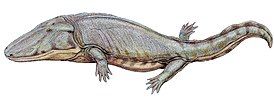
Мастодонзавры — род гигантских темноспондильных амфибий триасового периода.

Темноспонди́льные, или расчленённопозвонковые (лат. Temnospondyli, от др.-греч. τέμνειν — резать и σπόνδυλος — позвонок) — в высшей степени разнообразная группа вымерших животных, включающая от маленьких до гигантских примитивных земноводных. Название группы отсылает к тому, что каждый позвонок темноспондильных разделён на несколько частей.

Тероцефалы (лат. Therocephalia) — клада терапсид, традиционно рассматриваемая в ранге подотряда или отряда. Очень примитивные териодонты, возможно, родственные горгонопсам с одной стороны и цинодонтам — с другой. Вместе с цинодонтами образуют группу эутериодонтов (Eutheriodontia).

Платиопозавр (Platyoposaurus) — род пермских темноспондилов. Представитель семейства Archegosauridae, подсемейства Platyoposaurinae. В старой литературе известен как платиопс (Platyops). Отличается длинной, резко суженной ростральной частью черепа, с ложковидным озубленным расширением на концах челюстей. Наружные ноздри щелевидные, плохо заметны на ископаемых черепах, сдвинуты назад. Ушные вырезки узкие. В заглазничной части черепа бугры на таблитчатых костях и впереди ушных вырезок. Зубы довольно крупные, особенно на концах челюстей. Тело массивное, коренастое, уплощённое, примерно 22 предкрестцовых позвонка. Сильно развиты крючковидные отростки туловищных рёбер. Брюхо покрыто мелкими удлинённо-овальными чешуями, спинных чешуй не обнаружено. Конечности довольно длинные, особенно задние. Высокие остистые отростки, особенно на основании хвоста. Хвост не очень длинный, уплощённый с боков. В целом животное напоминало небольшого длинномордого крокодила. Вероятно, рыбоядный хищник, скорее всего постоянноводный.

Крассигирин — примитивный стегоцефал начала каменноугольной эпохи, единственный вид в роде Crassigyrinus и семействе Crassigyrinidae.

Капториниды (лат. Captorhinidae) — семейство примитивных пресмыкающихся верхнего палеозоя. Традиционно относились к так называемым анапсидным рептилиям, лишённым височных окон. По последним исследованиям, стоят ближе к диапсидным рептилиям, чем к другим анапсидам. С капторинидами сближают также древнейших известных амниот — семейство Protorothyrididae, самые известные представители которых: гилономус и палеотирис. По последним данным, проторотиридиды находятся в основании ствола диапсид.

Ринезухиды (лат. Rhinesuchidae) — семейство вымерших земноводных из подотряда стереоспондильных отряда темноспондильных, живших во времена окончания пермского — начала триасового периодов в основном на территории Южной Африки. Занимают промежуточное положение между нижнепермскими Eryopoidea и триасовыми капитозаврами.

Циклотозавр (Cyclotosaurus) — вымершее позднетриасовое земноводное из группы темноспондилов. Относится к отряду капитозавров, семейству Cyclotosauridae.

Интазух (лат. Intasuchus silvicola) — вымерший вид темноспондилов. Единственный представитель в семействе Intasuchidae (интазухиды) из надсемейства Archegosauroidea. Родич архегозавров, платиопозавров и мелозавров. Типовой и единственный вид рода Intasuchus, описанный Е. Д. Конжуковой в 1956 году из пермских отложений Инты. Название дано в честь места обнаружения.
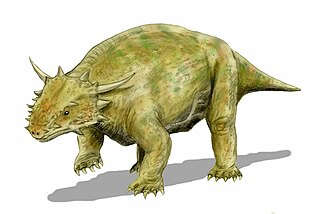
Элгиния — вид парарептилий-анапсид из семейства Elginiidae клады парейазавров, живших во времена пермского периода на территории современной Шотландии (Великобритания).

Бафетиды (лат. Baphetidae) — семейство примитивных стегоцефалов, живших в каменноугольном периоде. Одни из первых ископаемых «земноводных», ставших известными науке — впервые описаны У. Даусоном в 1850 году. В то же время, это одни из самых древних последевонских стегоцефалов. Известны практически только по ископаемым черепам. Черепа обычно низкие, по структуре костей близки к примитивным темноспондилам. Желобки боковой линии обычно присутствуют. Орбита расширена вниз и вперёд, образуя подобие «замочной скважины», расширение превосходит орбиту по размерам. Есть довольно значительно развитая «ушная» вырезка. Нёбо закрытое, как у антракозавров, строение таблитчатых костей сходно с таковым у темноспондилов. Есть шов между щекой и крышей черепа, слуховая капсула доходит до таблитчатой кости. Зубы лабиринтодонтные. Развиты крупные клыки на нёбе, краевые зубы также крупные, двухрядные на нижней челюсти. Нижняя челюсть высокая, выше чем верхняя челюсть. По-видимому, рыбоядные хищники, постоянноводные. Назначение расширения глазницы неясно. Высказываются следующие предположения: в «окне» могло быть крепление мышц, какие-либо железы, электрические органы.

Диссорофиды — семейство темноспондилов позднего карбона — перми. Наиболее наземные из всех темноспондилов. Скелет массивный, тело короткое, конечности длинные, сильные. Голова огромная относительно размеров тела. Череп высокий и широкий, огромные глазницы, крупное теменное отверстие. Огромные ушные вырезки, судя по строению окружающей кости, содержали барабанную перепонку. Затылочный мыщелок парный. Челюстное сочленение на уровне затылочного. Скульптура черепа гребнистая. Края орбит и теменного отверстия окружены костными валиками, что предполагает толстую кожу. У большинства известен дермальный панцирь на спине из парных рядов костных пластин, приросших к остистым отросткам позвонков, у какопса пластины лежали в два слоя. Кожа, по-видимому, мягкая, следов чешуи не выявлено. Хвост короткий. Вероятно, прибрежные амфибиотические и полуназемные хищники. Есть свидетельства, что крупные диссорофиды могли питаться и падалью.

Гефиростегиды (лат. Gephyrostegidae) — семейство вымерших рептилиоморф из монотипического отряда или подотряда гефиростегов (Gephyrostegida), живших во времена каменноугольного периода на территории современных Германии, Чехии и США. В отличие от других рептилиоморф, лишены зубов на поперечных отростках птеригоидов.

Шонизавры — род вымерших пресмыкающихся из семейства Shastasauridae, самые крупные ихтиозавры, известные науке не по фрагментам скелетов. Жили во времена среднетриасовой эпохи.

Лэдлерия — раннетриасовый темноспондил. Животное, из группы трематозавров. Череп очень плоский, в виде равнобедренного треугольника, с острой мордой. Зубы нижней челюсти крупнее, чем на верхней, есть нижнечелюстные «клыки». Верхнечелюстная кость отделена от хоаны шовным соединением между сошником и нёбом. Более 8 нёбных зубов. Орбиты некрупные, вблизи середины черепа, сближены, направлены вверх. Есть боковой вырост квадратоскуловой кости. Стремя массивное, барабанной перепонки не было. Орнамент черепа в виде ямок и гребней. Сохраняются желобки «боковой линии» на передне-верхней поверхности черепа. Тело уплощённое, массивное, покрытое мелкими многоугольными костными пластинками. Конечности плохо известны, хвост, вероятно, короткий. Длина черепа до 10 см, общая длина 30—40 см. Происходит из раннего триаса Южной Африки. Единственный вид — L. gracilis, описан Дж. Китчингом в 1958 году. Близкий род Uruyiella с единственным видом U. liminea описан в 2007 году из низов триаса Уругвая на основании фрагментов черепа. Образ жизни лэдлерий малопонятен. Иногда их считают наземными животными, но против этого свидетельствует наличие органов «боковой линии». Вероятно, всё же эти родичи плагиозавров были в основном водными малоподвижными охотниками за некрупной добычей.
Затрахидиды или затрахеиды — семейство темноспондилов из ранней перми.

Эрициолацерта (Ericiolacerta) — мелкий тероцефал раннетриасовой эпохи, принадлежащий к бауриаморфам. Описана Д. Уотсоном в 1931 году. Единственный вид — E. parva из зоны Lystrosaurus раннего триаса Южной Африки. Мелкое животное, общая длина до 20—25 см. Череп грушевидный, удлинённый, уплощённый. Заглазничная дуга неполная. Теменного отверстия нет. Хорошо развито вторичное нёбо, образованное сошником и нёбными костями. Квадратные ветви крыловидных костей очень тонкие. Предчелюстная и челюстная кость покрыты множеством отверстий. Нижняя челюсть длинная и тонкая, с зачаточным венечным отростком. Симфиз нижней челюсти длинный и скошенный. Зубы мелкие, цилиндрические, клыки не выделяются, первая пара нижнечелюстных зубов направлена косо вперёд («резцы»). Заклыковые зубы со слабо трёхраздельными тупыми коронками. Скелет довольно компактный, выражен поясничный отдел позвоночника, три крестцовых позвонка. Хвост короткий. Лопатка длинная и тонкая, верхний конец её приближен к черепу. Таз низкий, короткий, широкий, с тироидными отверстиями. Кости конечностей длинные, тонкие. Стопа пальцеходящая. В целом по очертаниям скелета животное несколько напоминало ежа. По-видимому, этот высокоразвитый тероцефал был покрыт шерстью, имел развитые вибриссы на морде и внешне походил на млекопитающее. Питалась эрициолацерта насекомыми и другими беспозвоночными, занимая экологическую нишу современных насекомоядных. Остатки этого вида известны и из одновозрастной формации Фремоу в Антарктиде.

Аскептозавр — род рептилий из отряда талаттозавров. Череп низкий, с удлиненной заглазничной частью. Глаза крупные, защищены склеротическими кольцами. Предплечье и голень сильно укорочены. Обитал в море, выползая на сушу только для того, чтобы отложить яйца. У аскептозавра было стройное тело с длинной шеей и вытянутыми зубастыми челюстями. Вероятно, он плавал, волнообразно извиваясь в воде, как змея или угорь, и помогая себе перепончатыми лапами. Кормом служила рыба, за которой ящер, судя по всему, мог довольно глубоко нырять.
- Время жизни — средний триас, около 220 млн лет назад
- Место находки — Европа
- Длина — 2 м
- Вес — 45 кг

Диадектоморфы (лат. Diadectomorpha) — отряд крупных рептилиоморф из карбона и нижней перми Еврамерики и верхней перми Азии, очень близкий к родоначальникам амниот. Включают крупные плотоядные и еще более крупные травоядные формы, некоторые — полуводные, другие — полностью наземные. Диадектоморфы, видимо, возникли в верхнем миссисипии, но широкое распространение получили после коллапса каменноугольных лесов и процветали во времена верхнего пенсильвания и нижней перми. Несмотря на широкое распространение и видовое разнообразие диадектоморфы не были доминантами. Часто их описывают по фрагментарным останкам. В Еврамерике полностью вымирают на границе нижней и средней перми, из средней перми Татарстана известен один образец, который, возможно, относится к диадектам. Позднейший из найденных образцов Alveusdectes fenestratus происходит из верхней перми Цзиюань (Китай).

Скелет акилозавров, как и у всех позвоночных животных, являлся частью опорно-двигательной системы, и был устроен по общему принципу четвероногих динозавров. Тем не менее анкилозавры представляли собой достаточно своеобразную группу, характеризующуюся обширными продольными рядами дермальных окостенений на верхней и боковой поверхностях шеи, туловища и хвоста, в результате чего в скелете наблюдается приспособление к ношению и поддержке остеодермального панциря. В дополнение к этому анкилозавры обладают необычной черепной архитектурой, которая включает костный «орнамент».