Орексин
| Prepro-orexin | |
|---|---|
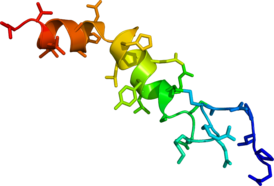 Solution phase NMR structure of orexin A based on the PDB coordinates 1R02 | |
| Идентификаторы | |
| Символ | Orexin |
| Pfam | PF02072 |
| InterPro | IPR001704 |
| SCOP | 1cq0 |
| SUPERFAMILY | 1cq0 |
| OPM superfamily | 154 |
| OPM protein | 1wso |
| Доступные структуры белков | |
| Pfam | структуры |
| PDB | RCSB PDB; PDBe; PDBj |
| PDBsum | 3D-модель |
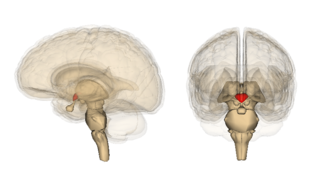
Орексины или гипокретины (англ. Orexins/hypocretins) — название двух нейропептидов (белковых нейромедиаторов), независимо обнаруженных двумя группами исследователей в 1998 году[1]. Два орексина, Орексин А и Орексин Б, имеют 50-процентную гомологию последовательности, не имея при этом значительной гомологии с остальными пептидами в организме. Орексины синтезируются сравнительно небольшой популяцией клеток латерального гипоталамуса (около 50000[1]), чьи аксоны достигают практически всех регионов мозга. Считается, что недостаток орексинов в мозге приводит к развитию нарколепсии. Орексины демонстрируют сильно сохранившуюся последовательность аминокислот и найдены у всех позвоночных, что указывает на их раннее появление в эволюции позвоночных.
Открытие и название
Группа Masashi Yanagisawa и T. Sakurai из Юго-Западного Медицинского Центра Техасского Университета в Далласе проводила в 1998 г. поиск неизвестных науке гормонов, используя метод «обратной фармакологии». В нашем геноме известно около 100—150 последовательностей, предположительно кодирующих G-белок-связанные клеточные рецепторы, чья роль в организме не выявлена (рецепторы без известных лигандов, или «рецепторы-сироты»). Внедрив эти последовательности в колонии клеток путём генной инженерии, можно «отлавливать» с их помощью лиганды из экстрактов тканей. «Обратным фармакологическим» этот метод называют потому, что, в отличие от обычных исследований, первым находится не лиганд, а рецептор. При своей перспективности этот метод менее надёжный и более трудоёмкий, чем обычные[2].
Обнаружив два новых лиганда, и установив, что они влияют на аппетит (это предположение было выдвинуто в самом начале, так как латеральный гипоталамус тесно связан с регулировкой аппетита), они предложили для них название «орексины», производное от греческого ορεξις-аппетит.
Группа Luis de Lecea и Thomas Kilduff открыла в том же году два новых пептида при изоляции цепочек мРНК в гипоталамусе, которые они назвали «гипокретинами» из-за структурного подобия с гормоном секретином («гипо»- от гипоталамуса, «кретин»- от секретина).
Консенсус касательно их названия в научном обществе до сих пор не достигнут: некоторые отвергают название «орексины», указывая на то, что стимуляция аппетита не является их основной ролью, к тому же она может быть косвенной; их противники же считают, что название «гипокретин» является ещё более странным и менее информативным, тем более что названия многих нейропептидов не отражают их основную функцию.
При этом, следует знать, что ορεξις не самое точное слово в греческом для обозначения аппетита, его смысл более общий — «желание», «стремление», «страсть» (более близкий перевод слова «аппетит» на греческий βουλεμια, от которого происходит название болезни булимия). При этом, как указывает один автор, это говорит в поддержку названия «орексин», так как последние исследования обнаружили важную роль орексинов в мотивированном поведении (reward circuit)[3].
Биохимия
Оба орексина синтезируются из общего прекурсора (препроорексина, ППО), разрезаемого на две части. ППО имеет типичную секреторную последовательность на N-терминальном конце.
Орексин А длиной в 33 аминокислоты и имеет две дисульфидные связи (Cys6-Cys12, Cys7-Cys14), необходимые для его функционирования. Орексин Б — прямая цепочка из 28 аминокислот. Орексин А имеет примерно одинаковою аффинность к обоим рецепторам, в то время как Орексин Б реагирует в основном с OX2 и в 5 раз слабее Орексина А[2].
Анатомия
Гипоталамус расположен ниже таламуса, над хиазмой зрительных нервов. Проекции орексинных аксонов и орексинные рецепторы находятся в следующих областях мозга:
- Моноаминные нейроны:
- Голубое пятно, (лат. locus caeruleus, LC) — выделяет норадреналин
- Ядра сосцевидного тела, (nuclei corporis mamillaris medialis et lateralis, tubero-mammillary nucleus - TMN) — выделяют гистамин
- Дорзальные ядра шва (Dorsal raphe nucleus - DR) — выделяют серотонин, связаны с синдромом катаплексии
- Дугообразное ядро (Arcuate nucleus) — связан с регулировкой аппетита, выработки гормонов и сексуального поведения
- Задние обонятельные центры (Tenia tecta) — участвуют в поддержании постоянства внутренней среды организма, регуляции вегетативных функций и формировании эмоций и мотиваций.
- Гиппокамп — участвует в механизмах формирования эмоций, консолидации памяти
- Паравентрикулярное ядро гипоталамуса (PVN)
- Латеральная область гипоталамуса.
Функции
Сон
Основной функцией орексинов является поддержание состояния бодрствования. Эндогенная нехватка орексинов приводит к нарколепсии — заболеванию, характеризующемуся нарушениями циклов сна-бодрствования[1][4].
Орексины сохраняют баланс между нейронами, производящими моноамины и вентролатеральным преоптическим ядром (VLPO). В состоянии бодрствования орексинные нейроны посылают возбуждающие (экситаторные) сигналы моноаминным нейронам, а те посылают обратно (осуществляя обратную связь) тормозящие (ингибиторные) сигналы малой мощности. Моноаминные нейроны возбуждают таламус и кору головного мозга и тем самым поддерживают бодрствование. При понижении уровня экситаторных сигналов, поступающих в орексинные нейроны из районов мозга, связанных с циркадными ритмами и балансом энергии, VLPO и моноаминные нейроны тормозят орексинные нейроны, уровень моноаминов падает и организм погружается в сон. В состоянии сна происходит постоянная ингибиция орексинных нейронов центром VLPO.
В отсутствие орексинов в мозге теряется важное звено гомеостатической системы: между моноаминными нейронами и VLPO существует постоянная взаимная ингибиция (нет порогового уровня сигналов, активация одного из центров полностью подавляет второй), поэтому плавность и цикличность переходов от одного состояния к другому исчезает, сменяясь на неожиданные мгновенные переходы (flip-flop) от одного состояния к другому.
Некоторые учёные считают, что основной эффект орексинов реализуется повышением уровня гистамина в ЦНС через OX2.
Регулировка энергетического баланса
При наблюдении за людьми и животными, страдающими нарколепсией, была отмечена (не сильно выраженная) склонность к полноте и ожирению, хотя аппетит при этом снижался. Опыты на мышах позволили учёным сделать вывод, что орексины повышают одновременно и аппетит, и базальный уровень метаболизма.
Установлено, что орексинные нейроны реагируют на уровень глюкозы в крови, лептин (оба имеют ингибиторный эффект) и другие нейротрансмиттеры и физиологические факторы, связанные с энергетическим балансом в организме[5].
Считается, что орексин действует через NPY (нейропептид Y), а также через подавление POMC (ПОМК)-нейронов.
Эффекты в периферийных тканях
Орексины имеют значение и в периферийных тканях. Известно влияние орексинов на уровень сахара в крови, метаболизм в жировых тканях, кровяное давление (повышает) и овуляцию[6].
В тонкой кишке многих млекопитающих были найдены как орексинные рецепторы, так и препроорексин (ППО), орексины А и Б. В желудке морских свинок орексин А был обнаружен в клетках, выделяющих гастрин. Микроинъекции орексина значительно усиливали кишечную подвижность в опытах с мышами. Кроме того, орексины приводят к усиленному выделению желудочной кислоты независимо от гастрина. У сытых крыс наблюдался также эффект усиленного выделения бикарбоната натрия в двенадцатиперстной кишке.
Орексинные нейроны находятся и в поджелудочной железе, где они выделяют орексин в случае понижения концентрации глюкозы. Орексин влияет и на экзокринную функцию поджелудочной железы.
ППО и орексины А и Б были обнаружены и в корковом веществе надпочечников, где они (независимо от центральных механизмов) стимулируют выделение кортикостероидов. Согласно нынешним данным, в надпочечниках практически отсутствует рецептор OX1. Высокие уровни OX2 были найдены в феохромоцитомах.
В репродуктивной системе человека орексин, по-видимому, у мужчин регулирует экспрессию генов в клетках Сертоли, у женщин наблюдаются цикличные колебания выделения орексина, соответствующие менструальному циклу.
Небольшие количества орексинов были найдены и в крови, но значение этого факта остаётся неизвестным.
Участие в процессах развития зависимости и привыкания
Орексины занимают центральное место в мезолимбическом и мезокортикальном путях (цепочках) в мозге, отвечающие за выработку мотивированного поведения и появлении психологической зависимости. Одним из самых распространённых способов лечения нарколепсии является приём психостимуляторов, таких как амфетамины, метилфенидат (в ряде стран запрещён), модафинил и лекарства из группы СИОЗН. Несмотря на то, что психостимуляторы (кроме модафинила) вызывают зависимость у большинства населения, это не наблюдается у больных нарколепсией, отсюда был сделан вывод о ключевой роли орексинов в становлении психофизической зависимости.
Известно о взаимодействии орексинных нейронов с регионом VTA (ventral tegmental area), которое делает возможным нейронную пластичность, связанную с процессом аддикции. Активация этой системы может возобновить поведение, направленное на получение удовольствия от наркотика через долгое время после прекращения его приёма[1].
Рецепторы
Существуют два орексиновых рецептора — OX1 and OX2. Они являются рецепторами, сопряженными с G белком, (G protein-coupled receptors, GPCRs). У двух рецепторов наблюдается 64-процентная идентичность в последовательности аминокислот[2].
Последние исследования показывают, что засыпание связано в основном с блокировкой рецептора OX2, а фаза быстрого сна связана с OX1[1].
Рецепторы в ЦНС
Рецепторы OX1 находятся в основном в LC, dorsal raphe, tenia tecta, hyppocampal formation.
Рецепторы OX2 находятся в основном в TMN, DMH (в гипоталамусе), PVN, LHA (в гипоталамусе)[1].
Рецепторы на периферии
Фармакологический потенциал
GPCRs рецепторы являются мишенью до 40 % выпускаемых лекарств, поэтому с открытием орексинов появилась надежда, что будут разработаны новые лекарства против различных расстройств сна, эффективные и обладающие немногими побочными эффектами.
Проводились опыты по пересадке крысам орексинных нейронов, что может служить основой для хирургического лечения нарколепсии.
Также проводились эксперименты, в ходе которых с помощью вмешательства в систему орексинов пытались бороться с наркотической зависимостью.
Сегодня разрабатываются как орексинные агонисты (для лечения нарколепсии), так и антагонисты (для лечения бессонницы - суворексант, лемборексант). Эксперименты показали эффективность интраназального приёма орексина (полипептид Орексина А, не уступает внутривенному приёму по эффекту)[7].
Препараты в стадии разработки
- SB-649,868 — неселективный антагонист орексинных рецепторов (GlaxoSmithKline).
- ACT-078573, almorexant — неселективный антагонист орексинных рецепторов (разработан швейцарской фирмой Actelion, права куплены GlaxoSmithKline)[1] (англ.)
К тому же, есть несколько небелковых препаратов:
- SB-334,867 — селективный антагонист OX1
- SB-408,124 — селективный антагонист OX1
- TCS-OX2-29 — селективный антагонист OX2
См. также
Примечания
- ↑ 1 2 3 4 5 6 Kousaku Ohno and Takeshi Sakurai, Orexin neuronal circuitry: Role in the regulation of sleep and wakefulness, Frontiers in Neuroendocrinology, Т. 29, № 1, янв. 2008, С. 70—87.
- ↑ 1 2 3 T. Sakurai, Reverse pharmacology of orexin: from an orphan GPCR to integrative physiology, Regulatory peptides, 126 (2005), C. 3—10.
- ↑ John W. Muschamp, HYPOCRETIN (OREXIN), DOPAMINE, AND GOAL-DIRECTED BEHAVIOR, The Florida State University, College of Arts and Sciences, A dissertation submitted to the Department of Psychology in partial fulfillment of the requirements for the degree of Doctor of Philosophy, Spring Semester, 2007.
- ↑ Thomas Chacko Thannickal, A decade of hypocretin/orexin: Accomplishments in sleep medicine, Sleep Medicine Reviews, Т. 13, № 1, фев. 2009, С. 5—8.
- ↑ R. J. Rodgers et al, Orexins and appetite regulation, Neuropeptides Т. 36, № 5, окт. 2002, С. 303—325.
- ↑ M. V. Heinonen et al, Functions of orexins in peripheral tissues, Acta Physiologica Т. 192 № 4, С. 471—485.
- ↑ SV Dhuria, LR Hanson, WH Frey, Intranasal drug targeting of hypocretin-1 (orexin-A) to the central nervous system II — Journal of Pharmaceutical Sciences, 2008 — interscience.wiley.com
Источники
- http://vivovoco.astronet.ru/VV/JOURNAL/NATURE/11_05/SLEEP.HTM
- The neural circuit of orexin (hypocretin): maintaining sleep and wakefulness, Takeshi Sakurai, Nature Reviews Neuroscience 8, 171—181 (March 2007)
- Blockade of orexin-1 receptors attenuates orexin-2 receptor antagonism-induced sleep promotion in the rat, Christine Dugovic et al, JPET Fast Forward. Published on April 10, 2009 .
- The orexin/hypocretin system: a critical regulator of neuroendocrine and autonomic function, Alastair V. Fergusona, and Willis K. Samsonb, 2003 Elsevier Inc.
- Promotion of sleep by targeting the orexin system in rats, dogs and humans, Brisbare-Roch C. et al. Nat Med. 2007 Feb;13 (2):150-5.
- http://edrv.endojournals.org/cgi/content/full/27/7/779
- https://web.archive.org/web/20090523104859/http://www1.actelion.com/en/scientists/development-pipeline/phase-3/almorexant.page
- https://web.archive.org/web/20090621165352/http://www.redorbit.com/news/health/1052463/new_data_on_orexin_receptor_antagonist_almorexant_shows_therapeutic_potential/index.html