта 2007. Архивировано 22 августа 2011 года.
↑Hegardt F. Mitochondrial 3-hydroxy-3-methylglutaryl-CoA synthase: a control enzyme in ketogenesis (англ.) // Biochem J[англ.] : journal. — 1999. — Vol. 338, no. Pt 3. — P. 569—582. — doi:10.1042/0264-6021:3380569. — PMID10051425. — PMC1220089.
↑Raman R., Raguram S., Venkataraman G., Paulson J., Sasisekharan R. Glycomics: an integrated systems approach to structure-function relationships of glycans (англ.) // Nat Methods : journal. — 2005. — Vol. 2, no. 11. — P. 817—824. — doi:10.1038/nmeth807. — PMID16278650.
↑Sierra S., Kupfer B., Kaiser R. Basics of the virology of HIV-1 and its replication (англ.) // J Clin Virol : journal. — 2005. — Vol. 34, no. 4. — P. 233—244. — doi:10.1016/j.jcv.2005.09.004. — PMID16198625.
↑ 12Wimmer M., Rose I. Mechanisms of enzyme-catalyzed group transfer reactions (англ.) // Annu Rev Biochem[англ.] : journal. — 1978. — Vol. 47. — P. 1031—1078. — doi:10.1146/annurev.bi.47.070178.005123. — PMID354490.
↑Mitchell P. The Ninth Sir Hans Krebs Lecture. Compartmentation and communication in living systems. Ligand conduction: a general catalytic principle in chemical, osmotic and chemiosmotic reaction systems (англ.) // Eur J Biochem[англ.] : journal. — 1979. — Vol. 95, no. 1. — P. 1—20. — doi:10.1111/j.1432-1033.1979.tb12934.x. — PMID378655.
↑ 1234Dimroth P., von Ballmoos C., Meier T. Catalytic and mechanical cycles in F-ATP synthases. Fourth in the Cycles Review Series (англ.) // EMBO Rep[англ.] : journal. — 2006. — March (vol. 7, no. 3). — P. 276—282. — doi:10.1038/sj.embor.7400646. — PMID16607397. — PMC1456893.
↑Coulston, Ann; Kerner, John; Hattner, JoAnn; Srivastava, Ashini. Stanford School of Medicine Nutrition Courses (англ.). — SUMMIT, 2006.
↑Pollak N., Dölle C., Ziegler M. The power to reduce: pyridine nucleotides—small molecules with a multitude of functions (англ.) // Biochem J[англ.] : journal. — 2007. — Vol. 402, no. 2. — P. 205—218. — doi:10.1042/BJ20061638. — PMID17295611. — PMC1798440.
↑ 12Heymsfield S., Waki M., Kehayias J., Lichtman S., Dilmanian F., Kamen Y., Wang J., Pierson R. Chemical and elemental analysis of humans in vivo using improved body composition models (англ.) // American Physiological Society[англ.] : journal. — 1991. — Vol. 261, no. 2 Pt 1. — P. E190—8. — PMID1872381.
↑Dulhunty A. Excitation-contraction coupling from the 1950s into the new millennium (англ.) // Clin Exp Pharmacol Physiol[англ.] : journal. — 2006. — Vol. 33, no. 9. — P. 763—772. — doi:10.1111/j.1440-1681.2006.04441.x. — PMID16922804.
↑Husted S., Mikkelsen B., Jensen J., Nielsen N. Elemental fingerprint analysis of barley (Hordeum vulgare) using inductively coupled plasma mass spectrometry, isotope-ratio mass spectrometry, and multivariate statistics (англ.) // Anal Bioanal Chem[англ.] : journal. — 2004. — Vol. 378, no. 1. — P. 171—182. — doi:10.1007/s00216-003-2219-0. — PMID14551660.
↑Finney L., O'Halloran T. Transition metal speciation in the cell: insights from the chemistry of metal ion receptors (англ.) // Science : journal. — 2003. — Vol. 300, no. 5621. — P. 931—936. — doi:10.1126/science.1085049. — PMID12738850.
↑Dunn L., Rahmanto Y., Richardson D. Iron uptake and metabolism in the new millennium (англ.) // Trends Cell Biol[англ.] : journal. — 2007. — Vol. 17, no. 2. — P. 93—100. — doi:10.1016/j.tcb.2006.12.003. — PMID17194590.
↑Микробиология: учебник для студ. высш. учеб. заведений / А. И. Нетрусов, И. Б. Котова — М.: Издательский центр «Академия», 2006. — 352 с. ISBN 5-7695-2583-5
↑Микробиология: учебник для студ. биол. специальностей вузов / М. В. Гусев, Л. А. Минеева — 4-е изд., стер. — М.: Издательский центр «Академия», 2003. — 464 с. ISBN 5-7695-1403-5
↑Пиневич А.В. Микробиология. Биология прокариотов: Учебник. В 3 т. Том 2.. — СПб.: Изд-во С.-Петерб. ун-та, 2007. — 331 с. с. — ISBN ISBN 978-5-288-04269-0 (т.II)ISBN 5-288-04056-7.
↑Capaldi R., Aggeler R. Mechanism of the F(1)F(0)-type ATP synthase, a biological rotary motor (англ.) // Trends Biochem Sci[англ.] : journal. — 2002. — Vol. 27, no. 3. — P. 154—160. — doi:10.1016/S0968-0004(01)02051-5. — PMID11893513.
↑Weber K., Achenbach L., Coates J. Microorganisms pumping iron: anaerobic microbial iron oxidation and reduction (англ.) // Nat Rev Microbiol : journal. — 2006. — Vol. 4, no. 10. — P. 752—764. — doi:10.1038/nrmicro1490. — PMID16980937.
↑Jetten M., Strous M., van de Pas-Schoonen K., Schalk J., van Dongen U., van de Graaf A., Logemann S., Muyzer G., van Loosdrecht M., Kuenen J. The anaerobic oxidation of ammonium (неопр.) // FEMS Microbiol Rev. — 1998. — Т. 22, № 5. — С. 421—437. — doi:10.1111/j.1574-6976.1998.tb00379.x. — PMID9990725.
↑Simon J. Enzymology and bioenergetics of respiratory nitrite ammonification (англ.) // FEMS Microbiol Rev : journal. — 2002. — Vol. 26, no. 3. — P. 285—309. — doi:10.1111/j.1574-6976.2002.tb00616.x. — PMID12165429.
↑Munekage Y., Hashimoto M., Miyake C., Tomizawa K., Endo T., Tasaka M., Shikanai T. Cyclic electron flow around photosystem I is essential for photosynthesis (англ.) // Nature : journal. — 2004. — Vol. 429, no. 6991. — P. 579—582. — doi:10.1038/nature02598. — PMID15175756.
↑Boiteux A., Hess B. Design of glycolysis (англ.) // Philos Trans R Soc Lond B Biol Sci : journal. — 1981. — Vol. 293, no. 1063. — P. 5—22. — doi:10.1098/rstb.1981.0056. — PMID6115423.
↑Pilkis S., el-Maghrabi M., Claus T. Fructose-2,6-bisphosphate in control of hepatic gluconeogenesis. From metabolites to molecular genetics (англ.) // Diabetes Care[англ.] : journal. — 1990. — Vol. 13, no. 6. — P. 582—599. — doi:10.2337/diacare.13.6.582. — PMID2162755.
↑ 12Ensign S. Revisiting the glyoxylate cycle: alternate pathways for microbial acetate assimilation (англ.) // Microbiology[англ.] : journal. — Microbiology Society[англ.], 2006. — Vol. 61, no. 2. — P. 274—276. — doi:10.1111/j.1365-2958.2006.05247.x. — PMID16856935.
↑Finn P., Dice J. Proteolytic and lipolytic responses to starvation (неопр.) // Nutrition. — 2006. — Т. 22, № 7—8. — С. 830—844. — doi:10.1016/j.nut.2006.04.008. — PMID16815497.
↑ 12Kornberg H., Krebs H. Synthesis of cell constituents from C2-units by a modified tricarboxylic acid cycle (англ.) // Nature : journal. — 1957. — Vol. 179, no. 4568. — P. 988—991. — doi:10.1038/179988a0. — PMID13430766.
↑Hudson H. Freeze, Gerald W. Hart, Ronald L. Schnaar.Glycosylation Precursors // Essentials of Glycobiology / Ajit Varki, Richard D. Cummings, Jeffrey D. Esko, Pamela Stanley, Gerald W. Hart, Markus Aebi, Alan G. Darvill, Taroh Kinoshita, Nicolle H. Packer, James H. Prestegard, Ronald L. Schnaar, Peter H. Seeberger. — Cold Spring Harbor (NY): Cold Spring Harbor Laboratory Press, 2015. Архивировано 24 февраля 2022 года.
↑ 12Kuzuyama T., Seto H. Diversity of the biosynthesis of the isoprene units (англ.) // Nat Prod Rep[англ.] : journal. — 2003. — Vol. 20, no. 2. — P. 171—183. — doi:10.1039/b109860h. — PMID12735695.
↑Lengyel P., Söll D. Mechanism of protein biosynthesis (англ.) // Microbiology and Molecular Biology Reviews[англ.] : journal. — American Society for Microbiology[англ.], 1969. — Vol. 33, no. 2. — P. 264—301. — PMID4896351. — PMC378322.
↑ 12Rudolph F. The biochemistry and physiology of nucleotides (англ.) // J Nutr[англ.] : journal. — 1994. — Vol. 124, no. 1 Suppl. — P. 124S—127S. — PMID8283301.
↑Stasolla C., Katahira R., Thorpe T., Ashihara H. Purine and pyrimidine nucleotide metabolism in higher plants (англ.) // Plant Physiology : journal. — American Society of Plant Biologists, 2003. — Vol. 160, no. 11. — P. 1271—1295. — doi:10.1078/0176-1617-01169. — PMID14658380.
↑Smith J. Enzymes of nucleotide synthesis (неопр.) // Curr Opin Struct Biol. — 1995. — Т. 5, № 6. — С. 752—757. — doi:10.1016/0959-440X(95)80007-7. — PMID8749362.
↑Testa B., Krämer S. The biochemistry of drug metabolism—an introduction: part 1. Principles and overview (англ.) // Chem Biodivers : journal. — 2006. — Vol. 3, no. 10. — P. 1053—1101. — doi:10.1002/cbdv.200690111. — PMID17193224.
↑Danielson P. The cytochrome P450 superfamily: biochemistry, evolution and drug metabolism in humans (англ.) // Curr Drug Metab[англ.] : journal. — 2002. — Vol. 3, no. 6. — P. 561—597. — doi:10.2174/1389200023337054. — PMID12369887.
↑King C., Rios G., Green M., Tephly T. UDP-glucuronosyltransferases (англ.) // Curr Drug Metab[англ.] : journal. — 2000. — Vol. 1, no. 2. — P. 143—161. — doi:10.2174/1389200003339171. — PMID11465080.
↑Galvão T., Mohn W., de Lorenzo V. Exploring the microbial biodegradation and biotransformation gene pool (англ.) // Trends Biotechnol[англ.] : journal. — 2005. — Vol. 23, no. 10. — P. 497—506. — doi:10.1016/j.tibtech.2005.08.002. — PMID16125262.
↑Janssen D., Dinkla I., Poelarends G., Terpstra P. Bacterial degradation of xenobiotic compounds: evolution and distribution of novel enzyme activities (англ.) // Environ Microbiol : journal. — 2005. — Vol. 7, no. 12. — P. 1868—1882. — doi:10.1111/j.1462-2920.2005.00966.x. — PMID16309386.
↑Davies K. Oxidative stress: the paradox of aerobic life (неопр.) // Biochem Soc Symp. — 1995. — Т. 61. — С. 1—31. — PMID8660387.
↑Soyer O., Salathé M., Bonhoeffer S. Signal transduction networks: topology, response and biochemical processes (англ.) // J Theor Biol[англ.] : journal. — 2006. — Vol. 238, no. 2. — P. 416—425. — doi:10.1016/j.jtbi.2005.05.030. — PMID16045939.
↑Westerhoff H., Groen A., Wanders R. Modern theories of metabolic control and their applications (review) (англ.) // Biosci Rep : journal. — 1984. — Vol. 4, no. 1. — P. 1—22. — doi:10.1007/BF01120819. — PMID6365197.
↑Salter M., Knowles R., Pogson C. Metabolic control (неопр.) // Essays Biochem. — 1994. — Т. 28. — С. 1—12. — PMID7925313.
↑Fell D., Thomas S. Physiological control of metabolic flux: the requirement for multisite modulation (англ.) // Biochem J[англ.] : journal. — 1995. — Vol. 311, no. Pt 1. — P. 35—9. — PMID7575476. — PMC1136115.
↑Cohen P. The regulation of protein function by multisite phosphorylation—a 25 year update (англ.) // Trends Biochem Sci[англ.] : journal. — 2000. — Vol. 25, no. 12. — P. 596—601. — doi:10.1016/S0968-0004(00)01712-6. — PMID11116185.
↑Romano A., Conway T. Evolution of carbohydrate metabolic pathways (неопр.) // Res Microbiol. — 1996. — Т. 147, № 6—7. — С. 448—455. — doi:10.1016/0923-2508(96)83998-2. — PMID9084754.
↑Schmidt S., Sunyaev S., Bork P., Dandekar T. Metabolites: a helping hand for pathway evolution? (англ.) // Trends Biochem Sci[англ.] : journal. — 2003. — Vol. 28, no. 6. — P. 336—341. — doi:10.1016/S0968-0004(03)00114-2. — PMID12826406.
↑Light S., Kraulis P. Network analysis of metabolic enzyme evolution in Escherichia coli (англ.) // BMC Bioinformatics[англ.] : journal. — 2004. — Vol. 5. — P. 15. — doi:10.1186/1471-2105-5-15. — PMID15113413. — PMC394313.Alves R., Chaleil R., Sternberg M. Evolution of enzymes in metabolism: a network perspective (англ.) // J Mol Biol[англ.] : journal. — 2002. — Vol. 320, no. 4. — P. 751—770. — doi:10.1016/S0022-2836(02)00546-6. — PMID12095253.
↑Kim H. S., Mittenthal J. E., Caetano-Anolles G. MANET: tracing evolution of protein architecture in metabolic networks (англ.) // BMC Bioinformatics[англ.] : journal. — 2006. — Vol. 19, no. 7. — P. 351. — doi:10.1186/1471-2105-7-351. — PMID16854231. — PMC1559654.
↑Teichmann S. A., Rison S. C., Thornton J. M., Riley M., Gough J., Chothia C. Small-molecule metabolsim: an enzyme mosaic (англ.) // Trends Biotechnol[англ.] : journal. — 2001. — Vol. 19, no. 12. — P. 482—486. — doi:10.1016/S0167-7799(01)01813-3. — PMID11711174.
↑Lawrence J. Common themes in the genome strategies of pathogens (англ.) // Curr Opin Genet Dev : journal. — 2005. — Vol. 15, no. 6. — P. 584—588. — doi:10.1016/j.gde.2005.09.007. — PMID16188434.Wernegreen J. For better or worse: genomic consequences of intracellular mutualism and parasitism (англ.) // Curr Opin Genet Dev : journal. — 2005. — Vol. 15, no. 6. — P. 572—583. — doi:10.1016/j.gde.2005.09.013. — PMID16230003.
↑Pál C., Papp B., Lercher M., Csermely P., Oliver S., Hurst L. Chance and necessity in the evolution of minimal metabolic networks (англ.) // Nature : journal. — 2006. — Vol. 440, no. 7084. — P. 667—670. — doi:10.1038/nature04568. — PMID16572170.
↑Phair R. Development of kinetic models in the nonlinear world of molecular cell biology (англ.) // Metabolism : journal. — 1997. — Vol. 46, no. 12. — P. 1489—1495. — doi:10.1016/S0026-0495(97)90154-2. — PMID9439549.
↑Sterck L., Rombauts S., Vandepoele K., Rouzé P., Van de Peer Y. How many genes are there in plants (... and why are they there)? (англ.) // Curr Opin Plant Biol : journal. — 2007. — Vol. 10, no. 2. — P. 199—203. — doi:10.1016/j.pbi.2007.01.004. — PMID17289424.
↑Borodina I., Nielsen J. From genomes to in silico cells via metabolic networks (англ.) // Curr Opin Biotechnol : journal. — 2005. — Vol. 16, no. 3. — P. 350—355. — doi:10.1016/j.copbio.2005.04.008. — PMID15961036.
↑Gianchandani E., Brautigan D., Papin J. Systems analyses characterize integrated functions of biochemical networks (англ.) // Trends Biochem Sci[англ.] : journal. — 2006. — Vol. 31, no. 5. — P. 284—291. — doi:10.1016/j.tibs.2006.03.007. — PMID16616498.
↑Ma H. W., Zeng A. P. The connectivity structure, giant strong component and centrality of metabolic networks (англ.) // Bioinformatics : journal. — 2003. — Vol. 19, no. 11. — P. 1423—1430. — doi:10.1093/bioinformatics/btg177. — PMID12874056.
↑Zhao J., Yu H., Luo J. H., Cao Z. W., Li Y. X. Hierarchical modularity of nested bow-ties in metabolic networks (англ.) // BMC Bioinformatics[англ.] : journal. — 2006. — Vol. 7. — P. 386. — doi:10.1186/1471-2105-7-386. — PMID16916470. — PMC1560398.
↑Thykaer J., Nielsen J. Metabolic engineering of beta-lactam production (англ.) // Metab Eng[англ.] : journal. — 2003. — Vol. 5, no. 1. — P. 56—69. — doi:10.1016/S1096-7176(03)00003-X. — PMID12749845.González-Pajuelo M., Meynial-Salles I., Mendes F., Andrade J., Vasconcelos I., Soucaille P. Metabolic engineering of Clostridium acetobutylicum for the industrial production of 1,3-propanediol from glycerol (англ.) // Metab Eng[англ.] : journal. — 2005. — Vol. 7, no. 5—6. — P. 329—336. — doi:10.1016/j.ymben.2005.06.001. — PMID16095939.Krämer M., Bongaerts J., Bovenberg R., Kremer S., Müller U., Orf S., Wubbolts M., Raeven L. Metabolic engineering for microbial production of shikimic acid (англ.) // Metab Eng[англ.] : journal. — 2003. — Vol. 5, no. 4. — P. 277—283. — doi:10.1016/j.ymben.2003.09.001. — PMID14642355.
↑Фельдман Г.Э. Майкл Фостер. — Ленинград: Наука, 1986. — С. 52.
↑Dr. Abu Shadi Al-Roubi (1982), «Ibn Al-Nafis as a philosopher», Symposium on Ibn al Nafis, Second International Conference on Islamic Medicine: Islamic Medical Organization, Kuwait (cf. Ibnul-Nafees As a Philosopher, Encyclopedia of Islamic World[1]).
↑Eknoyan G. Santorio Sanctorius (1561—1636) — founding father of metabolic balance studies (англ.) // Am J Nephrol : journal. — 1999. — Vol. 19, no. 2. — P. 226—233. — doi:10.1159/000013455. — PMID10213823.
↑Dubos J. Louis Pasteur: Free Lance of Science, Gollancz. Quoted in Manchester K. L. (1995) Louis Pasteur (1822–1895)—chance and the prepared mind (англ.) // Trends Biotechnol[англ.] : journal. — 1951. — Vol. 13, no. 12. — P. 511—515. — doi:10.1016/S0167-7799(00)89014-9. — PMID8595136.
↑Kinne-Saffran E., Kinne R. Vitalism and synthesis of urea. From Friedrich Wöhler to Hans A. Krebs (англ.) // Am J Nephrol : journal. — 1999. — Vol. 19, no. 2. — P. 290—294. — doi:10.1159/000013463. — PMID10213830.
↑Krebs H. A., Henseleit K. Untersuchungen über die Harnstoffbildung im tierkorper (нем.) // Z. Physiol. Chem.[англ.] : magazin. — 1932. — Bd. 210. — S. 33—66.Krebs H., Johnson W. Metabolism of ketonic acids in animal tissues (англ.) // Biochem J[англ.] : journal. — 1937. — April (vol. 31, no. 4). — P. 645—660. — PMID16746382. — PMC1266984.
Дезоксирибонуклеи́новая кислота́ (ДНК) — макромолекула, обеспечивающая хранение, передачу из поколения в поколение и реализацию генетической программы развития и функционирования организмов. Молекула ДНК хранит биологическую информацию в виде генетического кода, состоящего из последовательности нуклеотидов. ДНК содержит информацию о структуре различных видов РНК и белков.
Биохи́мия — наука о химическом составе живых клеток и организмов, а также о лежащих в основе их жизнедеятельности химических процессах. Термин «биохимия» эпизодически употреблялся с середины XIX века, в классическом смысле он был предложен и введён в научную среду в 1903 году немецким химиком Карлом Нейбергом.
Рибонуклеи́новая кислота́ (РНК) — одна из трёх основных макромолекул, которые содержатся в клетках всех живых организмов и играют важную роль в кодировании, прочтении, регуляции и экспрессии генов.
Белки́ — высокомолекулярные органические вещества, состоящие из альфа-аминокислот, соединённых в цепочку пептидной связью. В живых организмах аминокислотный состав белков определяется генетическим кодом, при синтезе в большинстве случаев используется 20 стандартных аминокислот. Множество их комбинаций определяет большое разнообразие свойств молекул белков. Кроме того, аминокислотные остатки в составе белка часто подвергаются посттрансляционным модификациям, которые могут возникать и до того, как белок начинает выполнять свою функцию, и во время его «работы» в клетке. Часто в живых организмах несколько молекул разных белков образуют сложные комплексы, например фотосинтетический комплекс и другие комплексы.
Фенилалани́н — ароматическая альфа-аминокислота. Существует в двух оптически изомерных формах l и d и в виде рацемата. По химическому строению соединение можно представить как аминокислоту аланин, в которой один из атомов водорода замещён фенильной группой.
Цитозо́ль — жидкое содержимое клетки. Большую часть цитозоля занимает внутриклеточная жидкость. Цитозоль разбивается на компартменты при помощи разнообразных мембран. У эукариот цитозоль располагается под плазматической мембраной и является частью цитоплазмы, в которую, помимо цитозоля, входят митохондрии, пластиды и другие органеллы, но не содержащаяся в них жидкость и внутренние структуры. Таким образом, цитозоль представляет собой жидкий матрикс, окружающий органеллы. У прокариот большая часть химических реакций метаболизма происходит в цитозоле, и лишь небольшая их часть происходит в мембранах и периплазматическом пространстве. У эукариот, хотя многие реакции протекают в органеллах, некоторые реакции, например, гликолиз или синтез жирных кислот, происходят в цитозоле.
Архе́и — домен живых организмов. Археи представляют собой одноклеточные микроорганизмы, не имеющие ядра, а также каких-либо мембранных органелл.
Никотинамидадениндинуклеоти́д — кофермент, имеющийся во всех живых клетках. NAD представляет собой динуклеотид и состоит из двух нуклеотидов, соединённых своими фосфатными группами. Один из нуклеотидов в качестве азотистого основания содержит аденин, другой — никотинамид.
Окисли́тельное фосфорили́рование — метаболический путь, при котором энергия, образовавшаяся при окислении питательных веществ, запасается в митохондриях клеток в виде АТФ. Хотя различные формы жизни на Земле используют разные питательные вещества, АТФ является универсальным соединением, в котором запасается энергия, необходимая для других метаболических процессов. Почти все аэробные организмы осуществляют окислительное фосфорилирование. Вероятно, широкому распространению этого метаболического пути способствовала его высокая энергетическая эффективность по сравнению с анаэробным брожением.
Убиквити́н — небольшой консервативный белок эукариот, участвующий в регуляции процессов внутриклеточной деградации других белков, а также в модификации их функций. Он присутствует почти во всех тканях многоклеточных эукариот, а также у одноклеточных эукариотических организмов. Убиквитин был открыт в 1975 году Гидеоном Голдштейном с соавторами и охарактеризован в 70—80-х годах XX века. В геноме человека есть четыре гена, кодирующих убиквитин: UBB, UBC, UBA52 и RPS27A.
Шикима́тный путь — метаболический путь, промежуточным метаболитом которого является шикимовая кислота (шикимат). Шикиматный путь отмечается как специализированный путь биосинтеза бензоидных ароматических соединений. Шикиматным путём в природе синтезируются такие известные соединения, как фенилаланин, тирозин, триптофан, бензоат, салицилат. Шикиматный путь является источником ароматических предшественников терпеноидных хинонов, токоферолов, фолата, флавоноидов, лигнинов, суберинов, меланинов, таннинов и огромного множества других соединений, играющих в природе различную роль. Шикиматный путь — один из древнейших эволюционно консервативных метаболических путей; как источник фундаментальных составляющих живой материи фактически представляет собой часть первичного метаболизма. Значение шикиматного пути велико, так как этот путь является единственным установленным путём биосинтеза ряда важнейших природных соединений, в том числе значимых в плане их практического использования. В строгом смысле под шикиматным путём следует понимать только общую часть разветвлённого метаболического процесса — путь биосинтеза хоризмовой кислоты (хоризмата).
Аминокислоты с разветвлёнными боковыми цепями — группа протеиногенных аминокислот, характеризующихся разветвлёнными строением алифатической боковой цепи. К таким аминокислотам относятся лейцин, изолейцин и валин.
Гипотеза мира сульфидов железа — гипотетический этап возникновения жизни на Земле и ранней эволюции, предложенный Гюнтером Вэхтерсхойзером, юристом из Мюнхена, имеющим также научную степень по химии. Опубликовал свои идеи при поддержке философа Карла Р. Поппера. Как следует из названия теории, она предполагает, что жизнь могла зародиться на поверхности кристаллов сульфидов железа.
Триптофановый репрессор является фактором транскрипции, участвует в управлении метаболизмом аминокислот. Его лучше всего изучать на примере кишечной палочки, где он является димерным белком, который регулирует транскрипцию 5 генов в триптофановом опероне. Когда аминокислота триптофан в изобилии в клетке, он связывается с белком, что вызывает конформационные изменения в белке Репрессорный комплекс затем связывается со своей операторной последовательностью в генах, которую он регулирует, отключая гены..
Непротеиногенные аминокислоты — аминокислоты, которые не участвуют в биосинтезе белка. Непротеиногенные аминокислоты часто не входят в состав непатогенных и «нормальных» белков человека, не следует их путать с нестандартными аминокислотами, которые образуются непосредственно при синтезе белка или в результате различных ковалентных модификаций молекулярных структур протеиногенных (кодируемых) аминокислот в составе белков, например, гидроксипролина, десмозина или аллизина. Однако они могут входить в состав некоторых небольших молекул или пептидов, синтез которых является нерибосомальным. Химически синтезированные аминокислоты можно назвать неприродными аминокислотами. Многие из них являются токсинами и ингибиторами ферментов разнообразных метаболических реакций. Известно свыше 400 природных аминокислот и, возможно, больше тысячи их комбинаций. Неприродные аминокислоты могут быть синтетически получены из их нативных аналогов с помощью модификаций, таких как алкилирование аминогруппы, замещение боковой цепи, циклизация за счёт удлинения структурных связей и изостерических замен в аминокислотном остове.
Фосфоенолпируваткарбоксилаза представляет собой фермент из семейства карбоксилаз, который встречается у растений и некоторых бактерий. Он катализирует присоединение бикарбоната (НСО3−) к фосфоенолпирувату (ФЕП) с образованием четырёх углеродного соединения оксалоацетата и неорганического фосфата:
ФЕП + НСО3− → оксалоацетат + Фн
Ацил-КоА-дегидрогеназы, также ACADs — семейство ферментов из класса оксидоредуктаз, которые катализируют реакции переноса протона (дегидрогенизация) от субстрата — ацил-КоА жирной кислоты на электрон-переносящий флавопротеин (FAD), участвуют в процессе β-окисления. Результатом реакции является образование двойной связи расположенной между атомами С2 (α) и С3 (β) в молекуле тиоэфира субстрата (ацил-КоА).
Карнитин-пальмитоилтрансфераза I, также карнитинацилтрансфераза I, карнитинацил-СоА-трансфераза I или пальмитоил-CoA-трансфераза I — митохондриальный фермент, одна из форм карнитин-О-пальмитоилтрансферазы, относится к семейству ацилтрансфераз. Катализирует перенос ацильной группы (—COR) от молекулы ацил-CoA жирных кислот с длинной углеводородной цепью на молекулу карнитина, с образованием ацилкарнитина и свободной молекулы кофермента А. Часто продуктом реакции является пальмитоилкарнитин, однако, в качестве субстрата могут выступать и другие остатки жирных кислот. Один из нескольких ферментов карнитиновой транспортной системы. Ген, кодирующий данный фермент локализуется в 11-й хромосоме — CPT1AАрхивная копия от 10 сентября 2016 на Wayback Machine.
Митохондриа́льный ма́трикс или просто ма́трикс — ограниченное внутренней мембраной пространство, расположенное внутри митохондрий. Слово «матрикс» происходит из того, что эта среда является намного более вязкой по сравнению с более водянистой цитоплазмой. В состав матрикса входит множество веществ, включая ферменты, митохондриальную ДНК (кольцевая), рибосомы, малые органические молекулы, нуклеотидные коферменты и неорганические ионы. Ферменты матрикса содействуют реакциям биохимических процессов, в ходе которых синтезируется АТФ, таких как цикл трикарбоновых кислот, окислительное фосфорилирование, окисление пирувата и бета-окисление жирных кислот.
Ферредоксин-НАДФ+-редуктаза, сокращенно ФНР, фермент из класса оксидоредуктаз, катализирующий реакцию восстановления НАДФ+, используя в качестве донора электронов ферредоксин.
Эта страница основана на статье Википедии. Текст доступен на условиях лицензии CC BY-SA 4.0; могут применяться дополнительные условия. Изображения, видео и звуки доступны по их собственным лицензиям.