Пектинэстераза
| Пектинэстераза | |
|---|---|
![Пектиновая метилэстераза из Dickeya dadantii в комплексе с гексасахаридом. PDB 2ntb [1]](https://upload.wikimedia.org/wikipedia/commons/thumb/5/50/2ntb.png/274px-2ntb.png) Пектиновая метилэстераза из Dickeya dadantii в комплексе с гексасахаридом. PDB 2ntb [1] | |
| Идентификаторы | |
| Шифр КФ | 3.1.1.11 |
| Номер CAS | 9025-98-3 |
| Базы ферментов | |
| IntEnz | IntEnz view |
| BRENDA | BRENDA entry |
| ExPASy | NiceZyme view |
| MetaCyc | metabolic pathway |
| KEGG | KEGG entry |
| PRIAM | profile |
| PDB structures | RCSB PDB PDBe PDBj PDBsum |
| Gene Ontology | AmiGO • EGO |
| Поиск | |
| PMC | статьи |
| PubMed | статьи |
| NCBI | NCBI proteins |
| CAS | 9025-98-3 |
Пектинэстераза (PE) (Шифр КФ 3.1.1.11) — это широко распространённый фермент, связанный с клеточной стенкой, который представляет несколько изоформ, которые способствуют модификации и последующему разрушению клеточной стенки растений. Он содержится во всех высших растениях, а также в некоторых бактериях и грибах. Пектинэстераза действует, прежде всего, за счёт изменения локального pH клеточной стенки, что приводит к нарушениям целостности клеточной стенки.
Pectinesterase катализирует де-этерификацию из пектина в пектат и метанол. Пектин — один из основных компонентов клеточной стенки растений. У растений пектинэстераза играет важную роль в метаболизме клеточной стенки во время созревания плодов. У патогенных бактерий растений, таких как Erwinia carotovora, и у грибковых патогенов, таких как Aspergillus niger, пектинэстераза участвует в мацерации и мягком гниении растительной ткани. Пектинэстеразы растений регулируются ингибиторами пектинэстеразы, которые неэффективны против микробных ферментов[2].
Функция
Недавние исследования показали, что манипуляции с экспрессией пектинэстеразы могут влиять на многочисленные физиологические процессы. У растений пектинэстераза играет роль в модуляции механической стабильности клеточной стенки во время созревания плодов, удлинении клеточной стенки во время прорастания пыльцы и росте пыльцевых трубок , опадении, удлинении ствола, урожайности клубней и развитии корней. Также было показано, что пектинэстераза играет роль в ответе растений на атаку патогенов. Связанная с клеточной стенкой пектинэстераза Nicotiana tabacum участвует в распознавании рецептора клетки-хозяина для белка движения вируса табачной мозаики, и было показано, что это взаимодействие необходимо для транслокации вируса от клетки к клетке.
Действие пектинэстеразы на компоненты клеточной стенки растений может вызывать два диаметрально противоположных эффекта. Первый — это вклад в укрепление клеточной стенки за счёт образования блоков неэтерифицированных карбоксильных групп, которые могут взаимодействовать с ионами кальция, образуя пектатный гель. Во-вторых, высвобождение протонов может стимулировать активность гидролаз клеточной стенки, способствуя разрыхлению клеточной стенки.
Этерификация пектина
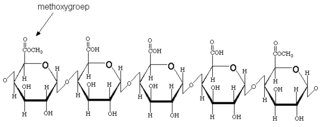
Пектины составляют примерно 35 % от сухого веса клеточных стенок двудольных. Они полимеризуются в цис-цистернах Гольджи, метилэстерифицируются в медиальных цистернах Гольджи и замещаются боковыми цепями в транс-цистернах Гольджи. Биохимия пектина может быть довольно сложной, но, проще говоря, основная цепь пектина состоит из 3 типов полимеров: гомогалактуронана (HGA); рамногалактуронан I (RGI); рамногалактуронан II (RGII).
Гомогалактуронан сильно этерифицируется метилом при экспорте в клеточные стенки и впоследствии деэтерифицируется под действием пектинэстеразы и других пектиновых ферментов. Пектинэстераза катализирует деэтерификацию метилэтерифицированных звеньев D-галактозидуроновой кислоты в пектиновых соединениях, давая субстраты для деполимеризационных ферментов, особенно кислых пектинов и метанола.
Большинство очищенных пектинэстераз растений имеют нейтральные или щелочные изоэлектрические точки и связаны с клеточной стенкой посредством электростатических взаимодействий. Однако пектинэстеразы могут проявлять кислые изоэлектрические точки, обнаруженные в растворимых фракциях тканей растений. До недавнего времени обычно считалось, что растительные пектинэстеразы удаляют метиловые эфиры постепенно, блочно, что даёт длинные непрерывные участки неэтерифицированных остатков GalA в гомогалактуронановых доменах пектина. Альтернативно считалось, что грибковые пектинэстеразы обладают случайной активностью, приводящей к деэтерификации отдельных остатков GalA на взаимодействия фермент/субстрат. В настоящее время показано, что некоторые изоформы пектинэстеразы растений могут проявлять оба механизма, и что такие механизмы управляются изменениями pH. Оптимальный pH для высших растений обычно находится между pH 7 и pH 8, хотя pH пектинэстеразы грибов и бактерий обычно намного ниже этого значения.
Молекулярная биология и биохимия
Белки PE синтезируются в виде пре-белков из 540—580 аминокислот, обладающих сигнальной последовательностью и большой аминоконцевой протяжённостью около 22 кДа. Это концевое удлинение в конечном итоге удаляют, получая зрелый белок с массой 34-37 кДа. У большинства PE отсутствуют консенсусные последовательности для N-гликозилирования в зрелом белке, хотя по крайней мере один сайт присутствует в области удлинения амино-конца.
Пространственная и временная регуляция активности пектинэстеразы во время развития растений основана на большом семействе изоформ. Недавно систематическое секвенирование генома Arabidopsis thaliana привело к идентификации 66 открытых рамок считывания, которые аннотированы как пектинэстеразы, большинство из которых кодируются как большие пре-пропротеины. Преобласть сигнального пептида необходима для нацеливания фермента на эндоплазматический ретикулум и состоит примерно из 25 аминокислотных остатков. Эти N-концевые области содержат несколько сайтов гликозилирования, и считается, что эти сайты также играют роль в нацеливании.
Считается, что пектинэстераза секретируется в апоплазму с высокометилированным пектином, хотя в какой-то момент на этом секреторном пути N-концевой пропептид отщепляется. В настоящее время роль про-региона неизвестна, хотя было высказано предположение, что он может действовать как внутримолекулярный шаперон, обеспечивая правильную укладку или дезактивирующую активность до завершения вставки PE в клеточную стенку.
В последнее время особое внимание было уделено молекулярным исследованиям пектинэстеразы, которые привели к характеристике нескольких родственных изоформ у различных видов высших растений. Было показано, что некоторые из этих пектинэстераз экспрессируются повсеместно, тогда как другие специфически экспрессируются во время созревания плодов, прорастания пыльцевого зерна или удлинения стебля. Такие данные позволяют предположить, что пектинэстеры кодируются семейством генов, которые по-разному регулируются в зависимости от типа клетки в ответ на специфические сигналы развития или окружающей среды.
Изоформы растений
У двудольных растений идентифицировано несколько изоформ пектинэстеразы, различающихся по молекулярной массе, изоэлектрической точке и биохимической активности. Изоформы пектинэстеразы кодируются семейством генов, некоторые из которых конститутивно экспрессируются во всем растении, тогда как другие по-разному экспрессируются в определённых тканях и на разных стадиях развития. Изоформы пектинэстеразы различаются по различным биохимическим параметрам, таким как относительная молекулярная масса, изоэлектрическая точка, оптимальный pH, сродство к субстрату, потребность в ионах и местоположение.
Структура
| Пектинэстераза, каталитическая | |
|---|---|
| Идентификаторы | |
| Символ | Pectinesterase_cat |
| Pfam | PF01095 |
| PROSITE | PDOC00413 |
| Доступные структуры белков | |
| Pfam | структуры |
| PDB | RCSB PDB; PDBe; PDBj |
| PDBsum | 3D-модель |
N-концевые пропептиды пектинэстеразы различаются по размеру и последовательности и демонстрируют низкий уровень аминокислотной идентичности. С другой стороны, С-концевой каталитический участок является высококонсервативным и представляет собой зрелый фермент. Первая трёхмерная структура, решённая для пектинэстеразы растений, была для изоформы из корня моркови (Daucus carota) и состоит из правой параллельной β-спирали, как это видно во всем семействе углеводных эстераз CE-8, трансмембранного домена и пектин-связывающая щель[3]. Точно так же несколько структур пектинэстеразы были выяснены у грибов и кишечной палочки и разделяют большинство структурных мотивов, наблюдаемых у растений.
Прокариотические и эукариотические пектинэстеразы имеют несколько общих областей сходства последовательностей. Кристаллическая структура пектинэстеразы Erwinia chrysanthemi выявила структуру бета-спирали, аналогичную структуре пектинолитических ферментов, хотя она отличается от большинства структур эстераз[4]. Предполагаемые каталитические остатки находятся в том же месте, что и активный центр и субстрат-связывающая щель пектатлиазы.
Примечания
- ↑ Fries, M.; Ihrig, J.; Brocklehurst, K.; Shevchik, V. E.; Pickersgill, R. W. (2007). "Molecular basis of the activity of the phytopathogen pectin methylesterase". The EMBO Journal. 26 (17): 3879—3887. doi:10.1038/sj.emboj.7601816. PMC 2000356. PMID 17717531.
- ↑ "Structural basis for the interaction between pectin methylesterase and a specific inhibitor protein". Plant Cell. 17 (3): 849—858. 2005. doi:10.1105/tpc.104.028886. PMID 15722470.
- ↑ PDB 1GQ8; "Crystal structure of plant pectin methylesterase". FEBS Lett. 514 (2—3): 243—9. March 2002. doi:10.1016/S0014-5793(02)02372-4. PMID 11943159.
- ↑ PDB 1QJV; "Three-dimensional structure of Erwinia chrysanthemi pectin methylesterase reveals a novel esterase active site". J. Mol. Biol. 305 (4): 951—960. 2001. doi:10.1006/jmbi.2000.4324. PMID 11162105.
Ссылки
| Активность | |
|---|---|
| Регуляция | |
| Классификация | |
| Типы |
|
| КФ 3.1.1: Гидролазы карбоновых эфиров | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| КФ 3.1.2: Тиоэстеразы[англ.] |
| ||||||||||||||
| КФ 3.1.3: Фосфатазы |
| ||||||||||||||
| КФ 3.1.4: Фосфодиэстеразы |
| ||||||||||||||
| КФ 3.1.6: Сульфатазы |
| ||||||||||||||
| Нуклеазы (включая дезоксирибонуклеазы и рибонуклеазы) |
| ||||||||||||||