Запросы «Паразит» и «Паразиты» перенаправляются сюда; см. также другие значения.
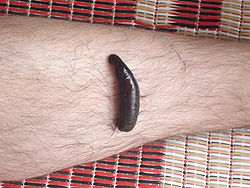
Гнездовой паразитизмСамка комара Anopheles stephensiПостельный клоп. От переднего края головы отходит хоботок, приспособленный для прокола тканей и сосания кровиПиявка на ноге человека. Гематофагия — одна из самых распространённых форм паразитизмаОбыкновенный вампир. Среди млекопитающих гемофагия встречается у вампировых летучих мышей. Это единственные облигатные паразиты среди теплокровных позвоночныхПчёлы-кукушки из рода Nomada. Пчёлы этой экологической группы проникают в гнёзда других видов и родов пчёл, где откладывают свои яйца. Вышедшая из яйца личинка паразита питается запасом провианта вида-хозяина.Заразиха — родрастений-паразитов из семействаЗаразиховые (Orobanchaceae), сорняк. Виды заразихи живут на корнях весьма разнообразных растений, преимущественно из семейства Бобовые, иногда причиняют своим хозяевам большой вред
Паразити́зм (от др.-греч.παράσιτος — «нахлебник, сотрапезник») — один из типов сосуществования организмов. Явление, при котором два и более организма, не связанных между собой филогенетически, генетически разнородных — сосуществуют в течение продолжительного времени и при этом находятся в антагонистических отношениях (или же в односторонних симбиотических[1]). Это вид взаимосвязей между различными видами, при котором один из них — паразит — определённое время использует другого (который называется хозяином) в качестве источника питания и среды обитания, частично или полностью возлагая на него регуляцию своих взаимоотношений с окружающей средой.[2]
Паразитизм встречается среди различных групп организмов: животных (простейшие, плоские черви, нематоды, кольчатые черви, моллюски, членистоногие), бактерий, грибов (мучнисторосяные, трутовики) и даже у покрытосеменных растений[3]. Физиология паразита подчинена физиологии хозяина, и его жизненный цикл (само его существование или размножение) невозможен (или сильно затруднён) без получения от хозяина необходимых для него биологических ресурсов. Такие ресурсы паразит может получить только от ограниченного числа типов хозяев. Чем дольше (филогенетически) продолжается сосуществование, тем лучше этот вид паразитов приспосабливается к своему хозяину и тем меньше вреда наносит ему. В сфере медицинской паразитологии термин «паразит» означает эукариотический патогенный организм. Простейшие и многоклеточные возбудители инфекции классифицируются как паразиты. Грибы не обсуждаются в учебниках медицинской паразитологии, хотя являются эукариотами. Ни один из известных представителей архей (они не являются эукариотами) не является паразитом, за исключением наноархеот, являющихся паразитами других архей. Среди внутриклеточных паразитов наименьшие размеры имеют эубактерии рода Mycoplasma, например Mycoplasma genitalium, диаметр клеток которой составляет 200—300 нм.
Формы паразитизма и связанные с этим взаимные адаптации паразитов и их хозяев чрезвычайно многообразны. Паразиты бывают как животные (зоопаразиты, то есть паразитирующие на животных организмах), так и растительные (фитопаразиты). Обычно это многие грибы, а также бактерии и миксомицеты. Иногда встречаются и паразитические семенные растения. Животных фитопаразитов обычно называют вредителями. Ими наиболее часто являются насекомые, клещи[4], нематоды и некоторые простейшие.
Под паразитологией традиционно понимают только зоопаразитологию. Эта наука делится на ветеринарную, медицинскую, агрономическую, лесную и общую. В более широком смысле паразитология подразделяется на вирусологию, микробиологию, мико[5]-, фито- и зоопаразитологию. Задачу объединения всех разделов паразитологии ставит перед собой комплексная наука паразитоценология.
Различают эктопаразитизм, при котором паразит обитает на хозяине и связан с его покровами (клещи, блохи, вши и др.), и эндопаразитизм, при котором паразит живёт в теле хозяина (паразитические черви, простейшие и др.)[6]. Эктопаразитизм в природе встречается гораздо реже эндопаразитизма. Эта форма паразитизма более характерна для растений паразитов, многих насекомых и клещей, а также грибов. Эктопаразиты, из группы паразитических перепончатокрылых, как правило, заражают скрытоживущих хозяев (развивающихся внутри плодов, галлов, древесины) и поэтому прикрепляют свои яйца снаружи, парализуя добычу. Разделение паразитов на экто- и эндопаразитов связано с глубиной их проникновения в кожные и подкожные ткани организма. Обычно эндопаразиты в поисках хозяина полагаются на пассивные механизмы (распространение яиц или личинок для случайного поедания животным-хозяином и т. д.), а эктопаразиты распространяются активно: так, у беременных самок кроликов перед родами заметно увеличивается количество блох, которые затем переходят на потомство.
Некротрофия
Паразиты-некротрофы используют хозяина, вызывая его смерть от недостатка питательных веществ или от побочных эффектов жизнедеятельности паразита (например, таким паразитом для человека является бактерия — возбудитель чумы). Паразиты-некротрофы в случае, когда они могут выживать после смерти хозяина, называются паразитоидами. Паразиты-биотрофы обычно не приводят к смерти хозяина и не могут выжить в случае его смерти. Такими паразитами, например, являются многочисленные вирусы.
Сверхпаразитизм
Сверхпаразиты (паразиты паразитов, надпаразиты или гиперпаразиты) заражают других паразитов. Паразит такого типа паразитирует на первичном паразите (то есть виде, паразитирующем на особи, не являющейся паразитом) и соответственно называется вторичным паразитом. Очень редко, но всё же встречаются и сверхпаразиты 3-го (третичные) и даже 4-го порядка. Например, наездник Asecodes albitarsus заражает многоядного сверхпаразита Dibrachys boucheanus, а он, в свою очередь, может поражать наездников Apanteles glomeratus, паразитирующих на гусеницах бабочек-белянок. Сверхпаразитизм широко распространён среди паразитических насекомых, в частности у наездников семейств Ichneumonidae, Encyrtidae и некоторых других[7].
Сверхпаразитизм отличается от множественного паразитизма (т. н. мультипаразитизма), где хозяин одновременно заражён двумя или несколькими первичными паразитами разных видов. Чрезмерное количество паразитов в одном хозяине (перезаражение) негативно сказывается на их развитии, но некоторые паразиты, по всей видимости, не способны отличить заражённого хозяина от незаражённого. Такое явление называется суперпаразитизмом (то есть отличается от вышеупомянутого сверхпаразитизма).
Прочие виды паразитизма
По степени тесноты связей паразита и хозяина выделяют две формы паразитизма: облигатный и факультативный. В первом случае вид ведёт только паразитический образ жизни и не выживает без связи с хозяином (паразитические черви, вши). Факультативные паразиты, как правило, ведут свободный образ жизни и лишь при особых условиях переходят к паразитическому состоянию. По продолжительности связей с хозяином существуют постоянные, то есть проводящие на хозяине большую часть жизни, и временные паразиты (нападают только для питания). Постоянные паразиты в свою очередь подразделяются на стационарных и периодических. Стационарные паразиты всю жизнь проводят на хозяине или внутри него[8]. Периодические паразиты в паразитическом состоянии проводят лишь часть своего жизненного цикла, а в остальное время обитают вне хозяина[9]. Например, комар является непостоянным паразитом, то есть взаимодействие между организмами носит эпизодический характер, когда самки комаров потребляют кровь хозяина. Следует отличать временный паразитизм от случайного паразитизма (псевдопаразитизма или ложного паразитизма), при котором имеется случайное проникновение какого-либо свободноживущего организма в тело другого организма, где он обитает и питается непродолжительное время. Например, личинки мух[10] могут случайно проникнуть и временно обитать в кишечнике человека и животных.
На сегодняшний день известен только один паразит, который функционально замещает собой орган хозяина — Cymothoa exigua. Существуют также различные формы «социального паразитизма»: клептопаразитизм (то есть присвоение чужой пищи), в том числе его особая форма — т. н. яичный паразитизм, наблюдаемый у некоторых видов рыб, птиц и насекомых, когда для высиживания яиц и воспитания новорождённых один организм подкидывает свои яйца в гнездо другого, которые затем выкармливают чужих птенцов (характерный пример — кукушка) и др. Социальные паразиты получают выгоду от взаимодействия с группировками социальных животных (например муравьёв или термитов), используя особенности их общественной организации (таковы, например, несколько видов жуков, живущих в муравейниках, которые питаются на «складах» продуктов внутри них и получают защиту от врагов). Клептопаразитами являются, например, тропические птицы фрегаты, которые сами почти не могут добывать рыбу, но отбирают её у других птиц. В 1909 году итальянский энтомологКарло Эмери (Carlo Emery) заметил, что социальные паразиты среди насекомых (например клептопаразиты), в основном, паразитируют на особях близкого им вида или рода[11][12]. С годами это было замечено и в других случаях. Данному феномену есть объяснение: паразиты могли изначально быть факультативными паразитами[англ.] среди представителей самого вида-хозяина (известно много примеров подобного внутривидового паразитизма), однако затем отделились от родительского вида и образовали свой собственный обособленный вид (один из примеров симпатрического видообразования).
Существует особая форма клептопаразитизма, при котором взрослая особь эктопаразита использует уже заражённого и парализованного хозяина, при этом клептопаразит устраняет личинок первичного паразита для устранения перезаражения и конкуренции для своих личинок. У клещей представителей надсемейства Argasidae встречается явление омовампиризма, когда голодная особь нападает на сытого «собрата» и питается выпитой им кровью.
Биологическое мошенничество как тип паразитизма обычно развивается в ситуациях генерализованного неспецифического мутуализма между широкими спектрами организмов — таких, например, как образование микоризы между грибами и растениями. Некоторые виды растений выработали приспособления к образованию «микоризоподобной» структуры с гифами грибницы, при этом не снабжая грибницу питательными веществами в обмен на минеральные (как это бывает в нормальной, симбиотической грибнице), а высасывая их отсюда — при этом гриб, образовавший микоризу, получает их из другого растения. Паразитическое растение, участвующее в отношениях, при которых растения получают часть или все необходимые питательные вещества не через фотосинтез, а паразитируя на грибе, называется микогетеротрофом. Полная, или облигатная, микогетеротрофия имеет место тогда, когда нефотосинтезирующее растение (растение, лишённое или хлорофилла, или функциональной фотосистемы) получает все необходимые питательные вещества от гриба, на котором паразитирует. О частичной, или факультативной, микогетеротрофии говорят тогда, когда растение способно к фотосинтезу, однако паразитирует на грибе как на дополнительном источнике питательных веществ. Существуют растения (например некоторые виды орхидных), которые не фотосинтезируют и являются облигатными микогетеротрофами на некотором этапе своего жизненного цикла, а в остальное время способны фотосинтезировать и являются факультативными микогетеротрофами или не используют микогетеротрофию вообще[13]. Не все нефотосинтезирующие, или бесхлорофилльные, растения являются микогетеротрофами: например, повилика паразитирует непосредственно на проводящих тканях других растений[14].
Эпифития
Эпифиты не являются паразитарными растениями. Они растут независимо от растения-опоры и используют её исключительно в качестве физической опоры. Эпифиты используют фотосинтез для получения энергии и питательных веществ и, в случае неводных видов, получают влагу из воздуха и осадков (дождь, туман и др.). Очевидным преимуществом эпифитов, особенно в сильно заросшей деревьями местности, является возможность не зависеть от грунта, а находиться ближе к источнику света; на эпифитах также меньше сказывается влияние травоядных. Тем не менее при слишком густом нарастании эпифитов они могут повредить растение-опору.
Паразитизм между растениями и другими царствами
В прошлом ошибочно полагали, что некоторые растения могут получать питательные вещества, разлагая органические вещества, подобно сапротрофным грибам. Такие растения называли сапрофитами. Сейчас известно, что ни одно растение не способно непосредственно разрушать органические соединения и нефотосинтезирующие растения получают необходимые питательные вещества через паразитизм: микогетеротрофию или непосредственный паразитизм на других растениях[15][16].
При микогетеротрофии у гриба и растения контактируют мицелий и корни соответственно. В этом отношении микогетеротрофия очень похожа на микоризу (в самом деле, считается, что микогетеротрофия произошла от микоризы), однако при микогетеротрофии углеродные соединения переносятся от гриба к растению, а не наоборот, как при микоризе[17][18]. Хотя термин «симбиоз» в англоязычной литературе используется в широком смысле — для обозначения мутуализма, комменсализма и паразитизма — эндосимбионтами обычно называют лишь организмы, которые находятся с хозяевами во взаимовыгодных (мутуалистических) взаимоотношениях. Отличие паразитизма от мутуализма состоит в том, что хотя каждый из участников мутуалистической пары действует эгоистично, выгодные отношения возникают лишь потому, что получаемая польза перевешивает затраты, требуемые на поддержание взаимоотношений. Большую часть микогетеротрофов можно рассматривать как эпипаразитов, поскольку они получают энергию от грибов, которые, в свою очередь, получают её от сосудистых растений[15][16][19]. Действительно, чаще всего микогетеротрофия существует в контексте микоризной сети[англ.][20], в которой растения используют микоризные грибы для обмена углеродом и питательными веществами с другими растениями[16]. В этих системах микогетеротрофы играют роль «микоризных обманщиков», поскольку они забирают углерод из общей системы и ничего не отдают взамен[15].
Ассоциация эндофита и растения часто описывается как пример симбиоза. Микроорганизм-эндофит выделяет вещества, способствующие росту растения (например, ацетобактерAcetobacter diazotrophicus, живущий в тканях сахарного тростника, по неустановленному механизму способен к фиксации атмосферного азота), или антибиотические вещества, подавляющие развитие патогенов или предотвращающие заражение здорового растения патогенами, Однако характеризация отношений эндофитов и их хозяев как мутуализм может быть достаточно условна. Различают облигатных и факультативных эндофитов. К облигатным относятся трудновыделяемые микроорганизмы, способные существовать только внутри растений. Факультативными эндофитами являются организмы, способные существовать в растении, не причиняя ему вреда, однако при иных обстоятельствах, например при угнетённости растения, становящиеся патогенами или сапротрофами.
Паразитизм и другие типы межвидовых отношений
Классификационное разграничение между паразитизмом и хищничеством может быть спорно, и иногда в широком смысле под термином хищничество понимают всякое выедание одних организмов другими (полное или частичное без умерщвления), то есть отношение паразитов и их хозяев.
паразитов, продолжительное время питающихся одной жертвой и не вызывающих её гибели.
Также существуют животные, совмещающие хищничество с паразитизмом. Клопы из семейства Хищнецы — активные хищники, питаются преимущественно насекомыми, но ряд тропических видов питается и кровью теплокровных животных и человека и таким образом представляет переход от хищников к паразитам.
Аналогично этому разграничение между комменсализмом и паразитизмом также неочевидно, например когда одно животное (инквилин), проникая в чужое жилище, уничтожает его хозяина, после чего использует жилище в своих целях (инквилинизм). Изначально инквилинизм выделялся как подвид комменсализма, хотя он очень близок к хищничеству и паразитизму.
Эволюция
Биотрофичный паразитизм обычно является весьма успешным приспособлением. В зависимости от системы определений, около половины известных видов животных имеет как минимум одну паразитическую стадию в своём жизненном цикле[21]; также паразитизм довольно часто встречается среди растений и грибов. С другой стороны, практически все свободноживущие виды животных являются хозяевами одного или нескольких таксонов паразитов.
Организмы-хозяева обычно в ходе эволюции также изменяются; у них появляются защитные механизмы против паразитов. Растения часто продуцируют токсины, которые могут одновременно оказывать эффективное воздействие на паразитических грибов, бактерий и растительноядных животных. Иммунная система позвоночных способна нейтрализовывать большинство паразитов (особенно микропаразитов[англ.]) при их контакте с жидкостями организма.
В свою очередь, большинство паразитов, особенно микроорганизмов, также приобретает защитную адаптацию для противодействия защитным механизмам хозяев. Такие адаптации могут принимать различные формы: утолщение клеточных стенок бактерий, клещи-захваты на конечностях, предотвращающие вычёсывание хозяевами блох и клещей из волосяного покрова, и т. д.
При описанном типе взаимодействия оба вида (хозяин и паразит) обычно совместно эволюционируют к более-менее стабильному состоянию, когда их влияние на численность друг друга становится минимально возможным. Однако это правило не относится к паразитоидам, имеющим хищнические задатки. Например, в период размножения летом мухи лягушкоедки откладывают свои яйца на спину или голову жаб и других амфибий. Вылупившиеся личинки проникают через носовые отверстия в голову хозяина, где они въедаются в мягкие ткани ещё живущего животного. Когда личинки добираются до мозга или других жизненно важных органов, функции организма нарушаются и животное погибает. В своей последней фазе личинки почти полностью съедают труп животного, включая кожу и соединительные ткани, а при определённых обстоятельствах даже хрящи и кости[22]. Затем они закапываются во влажную почву и окукливаются.
Среди высших растений прямые (контактные) взаимовлияния возникают при соприкосновении или проникновении организмов друг в друга. Подразделяются на физиологические (паразитизм и симбиоз), когда между организмами осуществляется активный обмен веществом и энергией, и механические (взаимоотношения эпифитов с форофитами — лиан с опорными растениями) — когда таковой отсутствует. Степень поражения растения паразитами зависит от особенностей растения-хозяина (разные виды поражаются неодинаково), от условий местообитания (в условиях засоления растения практически не поражаются), от наличия эволюционной сопряжённости паразита и хозяина (если её нет, у хозяина обычно отсутствуют механизмы защиты от паразита).
Против паразитических грибов и бактерий у растений в ходе эволюции сформировался комплекс защитных механизмов:
выделение растением фунгицидных и бактерицидных веществ, предотвращающих заражение паразитами или подавляющих их развитие;
наличие мощных покровных тканей, препятствующих проникновению паразитов;
особенности биохимического состава и метаболизма в клетках растения, препятствующие росту паразитов.
Несмотря на наличие подобных механизмов защиты, фитопаразиты способны вызывать многочисленные болезни растений, ведущие к их ослаблению и гибели. Растения-паразиты распространены не столь широко, но также могут значительно угнетать ценопопуляции и отдельные растения.
Иногда исследования паразитов помогают выявить родственные связи между различными видами их хозяев. Например, длительный диспут орнитологов относительно систематического положения фламинго — являются ли они более родственными с рядом Ciconiiformes (Аистообразные) или Anseriformes (Гусеобразные) — был решён благодаря исследованию их паразитов, которые оказались общими с таковыми у гусеобразных (впрочем, исследования ДНК в последние годы показали, что фламинго — не очень близкие родственники и этого отряда).
Вред, причиняемый паразитом хозяину
Такие определения, как «польза» и «вред», в случае паразитизма иногда относятся не к отдельным индивидам, а к виду в целом. Например, если при поражении паразитом индивид становится более сильным и выносливым, но теряет функцию размножения (как это бывает при поражении улитки некоторыми плоскими червями), надо считать, что в эволюционном смысле этот организм понёс ущерб от данного паразита.
Считается, что эволюция паразитов направлена на уменьшение летальности своих хозяев от заражения, настоящим паразитам выгодно долго эксплуатировать хозяев, а не убивать их. На начальном этапе коэволюции паразитов и хозяев происходит «гонка вооружений». Форма «паразитирования на убой» свойственна некоторым паразитоидам (например наездникам). Такая форма паразитизма может быть объяснена тем, что среди насекомых распространено запасание корма для личинок[23]. Многие перепончатокрылые и некоторые двукрылые откладывают яйца в тела живых животных (преимущественно насекомых), тем самым обеспечивая их легкодоступным кормом. Своеобразие этой формы запасания корма, являющейся комбинацией паразитизма с хищничеством, позволяет выделить паразитоидов в отдельную экологическую категорию.
Вред, причинённый паразитом хозяину, может варьироваться в очень широких пределах. Это могут быть механические повреждения, вызванные закупоркой различных каналов, что случается, если скопления паразитов могут закупорить кишечник или же протоки различных желез. Паразиты или их яйца могут тромбировать лимфатические сосуды, при этом вызывая вздутия в разных частях тела. Аналогично этому некоторые паразиты или продукты их жизнедеятельности могут вызывать эмболию кровеносных сосудов, приводящую к сердечной недостаточности, а закупорка капилляров приводит к их разрыву и возникновению кровоизлияний, эмболизация сосудов в жизненно важных участках организма в свою очередь приводит к смерти заражённого. Зачастую паразиты, распространяясь по организму, проникают в ткани и органы или прогрызают их: в результате в такие отверстия (например проделанные скребнями (Acanthocephala) в стенке кишечника) проникают бактерии, и развиваются болезни, такие как, например, перитонит. Паразиты также способны питаться тканями тела: так, в частности, возбудитель амёбной дизентерии Entamoeba histolytica переваривает клетки слизистой, образуя открытые раны, называемые амёбными язвами.
Паразиты приводят к нарушениям регуляторных механизмов или же даже кастрации или растворению костей и прибегают к гораздо более изощрённым методам использования хозяина в своих целях, таких как изменение поведения хозяина. Например, паразитический гриб Ophiocordyceps unilateralis развивается в муравьях-древоточцах Camponotus castaneus и Camponotus americanus, живущих в Южной Каролине (США). У заражённых муравьёв меняется поведение, и в итоге они умирают, накрепко вцепившись челюстями в ветви, что способствует распространению спор паразита. Американские биологи получили чистую культуру гриба-манипулятора и заразили им три вида муравьёв-древоточцев. Гриб успешно развивался в теле всех трёх видов, но выгодная грибу манипуляция предсмертногo поведения наблюдалась только в двух, которые являются жертвами паразита в природе. Выяснилось также, что клетки гриба выделяют различные наборы веществ в присутствии живой мозговой ткани муравьёв разных видов. Это свидетельствует о том, что влияние паразита на нервную систему хозяина видоспецифичнo, видимо, он подстраивается под биохимические особенности мозга жертвы.
Изумрудная тараканья оса нападает на тараканов и частично парализует их[24]. Для этого она жалит таракана, после чего он сохраняет способность к передвижению, но не может двигаться самостоятельно. Тогда оса берёт таракана за усики и ведёт к своей норе. Там она откладывает на его брюшке яйца и оставляет в норке. Появившиеся личинки используют таракана в качестве пищи, а затем окукливаются. Через некоторое время из норки выходит новое поколение ос. В 2007 году энтомологи выяснили, как именно осы заставляют тараканов идти за собой. Оса жалит два раза — первый раз она добивается уменьшения сопротивления, а второй раз она делает очень точный укол в мозг таракана. Это заставило учёных предположить, что жалящий яд может работать как блокатор ключевых химических сигналов в мозге. Исследование показало, что яд содержит блокатор нейромедиатораоктопамина.
Другим примером манипуляции поведeния хозяина является червь Leucochloridium paradoxum (из класса дигенетических сосальщиков (Digenea)), который паразитирует в кишечнике певчих птиц. Яйца с помётом птиц попадают на траву, а затем попадают в улитку Succinea, которая живёт во влажной среде. Яйца развиваются в личинки внутри этого промежуточного хозяина, а затем должны найти свой путь в пищеварительную систему подходящей птицы. Проблема здесь в том, что эти птицы не едят улиток, так что паразит должен заинтересовать своего будущего хозяина используя тактику «агрессивной мимикрии» — то есть используя какую-нибудь часть тела своего промежуточного хозяина в качестве «наживки». В теле улитки из яиц развивается личинка — мирацидий, превращающийся потом в спороцисту. Внутри спороцисты возникают мелкие личинки, напоминающие взрослого червя. Спороциста увеличивается в размерах, образуя выросты, которые распространяются по телу улитки. Когда один из выростов спороцисты попадает в щупальце улитки, он увеличивается в диаметре и становится ярко-зелёным. На выросте появляются тёмные круги, а на его конце — пятна. В таком виде он хорошо заметен снаружи сквозь кожный покров улитки. Вырост начинает совершать частые подёргивания. Благодаря раскраске и движению он становится похожим на гусеницу какой-нибудь бабочки. Паразит также влияет на поведение промежуточного хозяина: улитка движется к свету (которого она, как правило, избегает), тем самым увеличивая шансы быть замеченной птицей.
Род ос-наездников трибы Glyptapanteles паразитирует, в частности, на гусеницах Lymantria dispar, Chrysodeixis chalcites, Thyrinteina leucocerae. Самка откладывает в тело гусеницы около 80 яиц. Личинки наездника развиваются внутри хозяина, питаясь его гемолимфой, затем прогрызают себе выход наружу, закрепляются на близлежащей ветке или листе и плетут вокруг себя кокон. Но гусеница при этом не погибает, поскольку паразит выработал механизм использования хозяина для активной защиты личинок. В теле гусениц остаются одно или два яйца — «солдата-камикадзе», которые управляют хозяином в период окукливания остальных особей: всё ещё живая гусеница вместо того, чтобы продолжить своё обычное существование, остаётся на месте и выгибается дугой над коконами, охраняя их от хищников. Фактически гусеница остаётся живой на протяжении всей стадии окукливания ос. Почти одновременно с выходом из коконов взрослых насекомых хозяин погибает. При появлении клопов-щитников, которые питаются коконами ос, 17 из 19 заражённых пядениц начинают трясти головой во все стороны и, в конце концов, стряхивают хищника с ветки или обращают его в бегство. В то же время незаражённые особи не замечают щитника, даже если он забирается на них самих. В результате коконов остаётся почти в два раз больше, чем если бы они оказались без присмотра.
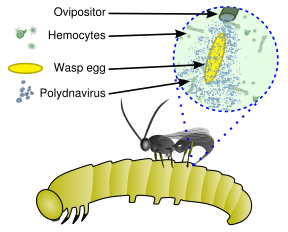
Диаграмма, показывающая ассоциацию паразита с поли-ДНК-вирусом
Ихневмоноидные наездники, личинки которых паразитируют на других насекомых, которые, в конечном счёте, погибают, выработали уникальный механизм контролирования поведения хозяина. Вместо яда многие виды впрыскивают в тело хозяина поли-ДНК-вирусы для подавления его иммунной системы. При этом геном вируса полностью встроен в геном паразита таким образом, что вирус не причиняет вреда паразиту и узко специализирован для подавления иммунитета хозяина, не размножаясь за пределами организма паразита.
Для современной науки появление вирусов на эволюционном древе жизни неясно: некоторые из них могли образоваться из плазмид, небольших молекул ДНК, способных передаваться от одной клетки к другой, в то время как другие могли произойти от бактерий. Существуют бактерии, использующие механизм передачи плазмид в целях инфицирования хозяина. Представители рода Agrobacterium[25] способны к горизонтальному переносу генов, при помощи которого вызывают опухоли у растений.
Аномальный рост этих корней (галлы) вызван бактерией Agrobacterium sp.
A. tumefaciens вызывает образование у растений злокачественных опухолей — галл. Такие опухоли возникают в результате конъюгационного переноса бактериальной Ti-плазмиды (Т-ДНК) в клетки растения. Близкородственный вид A. rhizogenes также вызывает корневые опухоли и обладает специальной Ri-плазмидой (англ.root-inducing — индуцирующая корни). Плазмидная T-ДНК полуслучайным образом внедряется в геном клетки хозяина[26], и происходит экспрессия генов, ответственных за образование опухоли, что в конечном итоге приводит к образованию галла. T-ДНК содержит гены, кодирующие ферменты, необходимые для синтеза нестандартных аминокислот, обычно октопина или нопалина. Здесь же закодированы ферменты для синтеза растительных гормонов ауксина и цитокина, а также для биосинтеза разного рода опинов, которые обеспечивают бактериям источник углерода и азота, недоступный для других микроорганизмов. Такая стратегия даёт Agrobacterium селективное преимущество[27]. Изменение гормонального баланса растения приводит к нарушению деления клеток и образованию опухоли.
Было доказано, что токсоплазма (лат.Toxoplasma — род паразитических протистов) может влиять на поведение хозяина: заражённые крысы и мыши меньше боятся кошек; замечены факты того, что заражённые крысы сами ищут места, где мочилась кошка. Этот эффект благоприятен для паразита, который сможет размножаться половым способом, если его хозяин будет съеден кошкой[28]. Механизм этого изменения до конца ещё не изучен, но существуют доказательства того, что токсоплазмоз повышает уровень дофамина у заражённых мышей.
Многие пухоеды птиц приносят минимальный ущерб своим хозяевам. Тем не менее в большинстве случаев может быть выявлен отрицательный эффект. Часто в основе этого эффекта лежит раздражение, вызывающее беспокойство и расчёсывания, что приводит косвенно к снижению общей жизнеспособности. Так, например, увеличение времени, необходимого для груминга, требует затрат времени и энергии, что, кроме прочего, приводит к повышению доступности для хищников более заражённых особей. Негативное воздействие инфекционных патогенных микроорганизмов обычно связано с их размножением в организме и воздействием продуктов их жизнедеятельности на хозяина.
Во многих случаях паразиты наносят серьёзный экономический ущерб скотоводам. Например, верблюжий клещ является одним из наиболее значительных эктопаразитов домашнего скота как в тропиках, так и субтропиках[29]. Этот паразит вызывает 65 % прямого ущерба и 35 % косвенного ущерба для скотоводства. Прямой ущерб включает потери в производстве молока, уменьшения массы и увеличения смертности животных, включая смертность от косвенного потребления акарицидов, а также потери кожевенной промышленности из-за проколов, вызванных этим паразитом. Косвенный ущерб связан с тем, что H. dromedarii действует как переносчик болезней для животных (dermatophilosis) и вызывает человеческие случаи геморрагической лихорадки в Саудовской Аравии. В Египте H. dromedarii является вторым наиболее распространённым клещом и рассматривается как один из важнейших распространителей протозойных болезней крови, тропического тейлериоза жвачных животных, вызванного Theileria annulata[30].
Адаптация к жизни в другом организме
Деревья, поражённые омелой.
Попав во внутреннюю среду хозяина, эндопаразит получает ряд преимуществ: богатство легко доступной для усваивания пищи, которая не требует перестройки процессов пищеварения. Например клеточный сок растений, кровь животных, содержимое их пищеварительного тракта, уже обработанное ферментами; защищённость от непосредственного воздействия абиотических и биотических факторов внешней среды. Поскольку все взаимодействии со сложными и изменчивыми окружающими условиями и врагами берёт на себя организм хозяина, паразиту обеспечена относительная стабильность условий существования. Внутренняя среда организма по ряду физико-химических факторов имеет высокую степень постоянства. В то же время организм как среда жизни создаёт для паразитов некоторые экологические трудности: ограниченность среды во времени и пространстве; трудность распространения от одной особи хозяина к другой; сложности в обеспечении кислородом; оборонительные реакции организма хозяина. Как преимущества, так и экологические трудности жизни в другом организме обусловили формирование у паразитов разнообразных адаптаций, позволяющих им эффективно размножаться и процветать.
Богатство легкодоступной пищи привело к упрощению системы пищеварения у паразитов. Например, у печеночного сосальщика пищеварительная система упрощена, а у бычьего цепеня и совсем утрачена. Всасывание питательных веществ у ленточных червей происходит всей поверхностью тела. Эволюция некоторых паразитов может приводить к потере метаболических функций[31]: метаболические процессы, которые не важны для выживания, утрачены, и готовые аминокислоты, нуклеотиды и углеводы получаются от хозяина[32].
У паразитических растений питание соками хозяина привело к редукции системы фотосинтеза и потери хлорофилла. Среди растений встречаются полные паразиты и полупаразиты. Паразиты не содержат хлорофилла, как, например, Повилика европейская, Заразиха, Раффлезия, Петров крест. Полупаразиты (например растение омела) имеют хлоропласты и берут от растения только минеральные вещества.
Омела белая имеет собственную хлорофиллоносную систему, благодаря чему она частично независима от хозяина, поэтому является полупаразитом.
Гаустории омелы белой: a = нижняя часть стебля омелы; i = главный корень омелы; f = корни омелы в коре дерева; g = почки омелы; e = поглотители; в d сечение древесины[33]Омела белая (сверху). Вид в поперечном разрезе на ветке гибридного чёрного тополя (Populus × canadensis).
Предпосылками для прорастания семян являются оптимальные температура, освещение и влажность. Семена прорастают весной одним или двумя выростами желтовато-зелёного цвета. В случае их контакта с перидермой растения-хозяина (около почек, черешков или листьев) внешний и расположенные под ним слои клеток перидермы буреют и постепенно теряют форму, становятся менее заметными, а впоследствии исчезают вследствие растворения пектина клеточных стенок перидермы специфическими ферментами[34].
Клетки выроста углубляются в перидерму перпендикулярно стеблю растения-хозяина. В первичной или вторичной коре от выроста в горизонтальном направлении начинают отходить боковые тяжи (первичные гаустории). В лубе (вторичная флоэма) из них образуется разветвлённая система сосущих гаусторий под зоной инфицирования. Гаустории радиально, сквозь луб и камбий, проникают к внешней поверхности вторичной ксилемы (сосудов, древесины). Часть клеток гаусторий соединяется с сосудами растения-хозяина, сквозь которые поглощается вода с растворёнными минеральными веществами — благодаря более высокому, чем у растения-хозяина, осмотическому потенциалу в ксилеме паразита. В связи с этим омеле присущ более высокий показатель транспирации, поддержанию которого способствует дополнительный механизм открытия устьиц. Так, у некоторых видов омелы показатели транспирации в десять раз выше, чем у растения-хозяина. С другой стороны, растения омелы имеют низкой водный потенциал даже тогда, когда растение-хозяин подвергается водному стрессу. Это позволяет омеле заселять довольно сухие экотипы.
Гаустории увеличиваются вместе со вторичным утолщением стебля хозяина благодаря наличию интеркалярной меристемы, активность которой синхронизирована с активностью меристемы хозяина. Таким образом, развитая сеть (эндофитная система) паразита является жизнеспособной довольно долго — от нескольких лет до десятилетий. Развитие экзофитной системы начинается после формирования сосущих гаусторий, первые побеги возникают в месте инфицирования. Типичным является развитие боковых побегов из дополнительных почек вдоль сосущих гаусторий, которые длительное время остаются жизнеспособными.
Открытым остаётся вопрос, следует ли считать представителей рода Подъельник растениями-паразитами. До недавнего времени считалось, что подъельник — микогетеротрофное сапрофитное растение, но организация его питания оказалась гораздо более сложной. Подъельник, как и большинство других представителей семейства Вересковые, живёт в симбиозе с микроскопическими грибами. Особенностями симбиоза у подъельника является то, что гифы одних и тех же грибов проникают как в корни подъельника, так и в корни расположенных рядом деревьев. Через эти гифы подъельник получает не только те питательные вещества, которые вырабатывают грибы, но и вещества от деревьев (например фосфаты), которые необходимы ему для нормального функционирования, в том числе формирования семян (именно по этой причине подъельник может обходиться без фотосинтезирующих частей); в обмен деревья получают посредством тех же грибных гифов избыток сахаров, вырабатываемых подъельником[35][36].
В отличие от свободноживущих организмов, которые сами взаимодействуют с окружающей средой, паразиты «переложили» заботу о регуляции отношений с окружающей средой на хозяина. Отсутствие у паразитов непосредственного контакта с факторами внешней среды привело также и к упрощению строения их нервной системы и редукции органов чувств. Исчезла и необходимость выработки приспособлений для активной и пассивной защиты от врагов.
Поскольку условия проживания паразитов постоянные и оптимальные, у них нет необходимости вырабатывать сложные механизмы адаптации. Это приводит к экономии расхода энергии на процессы, не связанные с поддержанием жизнедеятельности. Ограниченность размеров среды обитания паразитов компенсируется малыми размерами их тела и упрощением строения, а трудности в распространении — повышением способности к размножению. Большая плодовитость паразитов получила название «закона большого числа яиц»[37]. Например, человеческая аскарида за сутки способна отложить 250 тыс. яиц, а за 5-6 месяцев полового созревания — 50-60 млн яиц. Их суммарная масса в 1700 раз превышает массу самки. Интенсивное развитие половой системы обеспечивает высокие репродуктивные возможности вида. Этому также способствуют партеногенез (развитие без оплодотворения), полиэмбриония (когда с одного яйца появляется много зародышей), бесполое размножение (самки рожают только самок). Возникновение гермафродитизма (совмещение в одном организме мужской и женской половых систем) является гарантией полового размножения при наличии даже одной особи. Защита оплодотворённых яиц многослойными оболочками и обеспечение зародыша питанием способствуют выживанию потомства. Развитие приспособлений для выхода личинок из яйца и тела хозяина во внешнюю среду и их проникновение в организм нового хозяина способствуют расселению.
Благодаря смене хозяев в жизненном цикле паразита не допускается собирание в одном организме большого количества особей, что могло бы привести к гибели хозяина и утрате ценного для этого вида паразита ресурса.
Сложности в обеспечении кислородом привели к редукции дыхательной системы и перехода к анаэробному дыханию[38]. Поскольку их энергетические затраты невелики, а запасы пищи неисчерпаемы, то такой способ дыхания вполне оправдан.
Генетической основой эволюционных изменений, ведущих к упрощению уровня организации, являются мутации. Например, если сохранившиеся недоразвитые органы — рудименты, альбинизм (отсутствие пигментов) и другие мутации — не исчезают в процессе эволюции, то они встречаются у всех членов данной популяции.
Адаптационные механизмы эктопаразитов
Крючкообразные механизмы прикрепления на лапках вшиМиноги, присосавшиеся к речной рыбеIxodes holocyclus до и после питания кровью
Эктопаразиты выработали различные механизмы прикрепления к телу хозяина, такие как изменение формы тела: укорачивание и также сплющивание тела в дорсовентральном направлении[39]; сглаживание метамерии у сегментированных видов; редукция конечностей в крючкообразные механизмы прикрепления. Как правило, существует два основных типа прикрепительных аппаратов — крючкообразные выросты, которые могут различаться по их форме и расположению, и присоски, которые представляют собой различные ямки на теле, ограниченные мускулистым валиком. Иногда встречаются и другие способы прикрепления. У клещей это принцип прихлопывания двух створок. Встречается вариант обхватывающего крепления, когда особые выросты обхватывают участок тела хозяина кольцом, но без ущемления. Встречаются также распорки и стилеты. Также существуют различные нитеобразные структуры — стрекательные нити, липкие или прядильные нити, нитевидные отростки тела, стебельки.
Пищеварительная система также адаптируется, происходит усовершенствование строения ротовых аппаратов. У гематофагов вырабатываются специальные органы въедания в покровы хозяина, возникают приспособления к увеличению ёмкости кишечника. Имеется тенденция к упрощению или исчезновению кишечника.
Например, миноги (отряд бесчелюстных из класса круглоротых (Cyclostomata) Бесчелюстные) характеризуются отсутствием челюстей (однако у них имеется развитый череп, что отличает их от бесчерепных). В предротовой воронке миноги имеются роговые зубы. Мощный язык имеет крупный зуб, называемый тёркой. На дне воронки находится рот, который ведёт в глотку. Имеется печень и зачаточная поджелудочная железа. Миноги присасываются к рыбам и питаются кровью, тканевыми жидкостями и мышцами. Желудок не развит. Ферменты во рту миноги препятствуют свёртыванию крови жертвы. Круглоротые выделяют пищеварительные соки в тело жертвы (внекишечное пищеварение), а затем всасывают уже частично переваренную пищу. Жертвы обычно умирают от потери крови или инфекции.
Вирусы распространяются многими способами: вирусы растений часто передаются от растения к растению насекомыми, питающимися растительными соками, к примеру тлями; вирусы животных могут распространяться кровососущими насекомыми, такие организмы известны как переносчики. Вирус гриппа распространяется воздушно-капельным путём при кашле и чихании. Норовирус и ротавирус, обычно вызывающие вирусные гастроэнтериты, передаются фекально-оральным путём при контакте с заражённой пищей или водой. ВИЧ является одним из нескольких вирусов, передающихся половым путём и при переливании заражённой крови.
Первичным, или окончательным, хозяином называется хозяин, в котором паразит вырастает до зрелости; вторичным, или промежуточным, называется хозяин, который предоставляет среду обитания паразиту или симбионту только в течение короткого периода. Для трипаносом, возбудителей трипаносомозов, человек — окончательный хозяин, а муха цеце — промежуточный. Основные хозяева токсоплазм — представители семейства кошачьих. В качестве промежуточных хозяев выступают различные виды теплокровных животных, в том числе и люди. Токсоплазмоз (болезнь, вызываемая токсоплазмой) обычно протекает у человека легко. Однако для плода, в случае если мать заразилась токсоплазмозом во время беременности, а также для человека или кошки с пониженным иммунитетом эта болезнь может иметь серьёзные последствия, вплоть до летального исхода. Также существуют резервуарные хозяева, позвоночные животные, которые обеспечивают патогену как виду возможность непрерывного существования. В зависимости от естественного резервуара болезни человека делятся на антропонозы, где резервуаром является человек, и зоонозы, где резервуаром являются животные. Членистоногие хозяева, которые передают паразитирующий организм от одного хозяина к другому, называются переносчиками. Переносчиков в свою очередь подразделяют на механических, в организме которых возбудитель болезни не размножается (например мухи), и биологических, в организме которых протекает часть жизненного цикла возбудителя инфекции (комары, москиты). В последнем случае болезнь называется трансмиссивной.
В связи с паразитизмом у паразитов может быть сильно развита половая система, за счёт редуцирования пищеварительной и нервной системы. Максимальная плодовитость повышает возможность выживания паразитов, развивающихся со сменой хозяев. Жизненный цикл может состоять из нескольких этапов. Например, на первом этапе взрослые ленточные черви обитают в кишечнике окончательного хозяина, размножаются и продуцируют яйца. На втором этапе яйца попадают во внешнюю среду: в почву или в воду. На суше в яйцах формируется личинка или зародыш, представляющий фазу внедрения в промежуточного хозяина. У некоторых видов, яйца которых развиваются в воде, из яйца выходит свободноплавающая личинка, покрытая ресничками, а в ней формируется вторая личиночная фаза. На третьем этапе происходит развитие личинок в промежуточном хозяине, где развиваются в пузырчатую глисту — финну. Финна для дальнейшего развития должна попасть в кишечник основного хозяина, где головка из финны выворачивается, присасывается к стенке кишечника, после чего начинается процесс роста червя.
Овод лошадиный приклеивает яички на лету, по одному, на волосы передних ног, груди и шеи животного. Одна самка откладывает до 500 яиц. Через несколько дней вылупляются личинки, которые производят зуд на коже и тогда слизываются лошадью языком и попадают таким образом в её полость рта. Помимо того, личинки могут самостоятельно добираться до рта, цепляясь за волосы своими шипиками и крючками. С пищей личинки проходят в кишечник и там укрепляются с помощью своих ротовых крючков в слизистой оболочке глотки, пищевода или желудка, собираясь таким образом в одной лошади сотнями и даже тысячами. Каждая личинка обрастает на своём переднем конце опухолью прилежащих тканей, которая плотно охватывает при этом шипики её тела и прочно удерживает таким образом на месте в течение всей зимы. Весной вместе с испражнениями выпадает наружу. Окукливается в земле или в навозе и через месяц окрыляется.
Растения-паразиты
Растения-паразиты зачастую используют промежуточных носителей вкупе с производством огромного количества мелких семян. Так, например, в распространении омелы принимают участие птицы (эпизоохори́я, эндозоохория), преимущественно дрозды. Поедая её ягоды, они пачкают свой клюв клейкой ягодной массой — висцином, в котором находятся семена омелы. Затем, перелетая с дерева на дерево и очищая клюв о ветви, пачкают их этим клейким веществом, а также выбрасывают семена с испражнениями. Висцин на ветвях подсыхает, приклеивая к ним семена, которые через некоторое время начинают прорастать. Распространению омелы птицами способствуют небольшие размеры семян, их яркое и контрастное окрашивание. Сначала вытягивается его подсемянодольное колено, которое загибается к ветви и прикладывается кончиком корня к её поверхности; в это время кончик корня расширяется в кружок, при помощи которого молодое растеньице прикрепляется к дереву. После этого из центра кружка отходит небольшой стержневидный отросток, который прободает кору ветви и через её камбий дорастает до древесины. Этот отросток служит главной или первичной присоской; при помощи его омела вытягивает из растения воду и минеральные вещества. Как только присоска дорастает до древесины, её верхушечный рост прекращается, но по мере нарастания древесины она продолжает вытягиваться в длину при помощи вставочного роста, сосредоточенного в том участке присоски, который находится в камбии ветви. Древесина ветви обрастает присоску, так что в результате она оказывается внедрённой в древесину.
В Европе омелу распространяют свиристель (Bombycilla garrulus), различные виды дроздов — рябинник (Turdus pilaris) и деряба (Turdus viscivorus), а также славка-черноголовка (Sylvia atricapilla), для которых её плоды являются кормом. Птицы садятся преимущественно на верхние ветви разреженных крон старых деревьев, которые освещены и хорошо прогреваются, что необходимо для прорастания семян омелы. Расположение ветвей в кроне дерева, угол между ними и стволом предопределяют важные конкурентные параметры в отношениях «хозяин — паразит», которые положительно или отрицательно влияют на динамику повреждения омелой. Так, увеличение угла между веткой и стволом способствует потенциальному повреждению дерева паразитом, и наоборот — плотный лес, деревья в котором растут в высоту и не имеют развесистой кроны, практически свободен от поражения, очаги инфекции здесь нежизнеспособны и отмирают из-за недостатка света.
Сравнительно короткими расстояниями полёта птиц и быстрым прохождением семян через их желудочно-кишечный тракт объясняется формирование локальных очагов инфицирования и умеренное распространение паразита. Паразитирование омелы — типичное явление парковых лесов и аллей. Аллеи, поражённые омелой, могут выполнять функцию её распространения по принципу так называемых мостов. Распространение инфицирования на большие расстояния связано с перелётными птицами. Так, исследование повреждения омелой сосны чёрной, которая растёт на юго-западе Европы во французских Альпах, показало, что распространение полупаразита совпадало с основными курсами полёта дрозда-дерябы.
Предположительно, семенараффлезии, подобно семенам хорошо изученных растений-паразитов из семейства Заразиховые, пробуждаются к прорастанию под воздействием выделений потенциальных растений-хозяев. Эти же вещества ориентируют направление роста проростков. Развитие раффлезии происходит интраматрикально, то есть в тканях растения-хозяина. Тело растения-паразита при этом называется эндофитом, а такой способ паразитизма — эндопаразитизмом. В тканях растения-хозяина тело раффлезии распространяется в виде клеточных тяжей, напоминающих грибные гифы.
Недавние исследования показали, что повилика способна улавливать запах растений и таким образом находить жертву[40]. Хотя семена паразита могут лежать в земле больше пяти лет, семена невелики, собственных запасов питательных веществ в них хватает только на считанные дни, и без привлечения посторонних ресурсов росток вырастет не больше чем на 10 см. Почуяв запах потенциального хозяина, росток повилики приближается к нему и обвивает его стебель.
Паразиты используют и менее изощрённые способы поиска хозяина. Например, растения из подсемейства Ленноовые имеют корневые структуры двух типов. Направляющие корни довольно грубые, мясистые и не образуют гаустории, основная функция которых — поиск новых корней растений-хозяев. Когда направляющий корень приближается к корню растения-хозяина, на нём со стороны контакта возникают корнеподобные гаусториеобразующие органы, которые внедряются в ткани корня и устанавливают контакт с проводящей системой растения-хозяина[41].
Известен факт, что некоторые растения семейства Орхидных привлекают опылителей не пыльцой или нектаром, а воздействуя на их половые инстинкты. Поскольку в цветах этих орхидей отсутствует нектар, который мог бы быть атрагентом для потенциальных опылителей, то орхидеи пользуются структурами, напоминающими «самку», образуемую этими цветками[42][43][44]. Данное явление часто рассматривают в качестве примера высокоспециализированной энтомофилии[45]. Группа орхидей, цветки которых опыляются путём псевдокопуляциисамцами насекомых, распространена в Австралии. Примером перехода от нейтрализма к паразитизму служат взаимоотношения орхидеи Chiloglottis trapeziformis и осы Neozeleboria cryptoides[46]. Для этих ос характерен половой диморфизм: масса крылатых самцов около 16 мг (длина тела 9-12 мм; ширина головы 2 мм), самки бескрылые и значительно меньше, весят около 4 мг (длина тела около 6 мм, ширина головы 1 мм). Самец должен не только оплодотворить её, но ещё и перенести в место, пригодное для кладки яиц. Вместо этого самцы участвуют в опылении цветков орхидей Chiloglottis trapeziformis[47], которые по своей форме и запаху напоминают самок этих ос[48]. Орхидеи вырабатывают сходный с половым феромоном самок ос вещество 2-этил-5-пропилциклогексан-1,3-дион (по имени орхидеи названное Chiloglottones 1-6), которое привлекает самцов сильнее, чем их естественные партнёрши[49]. В результате самцы ос, не будучи способны отличить запах самки от запаха цветка, залетев в заросли орхидей, перелетают от одного растения к другому, не замечая самок, находящихся поблизости. Таким образом воспроизводству ос наносится определённый ущерб, и поэтому соответствие характера межвидовых взаимоотношений становится аналогично паразитизму.
Некоторые растения могут пахнуть разлагающейся падалью, чтобы заманить опыляющих насекомых, таких как мухи.
Бесклеточные
Бесклеточные — паразитарные организмы, не имеющие клеточного строения. Чаще всего к этой группе относят вирусы, однако некоторые ученые считают живыми и простые структуры, такие как вироиды, вирусоиды, плазмиды, транспозоны и прионы . Негативное воздействие инфекционных патогенных микроорганизмов обычно связано с их размножением в организме и воздействием продуктов их жизнедеятельности на хозяина.
В последние годы было предложено разделить все (живые) организмы на две группы: кодирующие рибосомы (англ. Ribosome encoding-organisms, REO, бактерии, археи и эукариоты) и те, которые кодируют капсиды (англ. Capsid encoding-organisms, CEO, вирусы). Это предложение подверглось критике. Существование неклеточных форм жизни является предметом научного спора в связи с тем, что чёткого и однозначного определения понятия «жизнь» пока нет. Многие учёные считают, что только клеточные организмы можно считать живыми.
Вирусы
Вирусы являются облигатными паразитами — они не способны размножаться вне клетки. Главная особенность — цитотропизм и облигатный внутриклеточный паразитизм вирусов, что делает их во всех отношениях (метаболически, энергетически и экологически) зависимыми от клетки-хозяина[50]. Вне клетки вирусные частицы не проявляют признаки живого и ведут себя как частицы биополимеров. От живых паразитарных организмов вирусы отличаются полным отсутствием основного и энергетического обмена и отсутствием сложнейшего элемента живых систем — аппарата трансляции (синтеза белка), степень сложности которого превышает таковую самих вирусов.
Вирусы — сборная группа, не имеющая общего предка. В настоящее время существует несколько гипотез, объясняющих происхождение вирусов. Считается, что крупные ДНК-содержащие вирусы происходят от более сложных (и, возможно, клеточных, таких как современные микоплазмы и риккетсии), внутриклеточных паразитов, утративших значительную часть своего генома. Обнаружены также вирусы, поражающие другие вирусы (вирусы-сателлиты). Они инфицируют клетки, для которых несвойственна естественная смерть от старости (например амёбы, бактерии). Когда клетку, заражённую вирусом-сателлитом, заражает обычный вирус, то вирус-сателлит нарушает производство вирионов этого вируса и сам начинает размножаться. Вирусы-сателлиты, по сути, являются гиперпаразитами.
Сверхпаразитизм среди вирусов отличается от прочих форм паразитизма тем, что вирусы не считаются живыми организмами и поэтому сверхпаразит живёт, как и первичный паразит, за счёт основного хозяина. Однако вторичные вирусы угнетают развитие первичных, что по сути дела определяет их как именно паразитов. Недавно были также обнаружены третичные паразиты вирусов. К примеру, в теле человека могут паразитировать амёбы рода Acanthamoeba. На этих амёбах паразитируют мимивирусы. На мимивирусах паразитируют вирофаги. А вирофаги содержат в себе паразитические генетические элементы — трансповироны[54].
Хламидия напоминает вирус тем, что является полностью внутриклеточной бактерией: зависит от питательных веществ и энергии клетки-хозяина, не синтезирует АТФ и, являясь энергетическим паразитом, полностью зависит от энергетических ресурсов клетки — то есть в процессе паразитирования разрушает её полностью. От всех других микроорганизмов хламидии отличаются совершенно особым жизненным циклом. Но, как и все бактерии, содержит как ДНК, так и РНК, размножается делением, имеет твёрдую оболочку наподобие грамотрицательных бактерий, которая содержит сходный липополисахарид.
Грибы
Хозяевами паразитических грибов чаще всего являются высшие растения, но ими могут быть и животные, а также грибы других видов. Для высасывания веществ из клетки хозяина на гифах паразитических грибов часто образуются гаустории, которые представляют собой боковые ответвления гифы, проникающие внутрь клетки хозяина. Паразиты проникают в тело хозяина через мелкие повреждения в его покровах, паразиты растений для этого используют естественные отверстия в эпидерме — устьица. В ходе роста гриб выделяет ферменты, разрушающие срединные пластинки между растительными клетками (пектиназы), в результате чего ткани размягчаются.
Hyaloperonospora parasitica: гифа с гаусториями
У мучнисторосяных грибов с поверхностным мицелием гаустории развиваются следующим образом: аппрессорий, прикрепляясь к клетке растения-хозяина, выделяет специфические ферменты, разрыхляющие клеточную кутикулу, и через разрушенные участки от основания аппрессориев выходит росток, внедряющийся в полость растительной клетки. В образовавшуюся гаусторию перетекает ядро. У ржавчинных грибов с межклеточным мицелием гаустории являются продолжением вегетативных гиф, которые, проникнув в клетку хозяина, изменяют свой внешний вид.
Для растительной клетки гриб является инородным телом, внедрение которого не проходит бесследно: клетка растения реагирует на присутствие гриба образованием каллозного чехла, препятствующего дальнейшему росту гаустории. Гаустории состоят из трёх частей: материнской грибной клетки, шейки гаустории — части, пронизывающей клеточную стенку, и собственно гаустории, расположенной внутри клетки хозяина. Иногда из одной материнской клетки может прорастать несколько гаусторий. В гаусториях, как правило, наблюдается большое число митохондрий и рибосом, хорошо развит эндоплазматический ретикулум, что является показателем того, что в гаустории активно проходят физиологические процессы. От клетки-хозяина гаустория обычно отделена инвагинацией (впячиванием) плазмалеммы клетки-хозяина. Между клеточной стенкой гаустории и плазмалеммой клетки-хозяина образуется аморфный слой (капсула), через который осуществляются все обменные процессы между хозяином и паразитом. В клетке организма-хозяина идут активные процессы синтеза разнообразных веществ, часть которых поступает в гаусторию, а часть идёт на формирование чехла, изолирующего гаусторию от клетки, в которой она находится.
Некоторые паразиты ограничиваются тем, что постепенно высасывают вещества из хозяина, но не приводят его к гибели, другие же выделяют ферменты, расщепляющие целлюлозуклеточной стенки, что приводит к гибели клетки-хозяина, после чего паразит питается органическими остатками хозяина. Некоторые грибы становятся паразитами только в определённых случаях. Например, известны виды, являющиеся сапротрофными, но при этом способные поселяться на ослабленных организмах (обычно высших растениях), переходя на паразитический образ жизни. Когда хозяин погибает, они продолжают жить на нём как сапротрофы, поглощая органические вещества того организма, на котором они недавно паразитировали. Грибы, способные вести исключительно паразитический образ жизни, называют облигатными паразитами. Формы, которые способны периодически менять образ жизни с сапротрофного на паразитический, называют факультативными паразитами. При этом облигатные паразиты обычно не приводят к смерти хозяина, поскольку для них это тоже будет губительно, тогда как деятельность факультативных паразитов чаще всего убивает хозяина, но они в дальнейшем могут жить и на мёртвых остатках.
Эпидемиологические и экологические аспекты паразитизма
no data ≤100 100-200 200-300 300-400 400-500 500-600 600-700 700-800 800-900 900-1000 1000-1500 ≥1500
Советский плакат призывающий к борьбе с тифозной вошью.
На млекопитающих и на человеке (волосы, кожа), в его организме (кишечник, печень, мозг, лёгкие, кровь, половые органы) паразитируют аскариды, трихинеллы, лентецы, острицы; печёночный сосальщик, цепни, вши, блохи, малярийный плазмодий, чесоточный зудень, клещи, трихомонада, различные бактерии, патогенные грибы, вирусы и др. Болезни, которые вызваны этими паразитами, весьма многочисленны. Таковыми являются: аскаридоз, трихинеллёз, трихомоноз, чесотка, сифилис и др. Хотя паразитарные болезни (группа заболеваний, вызываемых паразитами) являются частью инфекционных, обычно их разделяют и рассматривают инфекционные болезни как заболевания, вызванные одноклеточными организмами (вирусы, бактерии, грибы, простейшие). Возбудители паразитарных болезней — многоклеточные животные — различные гельминты и членистоногие. Клиника паразитарных болезней проявляется в широком диапазоне от субклинической до тяжёлой. Некоторые паразитозы не оказывают фактического влияния на жизнь и здоровье человека, другие нередко угрожают жизни (трихинеллёз, эхинококкоз, малярия) и значительно ухудшают здоровье (филяриатозы, шистосомозы). Вши и блохи также являются переносчиками опасных болезней человека и животных (сыпной тиф, чума).
Известные пандемии чумы, унёсшие миллионы жизней, оставили глубокий след в истории всего человечества. Кровососущие насекомые (комары, москиты, кровососущие мухи) также переносят инфекцию во время сосания крови. Так, например, осенняя жигалкаStomoxys calcitrans часто нападает и на человека, нанося болезненные укусы, и является переносчиком возбудителей сибирской язвы, сепсиса, туляремии, трипаносомозов и других заболеваний энтомозов. По последним оценкам ВОЗ, в год происходит от 124 до 283 миллионов случаев заражения малярией и от 367 до 755 тысяч смертей от заболевания. По данным ВОЗ около 10 % людей на Земле больны амёбиазом. В мире амёбная дизентерия является второй по важности причиной смертности от паразитарных заболеваний[55]. Всего на долю паразитарных заболеваний приходится около 14 млн смертей в год, что составляет 25 % от общемирового показателя смертности — каждая четвёртая смерть, по данным ВОЗ[56][57].
В 2010 году сонная болезнь (африканский трипаносомоз), переносимая мухой цеце, вызвала 9 тыс. смертей; уровень смертности понизился по сравнению с данными 1990 года (34 тыс. смертельных случаев)[58]. По оценкам, в настоящий момент заражено около 30 тыс. человек, из них 7 тыс. были заражены в 2012 году[59]. Другие животные, например коровы, могут переносить эту болезнь и заражаться ей. В начале XX века после сокращения поголовья рогатого скота от африканской чумы снизилось и заболевание сонной болезнью. Это послужило поводом к уничтожению миллионов голов диких животных. В первой половине XX века было обнаружено, что численность мухи цеце выше в лесистых районах. Основной мерой борьбы в тот период стала вырубка кустарников. С 1940-х годов для уничтожения мухи цеце применялись инсектициды (ДДТ)[60]. Однако именно наличие большого количества мух цеце спасло большую часть Африки от перевыпаса и эрозии почв, обычно вызываемой крупным рогатым скотом.
Источником возбудителя инвазии является больной, либо паразитоноситель (человек или животное) — хозяин паразита. При некоторых паразитарных болезней хозяин паразита может служить источником инвазии для самого себя (например повторное заражение острицами при привычке грызть ногти, под которыми у людей, больных энтеробиозом, могут быть яйца остриц).
Высокая заболеваемость населения тропических регионов имеет несколько причин, но главным образом имеет экономическую базу — перенаселённость этих стран и бедность населения. Группа инфекционных и паразитарных тропических заболеваний, поражающих преимущественно беднейшие и маргинализированные слои населения в наиболее отсталых регионах Азии, Африки и Латинской Америки, известна как «забытые болезни». В основной список Всемирной организации здравоохранения входит 17 заболеваний[61], из которых семь вызываются паразитирующими червями, три — протозойными паразитами и ещё три — бактериями. Это болезни с наиболее высокой заболеваемостью. Ещё двадцать болезней, также относящихся к забытым, вызываются грибками, вирусами и эктопаразитами.
Семь болезней из обоих списков характеризуются тем, что методы их профилактики и лечения известны, но недоступны в беднейших странах, где они наиболее распространены[62].
Забытые болезни поражают более миллиарда человек и вызывают около полумиллиона смертей ежегодно[63].
До половины населения мира инфицировано токсоплазмозом[64]. В США носителями являются 23 % населения[65], в России — около 20 %[66], а в некоторых районах мира доля носителей достигает 95 %[67].
Глобальная ежегодная заболеваемость врождённым токсоплазмозом оценивается на уровне 190 тыс. случаев. Высокие уровни заболевания были отмечены в Южной Америке, некоторых ближневосточных странах и странах с низким уровнем доходов[68]. При врождённом токсоплазмозе наблюдаются гибель плода в утробе матери, смерть новорождённого в результате общей инфекции или (у оставшихся в живых) поражение нервной системы, глаз и других органов.
Многие паразитоиды (напр. тахины или наездники) — естественные враги многих насекомых, регулирующие численность последних и приносящие таким образом большую пользу. Некоторые виды успешно акклиматизированы в разных странах для борьбы с колорадским и японским жуками, непарным шелкопрядом и другими вредителями. На клопах паразитируют Phasia crassipennis и Gymnosoma dolycoridis. В тридцатых годах удалось справиться с красной кровяной тлёй (Eriosoma lanigerum) при помощи колонизации наездника афелинуса.
Микроспоридии[70], паразитирующие у насекомых-вредителей сельскохозяйственного, медицинского и ветеринарного значения, являются перспективными агентами биологической борьбы с этими видами. Некоторые виды микроспоридий комаров вызывают высокую смертность личинок, снижают плодовитость самок, а также предотвращают развитие нескольких видов малярийных плазмодиев[71]. В 1978 году в США для борьбы с прямокрылыми вредителями был создан первый промышленный биопрепарат на основе микроспоридий — «Нолок», который получил высокую экономическую оценку благодаря низкой стоимости, высокой эффективности и отсутствию негативного влияния на окружающую среду[71].
Известны также случаи немедицинского заражения гельминтами под предлогом похудения, хотя такая практика является опасной для здоровья и жизни[73][74].
После открытия явлений бактериофагии одним из направлений онкологии является виротерапия — один из видов биотерапии, при котором применяются онкотропные/онколитические вирусы. Виротерапия мобилизует естественные защитные силы иммунной системы организма против клеток генетически модифицированных организмов и тканей, в том числе злокачественных клеток. В нечувствительных опухолях вирус способен модулировать структуры опухолевых клеток, делая их иммуногенными. В результате вирус не только причиняет прямой ущерб раковым клеткам, но и включает в борьбу с опухолью собственную иммунную систему больного раком. В силу этого виротерапия является также методом активизации антигенспецифических механизмов иммунной защиты. Лучшие результаты достигнуты на ранних стадиях опухоли, когда виротерапия применяется перед и после радикальной операции для профилактики метастазов. В Латвии в 2004 году был зарегистрирован первый генетически немодифицированный энтеровирус Ригвир (RIGVIR) для лечения меланомы. Последнее опубликованное исследование показывает, что у пациентов с меланомой, получавших Ригвир, вероятность выжить была выше, чем у тех, кто не получал никакой терапии. Данный показатель составил от 4,39 до 6,57 раз[75].
В 2005 году в Китае был зарегистрирован генетически модифицированный аденовирус H101 (позже известный как Oncorine) для лечения злокачественных опухолей головы и шеи[76].
Д’Эрелль развил учение о том, что бактериофаги патогенных бактерий, являясь их паразитами, играют большую роль в патогенезе инфекций, обеспечивая выздоровление больного организма, а затем создание специфического иммунитета. Это положение привлекло к явлению бактериофагии внимание многих исследователей, которые предполагали найти в фагах важное средство борьбы с наиболее опасными инфекционными болезнями человека и животных.
Одной из областей использования вирусов бактериофагов является антибактериальная терапия, альтернативная приёму антибиотиков Например, применяются бактериофаги: стрептококковый, стафилококковый, клебсиеллёзный, дизентерийный поливалентный, пиобактериофаг, коли, протейный и колипротейный и другие. В России зарегистрировано и применяется 13 медицинских препаратов на основе фагов[77]. В настоящее время их применяют для лечения бактериальных инфекций, которые не чувствительны к традиционному лечению антибиотиками, особенно в республике Грузия[78][79][80]. Обычно применение бактериофагов сопровождается большим, чем антибиотики, успехом там, где присутствуют биологические мембраны, покрытые полисахаридами, через которые антибиотики обычно не проникают[81]. В настоящее время терапевтическое применение бактериофагов не получило одобрения на Западе, хотя и применяются фаги для уничтожения бактерий, вызывающих пищевые отравления, таких как листерии[82]. В России терапевтические фаговые препараты делают давно, фагами лечили ещё до антибиотиков. В последние годы фаги широко использовали после наводнений в Крымске[83] и Хабаровске, чтобы предотвратить дизентерию.
Одной из самых известных характеристик рода грамотрицательных бактерий Bdellovibrio является способность его представителей паразитировать на других грамотрицательных бактериях, входя в их периплазменое пространствo и питаясь биополимерами, например белками и нуклеиновыми кислотами хозяев. После входа в периплазменое пространство хозяина бактерия Bdellovibrio формирует структуру, известную как бделопласт, состоящий как из структур паразита, так и с структур хозяина. Клетка паразита может остаться бездеятельной на этой стадии, не мешая жизнеспособности хозяина. В большинстве случаев, однако, Bdellovibrio пожирает свою добычу и идёт дальше. Клетка Bdellovibrio использует гидролитические ферменты для разрушения молекул клетки хозяина, которые она использует для создания нити из нескольких новых клеток Bdellovibrio. Когда питательные вещества клетки хозяина истощены, нить отделяется, формируя потомков Bdellovibrio. Потомки становятся подвижными перед тем, как они разрушат клетку хозяина и выходят из окружения. Полный жизненный цикл занимает от одного до трёх часов и приводит к образованию 3-6 клеток-потомков из единой клетки E. coli. Британские учёные из университета Ноттингема и Имперского колледжа провели исследование, в результате которых им удалось использовать Bdellovibrio bacteriovorus как живой антибиотик для борьбы с инфекцией Шигелла, от которой ежегодно умирает около одного миллиона человек. Введение бактерии существенно уменьшалo количество бактерий Шигелла. После успешных опытов на рыбе будут возможны эксперименты на организме человека[85].
Палеопатология
Реконструкция особи тираннозавра, заражённой паразитами, основанная на экземпляре MOR 980
Исследование 2009 года показало, что дыры, обнаруженные в черепе у некоторых экземпляров тираннозавров, которые прежде считали ранами, полученными в межвидовых конфликтах, на самом деле были вызваны паразитами вроде трихомонады — простейшего, которое обычно поражает птиц[86]. Эти паразиты способны не только поражать мягкие ткани, но и делать в костях рубцы изнутри. Возможно, паразиты передавались через воду, которую пили заражённые динозавры.
В массовой культуре
Тема паразитизма очень популярна. В XIX веке стал популярен образ вампира, возможно позаимствованный от восточноевропейских «упырей». В современной фантастике образ инопланетных паразитов, захватывающих человека-хозяина, повторялся множество раз. Такие фильмы, как «Чужой» и «Нечто», показывают инопланетян как классических паразитов с личиночным размножением. Другие фильмы, например «Кукловоды» или «Звёздные врата», показывают более умеренную форму сосуществования между хозяином и паразитом.
Тема невымышленных историй столкновения людей из США с паразитами была поднята американскими документалистами в серии телепередач Монстры внутри меня[англ.]. Каждый эпизод описывает историю заражения обычных американцев той или иной формой экзотического паразитического патогена.
Роль паразитов в различных экосистемах и их роль в эволюции стала темой научно-популярной книги Карла Циммера «Паразит — царь природы: Тайный мир самых опасных существ на Земле», заслужившей высоких похвал от специалистов [2].
Интересные факты
Чарлз Дарвин считал, что существование наездников несовместимо с центральным понятием естественной теологии, рассматривающей изучение природы как путь к демонстрации благожелательности Бога. В письме к американскому ботанику А. Грею Дарвин писал: «Я не могу убедить себя в том, что благодетельный и всемогущий Бог мог бы намеренно создать ихневмонид, чтобы они питались телами живых гусениц, или что кот должен играть с мышью».
У некоторых семейств рыб-удильщиков (Caulophrynidae, Ceratiidae, Neoceratiidae и Linophrynidae) существуют крайне необычные отношения между полами, не встречающиеся среди других рыб и позвоночных животных: Гренландская цератия (Cryptopsaras holboelli). Самка с крошечным самцом-паразитом. Препарат из Музея зоологии в Санкт-Петербурге. карликовые самцы живут в виде «паразитов» на теле самок. До перехода к паразитизму самцы обладают хорошо развитыми глазами и крупными обонятельными органами. Это позволяет им отыскивать самок по запаху феромонов, следы которого сохраняются в практически неподвижной воде больших глубин долгое время. Приблизившись к самке, самец, видимо, визуально распознает её видовую принадлежность по строению или по цвету и частоте её вспышек. Затем самец прицепляется к боку самки своими острыми зубами. Вскоре он срастается с самкой губами и языком, а его челюсти, зубы, глаза и кишечник редуцируются так, что в конце концов он превращается в простой придаток, вырабатывающий сперму. Питание самца осуществляется за счёт крови самки, так как их кровеносные сосуды тоже срастаются. На одной и той же самке может одновременно паразитировать до трёх самцов. Однажды прикрепившись, они полностью утрачивают самостоятельность. Биологическое значение этого явления, по-видимому, связано с облегчением нахождения полами друг друга во время размножения и с ограниченностью пищи на больших глубинах, и поэтому формально оно не должно рассматриваться как истинный паразитизм, а как специфическая форма социального. Учёные назвали такое поведение «сексуальным паразитизмом»[87].
См. также
Гельминтология — наука о паразитических червях и заболеваниях, вызываемых ими у человека и животных, — гельминтозах.
Мистель (нем. Mistel — «Омела»), также известен как «Папа и сын» (нем. Vati und Sohn) — авиационный комплекс военного назначения, созданный в конце Второй мировой войны.
Примечания
↑в более широком научном понимании симбиоз является любой формой взаимодействия между организмами разных видов, в том числе паразитизм — отношения, выгодные одному, но вредные другому симбионту.
↑Rozsa, L.; Garay, J. (2023). "Definitions of parasitism, considering its potentially opposing effects at different levels of hierarchical organization". Parasitology. 150 (9): 761—768. doi:10.1017/S0031182023000598. PMID37458178.
↑Паразитизм не обнаружен у мхов, и папоротникообразных растений
↑обыкновенной мясной, комнатной и других, что может вызывать такие болезни как например Кишечный миаз или схожие заболевания как например Сколециаз- энтомоз кишечника, вызванный личинками чешуекрылых
↑Dawson J. H., Musselman L. J., Wolswinkel P., Dörr I.Biology and control of Cuscuta // Reviews of Weed Science. — 1994. — Vol. 6. — P. 265–317. Архивировано 22 июля 2015 года.
↑Francis, Kirk E.; Spiker, Steven. Identification of Arabidopsis thaliana transformants without selection reveals a high occurrence of silenced T-DNA integrations (англ.) // The Plant Journal[англ.] : journal. — 2004. — Vol. 41, no. 3. — P. 464—477. — doi:10.1111/j.1365-313X.2004.02312.x. — PMID15659104.
↑Pitzschke, Andrea; Hirt, Heribert. New insights into an old story: Agrobacterium-induced tumour formation in plants by plant transformation (англ.) // The EMBO Journal : journal. — 2010. — Vol. 29, no. 6. — P. 1021—1032. — doi:10.1038/emboj.2010.8. — PMID20150897. — PMC2845280.
↑Berdoy M, Webster J, Macdonald D (2000). Fatal Attraction in Rats Infected with Toxoplasma gondii. Proceedings of the Royal Society of London, B267:1591-1594. полный текст (англ.) (недоступная ссылка)
↑Среди хозяев кроме верблюдов (Camelus dromedarius) также отмечены крупный рогатый скот, овцы, козы и лошади
↑Метаболизм обычно делят на две стадии: в ходе катаболизма сложные органические вещества деградируют до более простых; в процессах анаболизма с затратами энергии синтезируются такие вещества, как белки, сахара, липиды и нуклеиновые кислоты
↑Lawrence J. Common themes in the genome strategies of pathogens (англ.) // Curr Opin Genet Dev : journal. — 2005. — Vol. 15, no. 6. — P. 584—588. — doi:10.1016/j.gde.2005.09.007. — PMID16188434.Wernegreen J. For better or worse: genomic consequences of intracellular mutualism and parasitism (англ.) // Curr Opin Genet Dev : journal. — 2005. — Vol. 15, no. 6. — P. 572—583. — doi:10.1016/j.gde.2005.09.013. — PMID16230003.
↑по Julius Sachs: Vorlesungen über Pflanzenphysiologie, zweite Auflage, Leipzig 1887
↑Кохно М. А. К биологии омелы = До біології омели (укр.). — К.: Изд-во АН УССР, 1960. — 32 с.
↑Кожевников Ю. П.Семейство вересковые (Ericaceae) // Жизнь растений. В 6-ти т. / под ред. А. Л. Тахтаджяна. — М.: Просвещение, 1981. — Т. 5. Ч. 2. Цветковые растения. — С. 88—95. — 300 000 экз.
↑Виар, Мишель. Цветы мира / Пер. с фр. В. А. Чекмарёв. — Изд. 2-е, стереотипное. — М.: Интербук-бизнес, 2001. — С. 58. — ISBN 5-89164-089-9.
↑Среди животных анаэробное фумаратное дыхание встречается у некоторых паразитических и свободноживущих червей, ракообразных, моллюсков; нитратное дыхание известно среди грибов (например, у Fusarium) [1]Архивная копия от 23 июля 2014 на Wayback Machine
↑Walsh J. Prevalence of Entamoeba histolytica infection. In: Ravdin JI, ed. Amebiasis: human infection by Entamoeba histolytica. New York: John Wiley and Sons, 1988: 93-105.
↑Lozano, R. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: a systematic analysis for the Global Burden of Disease Study 2010 (англ.) // The Lancet : journal. — Elsevier, 2012. — 15 December (vol. 380, no. 9859). — P. 2095—2128. — doi:10.1016/S0140-6736(12)61728-0. — PMID23245604.
↑— группа родственных грибампростейших, все представители которой являются облигатными внутриклеточными паразитами эукариотических организмов.
↑ 12Соколова Ю. Я., Исси И. В. Энтомопатогенные простейшие и особенности патогенеза протозойных заболеваний насекомых // Патогены насекомых: структурные и функциональные аспекты / под ред. В. В. Глупова. — М.: Круглый год, 2001. — С. 76—188.
Циммер К. Паразиты: Тайный мир = Parasite Rex: Inside the bizarre world of natures most dangerous creatures / Карл Циммер; Пер. с англ. Н. И. Лисовой. — М.: Альпина нон-фикшн, 2011. — 368 с. — 5000 экз. — ISBN 978-5-91671-081-6, ISBN 978-0-684-85638-4.
Органи́зм — живое тело, обладающее совокупностью свойств, отличающих его от неживой материи, в том числе обменом веществ, самоподдерживанием своего строения и организации, способностью воспроизводить их при размножении, сохраняя наследственные признаки. Термин организм введён Аристотелем. Он выявил, что любое живое существо характеризуется чёткой и строгой организацией, в отличие от неживого.
Вируле́нтность — степень способности данного инфекционного агента вызывать заболевание или гибель организма. Вирулентность является мерой патогенности.
Биотические факторы — формы воздействия организмов друг на друга, как внутри вида, так и между различными видами.
Симбио́з — это близкое сообщество живых организмов, принадлежащих к разным биологическим видам. Такое сообщество может принимать различные формы в зависимости от природы отношений между двумя видами и от того, полезны эти отношения или вредны.
Инфе́кция — заражение живого организма инфекционным агентом, процесс его размножения или развития в организме, а также реакция организма на присутствие в тканях инфекционного агента и на выделяемые им токсины. Под инфекционными агентами обычно понимают вирусы, микроорганизмы, прионы, а также черви-паразиты (гельминты). В случае бактерий инфекция может быть вызвана бактериофагами. Если в результате инфекции развивается заболевание, то такая инфекция называется инфекционным заболеванием. Инфекция подразумевает различные виды взаимодействия чужеродных микроорганизмов с организмом человека, животных или растений.
Грибы́ — царство живой природы, объединяющее эукариотические организмы, сочетающие в себе некоторые признаки как растений, так и животных. Грибы изучает наука микология, которая считается разделом ботаники, поскольку ранее грибы относили к царству растений.
Немато́ды, или кру́глые че́рви, — тип первичноротых из группы линяющих. В настоящее время описано более 24 тыс. видов паразитических и свободноживущих нематод, однако оценки реального разнообразия, основывающиеся на темпах описания новых видов, предполагают существование около миллиона видов.
Патоге́н — любой микроорганизм, а также особый белок — прион, способный вызывать патологическое состояние (болезнь) другого живого существа. В более общем случае под патогеном понимают любой фактор внешней среды, способный вызвать повреждение каких-либо систем организма или развитие каких-либо заболеваний. Патогенными микроорганизмами называются паразитирующие микроорганизмы по отношению к их хозяину.
Гаусто́рии — боковые ответвления гиф паразитических грибов, проникающие внутрь клетки-хозяина. Образование гаусторий паразитическими грибами является единственным способом получения грибом питательных веществ, которые всасываются гаусториями из клеток хозяина. Впервые обнаружены немецким фитопатологом А. де Бари. Гаусториями также называются аналогичные образования других организмов, например полупаразитического кустарника омела.
Оме́ла бе́лая — кустарник; типовой вид рода Омела семейства Омеловые (Viscaceae). Полупаразит: воду и минеральное питание получает от растения-хозяина, а органическое вещество фотосинтезирует самостоятельно. Имеет форму шара, прикреплённого к ветвям растения-хозяина.
Паразитоид — организм, который проводит значительную часть своей жизни, проживая на или внутри своего единственного хозяина, которого он постепенно убивает в процессе поедания. Таким образом, они подобны паразитам, за тем исключением, что паразиты не убивают хозяина. Взрослые стадии паразитоидов (имаго) — свободноживущие организмы.
Гнездовой паразитизм — тип клептопаразитизма, которым пользуются некоторые птицы, рыбы и насекомые, заключающийся в манипуляции и использовании другого животного-хозяина, или того же вида, или другого (межвидовой) для выращивания потомства животного-паразита.
Парази́ты — экологическая группа растений, получающих питательные вещества непосредственно из тканей других растений. В подавляющем большинстве случаев паразитизм известен среди представителей отдела покрытосеменных (Magnoliophyta), гораздо реже, в виде исключения — среди хвойных (Pinophyta), а также у низших растений — зелёных водорослей (Chlorophyta). Связь с растением-хозяином паразит осуществляет через гаустории, возникающие в результате преобразования зародышевого корня или, в редких случаях, тканей стебля.
Дициеми́ды(лат. Dicyemida) — тип примитивно устроенных беспозвоночных животных, паразитирующих на головоногих моллюсках. Содержит 70—75 видов. Ранее дициемид из-за простоты организации объединяли с ортонектидами в группу Mesozoa, рассматривая в качестве переходной группы между протистами и многоклеточными животными. Впоследствии, когда получила широкую поддержку гипотеза об их вторичном упрощении в связи с паразитическим образом жизни, эти представления были отвергнуты.
Сверхпаразити́зм, гиперпаразитизм — один из видов паразитизма, характеризующийся паразитированием одного паразита в другом. В этом случае сверхпаразит называется паразитом второго порядка, а его хозяин — паразитом первого порядка. Паразитизм более высоких порядков встречается очень редко. Один из примеров — заражение наездником Asecodes albitarsus многоядного сверхпаразита Dibrachys boucheanus, который, в свою очередь, нередко поражает наездников Apanteles glomeratus, паразитирующих на гусеницах белянок. Сверхпаразитизм достаточно широко распространён среди паразитических насекомых, в частности у наездников семейств Ichneumonidae, Encyrtidae и некоторых других. Однако сверхпаразитизм встречается и в других группах животных. Некоторые трематоды живут в ракообразных — эктопаразитах морских рыб. Сверхпаразитизм встречается и у некоторых ракообразных семейства тантулокарид. Обнаружено также как минимум одно семейство вирусов, поражающее другие вирусы. Гиперпаразитами называют и некоторые виды у грибов.
Микроспори́дии — клада родственных грибам простейших, все представители которой являются облигатными внутриклеточными паразитами эукариотических организмов. Описано около 1300 видов в 160 родах, что является малой частью реального разнообразия данной группы, так как огромное количество потенциальных хозяев не было исследовано на предмет заражения микроспоридиями. Данные патогены широко распространены среди животных практически всех систематических групп, от простейших до высших позвоночных, включая человека. Наиболее многочисленны и разнообразны микроспоридии ракообразных и насекомых.
Хозя́ин — организм, содержащий внутри вирус, паразита или симбиотического партнёра, обычно обеспечивая его питанием и убежищем. Например, клетка может быть хозяином для вируса, растение семейства бобовых может быть хозяином для полезной бактерии-диазотрофа, животное может быть хозяином для паразитического червя — нематоды.
Тафри́на — род грибов-аскомицетов, относящийся к монотипному семейству Тафри́новые. Все представители рода — паразиты высших растений, вызывающие различные повреждения листьев, побегов и плодов. Заболевания, вызываемые этими грибами, могут наносить урон сельскохозяйственным растениям — в частности, плодовым деревьям, а также лесным и другим насаждениям.
Паразиты человека — это паразиты, заражению которыми подвержен человек. Общее определение слова «паразит» касается не только многоклеточных животных и простейших, живущих за счёт своего хозяина и во вред последнему, но также вирусов, бактерий и грибов. Согласно исторической традиции, и с точки зрения медицинской паразитологии, паразитами принято называть любые существа, ведущие паразитический образ жизни, кроме вирусов и бактерий с подобными качествами.
Арахноэнтомозы — паразитарные болезни животных, культурных растений и человека, вызванные членистоногими.
Эта страница основана на статье Википедии. Текст доступен на условиях лицензии CC BY-SA 4.0; могут применяться дополнительные условия. Изображения, видео и звуки доступны по их собственным лицензиям.