Паразавролоф
| † Паразавролоф | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
Домен: Эукариоты Царство: Животные Подцарство: Эуметазои Без ранга: Двусторонне-симметричные Без ранга: Вторичноротые Тип: Хордовые Подтип: Позвоночные Инфратип: Челюстноротые Надкласс: Четвероногие Клада: Амниоты Клада: Завропсиды Клада: Архозавры Клада: Авеметатарзалии Клада: Динозавроморфы Клада: Динозавры Отряд: † Птицетазовые Подотряд: † Цераподы Инфраотряд: † Орнитоподы Парвотряд: † Игуанодонты Надсемейство: † Гадрозавроиды Семейство: † Гадрозавриды Подсемейство: † Ламбеозаврины Триба: † Parasaurolophini Род: † Паразавролоф | |||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Parasaurolophus Parks[англ.], 1922 | |||||||||||||||||||||||||||||||||||||||||
| Синонимы | |||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
| Виды | |||||||||||||||||||||||||||||||||||||||||
| Геохронология 76,5—73 млн лет
◄ Наше время◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание◄ Массовоепермское вымирание ◄ Девонское вымирание◄ Ордовикско-силурийское вымирание◄ Кембрийский взрыв | |||||||||||||||||||||||||||||||||||||||||


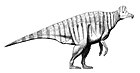
Паразавролоф[1][2], или паразауролоф[3] (лат. Parasaurolophus, от др.-греч. para — возле, и Saurolophus: др.-греч. sauros — ящер, и др.-греч. lophos — гребень[4]), — род птицетазовых динозавров из семейства гадрозаврид (Hadrosauridae), живших во времена позднемеловой эпохи, около 76,5—73 млн лет назад[5]. Ископаемые остатки обнаружены на территории Северной Америки. Окаменелости обнаружены в провинции Альберта в Канаде и Нью-Мексико и Юта в США. Впервые род описан в 1922 году Уильямом Парксом[англ.][6].
Описание

Как и в случае с большинством динозавров, скелеты паразавролофов являются неполными. Длина голотипа P. walkeri оценочно составляет 9,45 м[7], оценки массы тела на основе аллометрии показывают, что 9-метровая особь весила более 5 т[8]. Грегори Пол[англ.] оценил среднюю длину взрослого P. walkeri в 7,5 м, а массу в 2,6 т[3]. Череп P. walkeri достигает примерно 1,6 м в длину, включая гребень, в то время как череп P. tubicen превышает 2 м, что указывает на более крупные размеры животного[9]. Единственная известная передняя конечность относительно короткая для гадрозаврид, с короткой, но широкой лопаткой. Бедренная кость P. walkeri достигает в длину 103 см и массивная для своей длины, если сравнивать с костями других гадрозаврид[9]. Плечевая кость и кости таза тоже сложены очень крепко[10].
Как и другие гадрозавриды, паразавролофы могли ходить как на двух, так и на четырёх конечностях. Вероятно они предпочитали пастись на четырёх конечностях, а бегать на двух[11]. Нервные отростки на позвонках высокие, что характерно для ламбеозаврин (Lambeosaurinae)[9]. В самом высоком месте над бёдрами спина благодаря ним увеличивала высоту. Известны отпечатки кожи P. walkeri с однородными бугоркообразные чешуйками, но без более крупных структур[6].
Череп

Наиболее заметной особенностью животного является гребень, выступающий из задней части головы и состоящий из предчелюстной и носовой костей[9]. Гребень был полым, с отдельными трубками, ведущими от каждой ноздри к концу гребня, где они меняют направление и ведут обратно вниз по гребню в череп. Трубки самые простые у P. walkeri и более замысловатые у P. tubicen, у которого некоторые трубки были сплошные, а другие сходятся и расходятся[12]. В то время как P. walkeri и P. tubicen обладали длинными гребнями с небольшой кривизной, у P. cyrtocristatus имелся короткий гребень с более округлым профилем[13].
Палеоэкология
Альберта
Parasaurolophus walkeri из формации Парк динозавров[англ.] является частью разнообразной и хорошо задокументированной доисторической фауны, включающей хорошо известных динозавров, таких как рогатые Centrosaurus, хасмозавр (Chasmosaurus) и стиракозавр (Styracosaurus), орнитомимид струтиомим (Struthiomimus), утконосые грипозавр (Gryposaurus) и коритозавр (Corythosaurus), тираннозавриды горгозавр (Gorgosaurus) и дасплетозавр (Daspletosaurus), и бронированные эдмонтония (Edmontonia), Euoplocephalus и Dyoplosaurus[11]. P. walkeri — редкий элемент этой фауны[14]. Ландшафт формации Парк динозавров реконструируется как низкие речные поймы, которые со временем стали более заболоченными и подвергались влиянию моря, поскольку Западное внутреннее море трансгрессировало в западном направлении[14]. Климат был теплее, чем в современной Альберте, без заморозков, но с влажными и сухими сезонами. Хвойные деревья, видимо, преобладали в пологе лесов, тогда как в подлеске произрастали папоротники, древовидные папоротники и цветковые растения[14].
Некоторые из менее распространённых гадрозаврид формации Парк динозавров в Провинциальном парке динозавров, включая Parasaurolophus, могут быть представлены остатками особей, которые умерли во время миграции через данный регион. Не исключено, что их средой обитания являлись более высокие местности, где животные гнездились или кормились. Присутствие Parasaurolophus и Kritosaurus в ископаемых отложениях северных широт может иллюстрировать фаунистический обмен между различными северными и южными биомами в позднемеловой Северной Америке. Оба таксона редки за пределами южного биома, где, наряду с пентацератопсами (Pentaceratops), они являются преобладающими представителями фауны[15].
Нью-Мексико

В формации Фрутленд[англ.] в Нью-Мексико P. cyrtocristatus делил жизненное пространство с другими орнитисхиями и тероподами. В частности, его современниками были цератопс Pentaceratops sternbergii[11], пахицефалозавр Stegoceras novomexicanum[16]. В этих же отложениях найдены неопознанные окаменелости, принадлежащие тираннозавридам, ?Ornithomimus, ?Troodontidae, ?Saurornitholestes langstoni, ?Struthiomimus, Ornithopoda, ?Chasmosaurus, ?Corythosaurus, Hadrosaurinae, гадрозавридам и цератопсидам[11]. Во времена паразавролофов формация Фрутленд была болотистой местностью, расположенной в низинах и близко к берегу Западного внутреннего моря. Самая нижняя часть формации Фрутленд чуть моложе 75,56 ± 0,41 млн лет назад, а самая верхняя граница датируется 74,55 ± 0,22 млн лет назад[17].
Появившийся немного позже, чем вид из формации Фрутленд, P. tubicen также найден в Нью-Мексико, в формации Киртленд[англ.][11]. Из этой формации происходят многочисленные группы позвоночных, в том числе рыбы, крокодиломорфы[17], орнитисхии, заврисхии[11], птерозавры[18] и черепахи. Рыбы представлены двумя видами: Melvius chauliodous и Myledalphus bipartitus. К крокодиломорфам относятся Brachychampsa montana и Denazinosuchus kirtlandicus[17]. Орнитисхии из формации представлены гадрозавридами Anasazisaurus horneri, Naashoibitosaurus ostromi, Kritosaurus navajovius и P. tubicen, анкилозавридами Ahshislepelta minor и Nodocephalosaurus kirtlandensis, цератопсами Pentaceratops sternbergii[11] и Titanoceratops ouranos[19], пахицефалозаврами Stegoceras novomexicanum[16] и Sphaerotholus goodwini[17]. К заврисхиям относятся тираннозаврид Bistahieversor sealeyi[20], орнитомимид Ornithomimus sp.[11] и троодонтид «Saurornitholestes» robustus[21]. Известен один птерозавр — Navajodactylus boerei[18]. Черепахи довольно многочисленны: в формации найдены Denazinemys nodosa, Basilemys nobilis, Neurankylus baueri, Plastomenus robustus и Thescelus hemispherica. Неидентифицированные таксоны включают окаменелости крокодиломорфа ?Leidyosuchus[17] и теропод ?Struthiomimus, троодонтид и тираннозаврид[11]. Нижняя граница формации Киртленд датируется 74,55 ± 0,22 млн лет назад, а верхняя — около 73,05 ± 0,25 млн лет назад[17].
Юта

Аргон-аргоновое радиометрическое датирование[англ.] показывает, что формация Кайпаровиц[англ.] образовалась между 76,6 и 74,5 миллионами лет назад, в кампанский век позднего мелового периода[22][23]. В то время участок формации Кайпаровиц располагался недалеко от западного берега Западного внутреннего моря, разделявшего Северную Америку на два сухопутных массива: Ларамидию на западе и Аппалачию на востоке. Плато, где жили динозавры, представляло собой окружённую горами древнюю пойму, в которой преобладали большие каналы и обширные торфяные болота, пруды и озёра. Климат был влажным и благоприятствовал самым разным организмам[24]. В этой формации представлена одна из лучших и наиболее протяжённых во времени позднемеловых наземных биот в мире[25].
Паразавролоф делил окружающую среду с другими динозаврами, такими как дромеозавридные тероподы, троодонтид Talos sampsoni, орнитомимиды, такие как Ornithomimus velox, тираннозавриды, такие как тератофоней (Teratophoneus), панцирные анкилозавриды, гадрозаврид Gryposaurus monumentensis, цератопсы Utahceratops gettyi, Nasutoceratops titusi и Kosmoceratops richardsoni и овирапторозавр Hagryphus giganteus[26]. Палеофауна, присутствующая в формации Кайпаровиц, включала хрящевых рыб (акулы и скаты), лягушек, саламандр, черепах, ящериц и крокодилов, таких как высший хищник Deinosuchus. Здесь же обитали многочисленные ранние млекопитающие, включая многобугорчатых, сумчатых и насекомоядных[27].
Ссылки
- Parasaurolophus sound. Sandia National Laboratories (5 декабря 1997). Архивировано из оригинала 6 октября 2014 года.
Примечания
- ↑ Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М. : ГЕОС, 2009. — С. 144. — 377 с. : ил. — (Труды ПИН РАН ; т. 291). — 600 экз. — ISBN 978-5-89118-461-9.
- ↑ А. В. Лопатин. Палеонтологический музей имени Ю.А. Орлова. — Москва: ПИН РАН, 2012. — P. рис. 107, V-12. — ISBN 978-5-903825-14-1. Архивная копия от 8 октября 2020 на Wayback Machine
- ↑ 1 2 Пол Г. С.[англ.]. Динозавры. Полный определитель = The Princeton Field Guide to Dinosaurs : ориг. изд. 2016 / пер. с англ. К. Н. Рыбакова ; науч. ред. А. О. Аверьянов. — М. : Фитон XXI, 2022. — С. 341—342. — 360 с. : ил. — ISBN 978-5-6047197-0-1.
- ↑ Colbert, Edwin H. (Edwin Harris). The dinosaur book: the ruling reptiles and their relatives / Edwin H. (Edwin Harris) Colbert, Charles Robert Knight. — New York : McGraw-Hill, 1951. — P. 152.
- ↑ Evans, D.C.; Bavington, R.; Campione, N.E. (2009). "An unusual hadrosaurid braincase from the Dinosaur Park Formation and the biostratigraphy of Parasaurolophus (Ornithischia: Lambeosaurinae) from southern Alberta". Canadian Journal of Earth Sciences. 46 (11): 791—800. doi:10.1139/E09-050.
- ↑ 1 2 Parks, W.A. (1922). "Parasaurolophus walkeri, a new genus and species of trachodont dinosaur". University of Toronto Studies: Geological Series. 13: 5—32.
- ↑ Farke, A.A.; Chok, D.J.; Herrero, A.; Scolieri, B.; Werning, S. (2013). Hutchinson, John (ed.). "Ontogeny in the tube-crested dinosaur Parasaurolophus (Hadrosauridae) and heterochrony in hadrosaurids". PeerJ. 1: e182. doi:10.7717/peerj.182. PMC 3807589. PMID 24167777.
- ↑ Seebacher, F. (2001). "A new method to calculate allometric length-mass relationships of dinosaurs" (PDF). Journal of Vertebrate Paleontology. 21 (1): 51—60. doi:10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2. JSTOR 4524171. S2CID 53446536. Архивировано (PDF) 4 октября 2013. Дата обращения: 16 февраля 2024.
- ↑ 1 2 3 4 Lull, R.S. Hadrosaurian Dinosaurs of North America / R.S. Lull, N.E. Wright. — Geological Society of America, 1942. — P. 229.
- ↑ Brett-Surman, Michael K. Appendicular anatomy in Campanian and Maastrichtian North American hadrosaurids // Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs / Michael K. Brett-Surman, Wagner, Jonathan R.. — Bloomington and Indianapolis : Indiana University Press, 2006. — P. 135–169. — ISBN 978-0-253-34817-3.
- ↑ 1 2 3 4 5 6 7 8 9 Weishampel, David B. Dinosaur Distribution // The Dinosauria / Weishampel, David B., Barrett, Paul M., Coria, Rodolfo A. … [и др.]. — 2nd. — 2004. — P. 517–606.
- ↑ Sullivan, R.S.; Williamson, T.E. (1999). "A new skull of Parasaurolophus (Dinosauria: Hadrosauridae) from the Kirtland Formation of New Mexico and a revision of the genus" (PDF). New Mexico Museum of Natural History and Science Bulletin. 15: 1—52. Архивировано из оригинала (PDF) 8 апреля 2023.
- ↑ Ostrom, J.H. (1961). "A New Species of Hadrosaurian Dinosaur from the Cretaceous of New Mexico". Journal of Paleontology. 35 (3): 575—577. JSTOR 1301139.
- ↑ 1 2 3 Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. — Bloomington : Indiana University Press, 2005. — P. 312–348. — ISBN 978-0-253-34595-0.
- ↑ Mesozoic Vertebrate Life. — Indiana University Press, 2001. — P. 206–328. — ISBN 978-0-253-33907-2.
- ↑ 1 2 Jasinski, S.E.; Sullivan, R.M. (2011). "Re-evaluation of pachycephalosaurids from the Fruitland-Kirtland transition (Kirtlandian, late Campanian), San Juan Basin, New Mexico, with a description of a new species of Stegoceras and a reassessment of Texascephale langstoni". New Mexico Museum of Natural History and Science, Bulletin. Fossil Record 3. 53: 202—215.
- ↑ 1 2 3 4 5 6 Sullivan, R.M. The Kirtlandian Land-Vertebrate "Age"-Faunal Composition, Temporal Position, and Biostratigraphic Correlation in the Nonmarine Upper Cretaceous of Western North America // Late Cretaceous vertebrates from the Western Interior / R.M. Sullivan, S.G. Lucas. — 2006. — Vol. 35. — P. 7–23.
- ↑ 1 2 Sullivan, R.M.; Fowler, D.W. (2011). "Navajodactylus boerei, n. gen., n. sp., (Pterosauria, ?Azhdarchidae) from the Upper Cretaceous Kirtland Formation (upper Campanian) of New Mexico" (PDF). Fossil Record 3. New Mexico Museum of Natural History and Science, Bulletin. 53: 393—404. Архивировано (PDF) 4 февраля 2012. Дата обращения: 16 февраля 2024.
- ↑ Longrich, N.R. (2011). "Titanoceratops ouranous, a giant horned dinosaur from the Late Campanian of New Mexico" (PDF). Cretaceous Research. 32 (3): 264—276. doi:10.1016/j.cretres.2010.12.007. Архивировано (PDF) 22 февраля 2014. Дата обращения: 16 февраля 2024.
- ↑ Carr, T.D.; Williamson, T.E. (2010). "Bistahieversor sealeyi, gen. et sp. nov., a new tyrannosauroid from New Mexico and the origin of deep snouts in Tyrannosauroidea". Journal of Vertebrate Paleontology. 30 (1): 1—16. doi:10.1080/02724630903413032. S2CID 54029279.
- ↑ Evans, D.C.; Larson, D.W.; Cullen, T.M.; Sullivan, R.M. (2014). Sues, Hans-Dieter (ed.). ""Saurornitholestes" robustus is a troodontid (Dinosauria: Theropoda)". Canadian Journal of Earth Sciences. 51 (7): 730—734. Bibcode:2014CaJES..51..730E. doi:10.1139/cjes-2014-0073.
- ↑ Roberts, E.M.; Deino, A.L.; Chan, M.A. (2005). "40Ar/39Ar age of the Kaiparowits Formation, southern Utah, and correlation of contemporaneous Campanian strata and vertebrate faunas along the margin of the Western Interior Basin". Cretaceous Research. 26 (2): 307—318. doi:10.1016/j.cretres.2005.01.002.
- ↑ Eaton, J.G. (2002). "Multituberculate mammals from the Wahweap (Campanian, Aquilan) and Kaiparowits (Campanian, Judithian) formations, within and near Grand Staircase-Escalante National Monument, southern Utah". Miscellaneous Publication 02-4, UtahGeological Survey: 1—66.
- ↑ At the Top of the Grand Staircase: The Late Cretaceous of Southern Utah. — Indiana University Press, 2013. — P. 1–634.
- ↑ Clinton, William Presidential Proclamation: Establishment of the Grand Staircase-Escalante National Monument. September 18, 1996. Дата обращения: 16 февраля 2024. Архивировано из оригинала 28 августа 2013 года.
- ↑ Zanno, L.E.; Sampson, S.D. (2005). "A new oviraptorosaur (Theropoda; Maniraptora) from the Late Cretaceous (Campanian) of Utah". Journal of Vertebrate Paleontology. 25 (4): 897—904. doi:10.1671/0272-4634(2005)025[0897:ANOTMF]2.0.CO;2. S2CID 131302174.
- ↑ Eaton, J.G. Cretaceous vertebrate faunas from the Kaiparowits Plateau, south-central Utah // Vertebrate Paleontology in Utah / J.G. Eaton, R.L. Cifelli, J.H. Hutchinson … [и др.]. — Salt Lake City : Utah Geological Survey, 1999. — P. 345–353. — ISBN 978-1-55791-634-1.
| Базальные | 
 | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hadrosauro- morpha |
| ||||||||||||||||||||||||||||||