Первичная моторная кора
| Первичная моторная кора | |
|---|---|
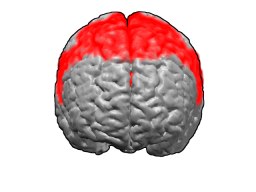
 Первичная моторная кора (поле Бродмана 4) | |
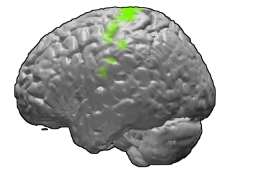
 Первичная моторная кора отмечена зелёным | |
| Часть | Кора больших полушарий |
| Каталоги | |

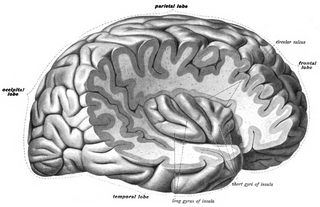
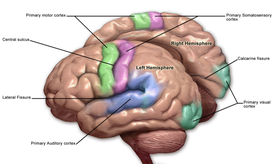
Первичная моторная кора, или первичная двигательная кора (англ. Primary motor cortex) — область моторной коры, расположенная в средне-задней части прецентральной извилины, примыкающей к центральной борозде. Соответствует цитоархитектоническому полю Бродмана 4 и каудальной части поля 6. Содержит клетки Беца и другие мотонейроны, аксоны которых достигают сегментов спинного мозга. Одной из основных функций первичной моторной коры является контроль произвольных дискретных движений, совершаемых группами мышц.
В первичной моторной коре находится топографическое представительство различных областей тела (кортикальный гомункулус) в перевёрнутом виде, от ног (вершина полушария) до рта (нижняя часть полушария). Первичная моторная кора каждого полушария контролирует мышцы на противоположной стороне тела. Площадь поверхности областей первичной моторной коры, управляющих различными частями тела, пропорциональна не физической поверхности этих частей, а числу и степени сложности их моторных реакций. Из-за этого около 3/4 основного объема первичной моторной коры занимает представительство незначительных (по массе), но выполняющих тонкие нестереотипные движения мышц кистей рук, пальцев, мимических мышц лица, губ, языка[1].
Наряду с первичной существует и вторичная моторная кора, к которой относят премоторную кору и дополнительную моторную область.
Структура
Первичная моторная кора расположена вдоль передней стенки центральной борозды, а также простирается вперёд от центральной борозды на прецентральную извилину. Передняя часть первичной моторной коры граничит с латеральной частью премоторной коры. Центральная борозда отделяет первичную моторную кору от первичной соматосенсорной коры, которая расположена вдоль задней стенки центральной борозды. Вентрально первичная моторная кора ограничена островковой долей, дорсально — тянется до верхней части полушария и затем продолжается на медиальной поверхности полушария.
Гистологически область первичной моторной коры отличается от других областей наличием клеток Беца. Слой V первичной моторной коры содержит гигантские (70—100 мкм) пирамидальные нейроны, которые названы клетками Беца. Аксоны этих нейронов идут в контралатеральные моторные ядра черепных нервов и нижние мотонейроны передних рогов серого вещества спинного мозга. Дендриты клеток Беца достигают поверхностного молекулярного слоя, аксоны этих нейронов передают информацию к базальным ганглиям, стволу и спинному мозгу, формируя кортикоспинальный тракт. Стоит отметить, что клетки Беца составляют лишь малую часть кортикоспинального тракта — около 10 % всех нейронов первичной моторной коры[2] и около 2—3% от общего объёма кортикальной проекции для спинного мозга[3]. Несмотря небольшую долю клеток Беца в общем объёме нейронов двигательной коры, они являются чётким маркером, определяющим именно первичную моторную кору[4].
Проводящие пути
Приблизительно 30 % волокон пирамидного тракта берут начало в V слое первичной моторной коры (поле 4), еще 30 % отправляется из премоторной коры (поле 6) и дополнительной моторной области, в которой возникают побудительные сигналы, предшествующие движению, а остальные 40 % — от соматосенсорных областей (теменная доля, поля 3, 2, 1).
Клиническое значение
Поражения прецентральной извилины вызывают паралич определённой группы мышц противоположной стороны тела (лицевой паралич, монопарез руки или ноги, гемипарез).
Кодирование движений
Эвартс[5] предположил, что каждый нейрон в моторной коре вносит вклад в мышечное усилие. Согласно его теории, бо́льшая активность нейронов в моторной коре соответствовала большему мышечному усилию.
Георгопулос и его коллеги[6][7][8] предположили, что «мышечное усилие» является слишком простым описанием функционирования моторной коры. Они изучали активность нейронов в мозге обезьян во время выполнения ими различных движений и обнаружили, что каждый нейрон в моторной коре был максимально активным в ходе специфического направления движения руки, и слабее активировался во время движений в другие, соседние, направления. На основании этого они сделали вывод, что влияние нейронов в моторной коре суммируется, в результате чего рассчитывается определённое направление движения.
Гипотеза о том, что нейроны моторной коры кодируют направление движения, была воспринята в научной среде неоднозначно. Скотт и Каласка[9] показали, что каждый нейрон в моторной коре в большей степени связан с движением сустава или мышечным усилием, нежели чем с направлением движения. Шварц и его коллеги[10] показали, что нейроны моторной моры тесно связаны со скоростью движения руки. Стрик и его коллеги[11] обнаружили, что некоторые нейроны моторной коры активизировались совместно с мышечным усилием, тогда как другие нейроны той же коры активизировались в связи с направлением движения в пространстве. Тодоров[12] предположил, что множество возможных корреляций между нейронами и видами движений руки связаны с мышечным «контролем», при котором параметры движения связаны с мышечным усилием.
В целом параметры, определяющие точный механизм кодирования движений моторной корой, остаются предметом дискуссий и различных гипотез.
Некоторый прогресс в понимании механизмов функционирования моторной коры был получен при изучении мозга грызунов. Так, моторная кора грызунов, как и у приматов, может содержать отдельные области, отвечающие за различные типы двигательных действий[13][14]. К примеру, одна из областей моторной коры отвечает за ритмический контроль вибрисс[13][15][16]. Нейроны в этой области связаны со специфическим подкорковым ядром, в котором центральный генератор упорядоченной активности координирует циклический ритм движения вибрисс. Это ядро посылает моторные команды на мышцы, осуществляющие движение вибрисс.
Дополнительные изображения


 Моторный (двигательный) тракт
Моторный (двигательный) тракт
См. также
Примечания
- ↑ Ерофеев Н.П. Физиология центральной нервной системы. Учебное пособие. — Москва: СпецЛит, 2017. — С. 146. — ISBN 78-5-299-00841-8.
- ↑ Rivara C.B., Sherwood C.C., Bouras C., Hof P.R. Stereologic characterization and spatial distribution patterns of Betz cells in the human primary motor cortex (англ.) // The Anatomical Record Part A: Discoveries in Molecular, Cellular, and Evolutionary Biology[англ.] : journal. — 2003. — Vol. 270, no. 2. — P. 137—151. — doi:10.1002/ar.a.10015. — PMID 12524689.
- ↑ Lassek, A.M. The pyramidal tract of the monkey (англ.) // J. Comp. Neurol.[англ.] : journal. — 1941. — Vol. 74. — P. 193—202. — doi:10.1002/cne.900740202.
- ↑ Ерофеев Н.П. Физиология центральной нервной системы. Учебное пособие. — Москва: СпецЛит, 2017. — С. 145—146. — ISBN 78-5-299-00841-8.
- ↑ Evarts, E.V. Relation of pyramidal tract activity to force exerted during voluntary movement (англ.) // J. Neurophysiol.[англ.] : journal. — 1968. — Vol. 31, no. 1. — P. 14—27. — PMID 4966614.
- ↑ Georgopoulos, A.P., Kalaska, J.F., Caminiti, R. and Massey, J.T. On the relations between the direction of two-dimensional arm movements and cell discharge in primate motor cortex (англ.) // J. Neurosci.[англ.] : journal. — 1982. — Vol. 2, no. 11. — P. 1527—1537. — PMID 7143039.
- ↑ Georgopoulos A.P., Kettner, R.E. and Schwartz, A.B. Primate motor cortex and free arm movements to visual targets in three-dimensional space. II. Coding of the direction of movement by a neuronal population (англ.) // J. Neurosci.[англ.] : journal. — 1988. — Vol. 8, no. 8. — P. 2928—2937. — PMID 3411362.
- ↑ Georgopoulos A.P., Schwartz, A.B. and Kettner, R.E. Neuronal population coding of movement direction (англ.) // Science. — 1986. — Vol. 233, no. 4771. — P. 1416—1419. — doi:10.1126/science.3749885. — PMID 3749885.
- ↑ Scott, S.H.; Kalaska, J.F. Changes in motor cortex activity during reaching movements with similar hand paths but different arm postures (англ.) // J. Neurophysiol.[англ.] : journal. — 1995. — Vol. 73, no. 6. — P. 2563—2567. — PMID 7666162.
- ↑ Moran, D.W.; Schwartz, A.B. Motor cortical representation of speed and direction during reaching (англ.) // J. Neurophysiol.[англ.] : journal. — 1999. — Vol. 82, no. 5. — P. 2676—2692. — PMID 10561437.
- ↑ Kakei, S., Hoffman, D. and Strick, P. Muscle and movement representations in the primary motor cortex (англ.) // Science : journal. — 1999. — Vol. 285, no. 5436. — P. 2136—2139. — doi:10.1126/science.285.5436.2136. — PMID 10497133.
- ↑ Todorov, E. Direct cortical control of muscle activation in voluntary arm movements: a model (англ.) // Nat Neurosci : journal. — 2000. — Vol. 3, no. 4. — P. 391—398. — doi:10.1038/73964. — PMID 10725930.
- ↑ 1 2 Haiss, F.; Schwarz, C. Spatial segregation of different modes of movement control in the whisker representation of rat primary motor cortex (англ.) // J. Neurosci.[англ.] : journal. — 2005. — Vol. 25, no. 6. — P. 1579—1587. — doi:10.1523/JNEUROSCI.3760-04.2005. — PMID 15703412.
- ↑ Ramanathan, D., Conner, J.M. and Tuszynski, M.H. A form of motor cortical plasticity that correlates with recovery of function after brain injury (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2006. — Vol. 103, no. 30. — P. 11370—11375. — doi:10.1073/pnas.0601065103. — PMID 16837575. — PMC 1544093.
- ↑ Brecht, M., Schneider, M., Sakmann, B. and Margrie, T.W. Whisker movements evoked by stimulation of single pyramidal cells in rat motor cortex (англ.) // Nature : journal. — 2004. — Vol. 427, no. 6976. — P. 704—710. — doi:10.1038/nature02266. — PMID 14973477.
- ↑ Cramer, N.P.; Keller, A. Cortical control of a whisking central pattern generator (англ.) // J. Neurophysiol.[англ.] : journal. — 2006. — Vol. 96, no. 1. — P. 209—217. — doi:10.1152/jn.00071.2006. — PMID 16641387. — PMC 1764853.
Ссылки
- Статья Архивная копия от 4 декабря 2020 на Wayback Machine на сайте Brain Info (англ.)
| Лобная доля |
| ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Теменная доля |
| ||||||||||||||
| Затылочная доля |
| ||||||||||||||
| Височная доля |
| ||||||||||||||
| Междолевые борозды |
| ||||||||||||||
| Сводчатая извилина |
| ||||||||||||||
| Островковая доля |
| ||||||||||||||
| Прочее |
| ||||||||||||||