Периферические мембранные белки

1. взаимодействие посредством амфипатической α-спирали, параллельной плоскости мембраны (внутриплоскостная мембранная спираль)
2. взаимодействие посредством гидрофобной петли
3. взаимодействие посредством ковалентно связанного мембранного липида ("липидирование")
4. электростатическое или ионное взаимодействие с мембранными липидами (например, через ион кальция — Ca2+).
Перифери́ческие мембра́нные белки́, или вне́шние мембра́нные белки́ (англ. Peripheral membrane proteins, extrinsic membrane proteins), представляют собой мембранные белки[1], которые обычно непрочно и лишь временно прикрепляются к биологической мембране, с которой они связаны[2]. Данные белки прикрепляются к интегральным мембранным белкам или проникают в периферические области липидного бислоя. Например, регуляторные белковые субъединицы многих ионных каналов и трансмембранных рецепторов можно определить как периферические мембранные белки. В отличие от интегральных мембранных белков, периферические мембранные белки обычно собираются в водорастворимый компонент, или фракцию, всех белков, выделенных в ходе процедуры очистки белков. Белки с GPI-якорями являются исключением из этого правила и могут иметь свойства очистки, аналогичные свойствам интегральных мембранных белков.
Обратимое прикрепление белков к биологическим мембранам регулирует клеточную сигнализацию и многие другие важные клеточные события посредством различных механизмов[3]. Например, тесная связь между многими ферментами и биологическими мембранами может привести их в непосредственную близость к липидному субстрату (субстратам)[4]. Связывание с мембраной может также способствовать перестройке, диссоциации/ассоциации или конформационным изменениям в структурных доменах многих белков, что приводит к увеличению/уменьшению их биологической активности[5][6]. Кроме того, расположение многих белков локализовано либо на внутренней, либо на внешней поверхности или створках их резидентной мембраны[7]. Это облегчает сборку мультибелковых комплексов, увеличивая вероятность любых соответствующих белок-белковых взаимодействий.
Связывание с липидным бислоем

Периферические мембранные белки могут взаимодействовать с другими белками или непосредственно с липидным бислоем. В последнем случае их называют амфитропными белками[5]. Некоторые белки, например G-белки и некоторые протеинкиназы, взаимодействуют с трансмембранными белками и липидным бислоем одновременно. Некоторые полипептидные гормоны, антимикробные пептиды и нейротоксины накапливаются на поверхности мембраны, прежде чем обнаруживают и взаимодействуют со своими рецепторами-мишенями на поверхности клетки, которые сами могут быть периферическими мембранными белками.
Фосфолипидный бислой, образующий поверхностную мембрану клетки, состоит из гидрофобного внутреннего ядра, расположенного между двумя гидрофильными областями, одной на внутренней и одной на внешней поверхности клеточной мембраны. Было показано, что внутренняя и внешняя поверхности, или межфазные области, модельных фосфолипидных бислоев имеют толщину около 8-10 Å (0,8-1 нм) , хотя она может быть больше в биологических мембранах, включающих большое количество ганглиозидов или липополисахаридов[8]. Гидрофобная внутренняя область ядра типичных биологических мембран может иметь толщину около 27-32 Å (2,7-3,2 нм), как показано с помощью малоуглового рентгеновского рассеяния (SAXS)[9]. Пограничная область между гидрофобным внутренним ядром и гидрофильными межфазными областями очень узкая, около 3 Å (0,3 нм). При движении наружу от гидрофобной области ядра к межфазной гидрофильной области эффективная концентрация воды быстро меняется в этом пограничном слое, от почти нуля до концентрации около 2 М[10][11]. Фосфатные группы в фосфолипидных бислоях полностью гидратированы или насыщены водой и расположены примерно на 5 Å (0,5 нм) за границей гидрофобной области ядра[12].
Некоторые водорастворимые белки необратимо связываются с липидными бислоями и могут образовывать трансмембранные альфа-спиральные или бета-баррельные каналы. Такие превращения происходят в порообразующих токсинах, таких как колицин А, альфа-гемолизин и другие. Они также могут происходить в BcL-2-подобном белке, в некоторых амфифильных антимикробных пептидах и в некоторых. Эти белки обычно называют периферическими, поскольку одно из их конформационных состояний — водорастворимое или слабо связанное с мембраной[13].

Механизмы связывания с мембраной
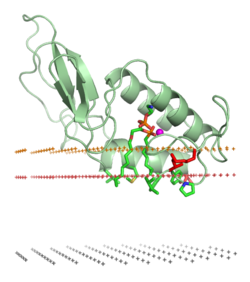
Ассоциация белка с липидным бислоем может сопровождаться значительными изменениями в третичной структуре белка. Они могут включать сворачивание участков структуры белка, которые ранее были развёрнуты, перестройку сворачивания или рефолдинг мембраноассоциированной части белков. Также может происходить образование или диссоциация четвертичных структур белка или олигомерных комплексов, специфическое связывание ионов, лигандов или регуляторных липидов.
Типичные амфитропные белки должны тесно взаимодействовать с липидным бислоем для выполнения своих биологических функций. К ним относятся ферментативная переработка липидов и других гидрофобных веществ, заякоривание мембран, связывание и перенос небольших неполярных соединений между различными клеточными мембранами. Эти белки могут быть прикреплены к бислою в результате гидрофобных взаимодействий между бислоем и открытыми неполярными остатками на поверхности белка[14], путём специфических нековалентных связывающих взаимодействий с регуляторными липидами или путём их присоединения к ковалентно связанным липидным якорям.
Было показано, что сродство к связыванию с мембраной многих периферических белков зависит от специфического липидного состава мембраны, с которой они ассоциированы[15].
Неспецифическая гидрофобная ассоциация
Амфитропные белки связываются с липидным бислоем через различные гидрофобные якорные структуры. Например, амфифильные α-спирали, открытые неполярные петли, посттрансляционно ацилированные или липидированные аминокислотные остатки или ацильные цепи специфически связанных регуляторных липидов, таких как фосфатидилинозитолфосфаты. Было показано, что гидрофобные взаимодействия важны даже для многококатионных пептидов и белков, таких как полиосно́вный домен белка MARCKS или гистактофилин, при наличии их естественных гидрофобных якорей[16].
Ковалентно связанные липидные якоря
Липидные якоря белков ковалентно прикреплены к различным ацильным цепям жирных кислот на цитоплазматической стороне клеточной мембраны посредством пальмитоилирования, миристоилирования или пренилирования. На экзоплазматической (внешней) стороне клеточной мембраны липидные якоря белков ковалентно прикреплены к липидам — гликозилфосфатидилинозитолу (GPI) и холестерину[17][18]. Ассоциация белков с мембранами с помощью ацилированных остатков — обратимый процесс, поскольку ацильная цепь может быть погружена в гидрофобном кармане связывания белка после диссоциации с мембраной. Этот процесс происходит в бета-субъединицах G-белков. Возможно, из-за этой дополнительной потребности в структурной гибкости липидные якоря обычно связываются с высокогибкими сегментами третичной структуры белков, которые плохо поддаются кристаллографическому исследованию.
Специфическое белок-липидное взаимодействие

Некоторые цитозольные белки рекрутируются в различные клеточные мембраны посредством распознавания определённых типов липидов, содержащихся в данной мембране[19]. Связывание белка с определённым липидом происходит через специфические мембранонацеливающие структурные домены, которые находятся внутри белка и имеют специфические карманы для связывания головных групп липидов, с которыми они связываются. Это типичное биохимическое взаимодействие белка с лигандом, которое стабилизируется за счёт образования межмолекулярных водородных связей, ван-дер-ваальсовых взаимодействий и гидрофобных взаимодействий между белком и липидным лигандом. Такие комплексы также стабилизируются за счёт образования ионных мостиков между аспартатными или глутаматными остатками белка и фосфатами липидов через промежуточные ионы кальция (Ca2+). Такие ионные мостики могут возникать и быть стабильными, когда ионы (такие как Ca2+) уже связаны с белком в растворе, до связывания липидов. Образование ионных мостиков наблюдается при белково-липидном взаимодействии между доменами типа C2 белка и аннексинами.
Электростатические взаимодействия сильно зависят от ионной силы раствора. Эти взаимодействия относительно слабы при физиологической ионной силе (0,14 М NaCl): ~ 3-4 ккал/моль для небольших катионных белков, таких как цитохром c, харибдотоксин или гистактофилин[16][20][21].
Пространственное положение в мембране
Ориентация и глубина проникновения многих амфитропных белков и пептидов в мембраны изучаются с помощью сайт-направленной спиновой маркировки[22], химического мечения, измерения сродства к связыванию с мембраной мутантов белков[23], флуоресцентной спектроскопии[24], жидкостной или твёрдотельной ЯМР-спектроскопии[25], спектроскопии ATR FTIR[26], рентгеновской или нейтронной дифракции[27], и вычислительных методов[28][29][30][31].
Были идентифицированы два различных способа мембранной ассоциации белков. Типичные водорастворимые белки не имеют открытых неполярных остатков или каких-либо других гидрофобных якорей. Поэтому они полностью остаются в водном растворе и не проникают в липидный бислой, что было бы энергетически затратно. Такие белки взаимодействуют с бислоями только электростатически, например, таким образом с мембранами взаимодействуют рибонуклеаза и полилизин. Однако типичные амфитропные белки имеют различные гидрофобные якоря, которые проникают в межфазную область и достигают углеводородной внутренней части мембраны. Такие белки «деформируют» липидный бислой, снижая температуру перехода жидкость-гель липида[32]. Связывание обычно представляет собой сильно экзотермическую реакцию[33]. Аналогично происходит ассоциация амфифильных α-спиралей с мембранами[27][34]. Неструктурированные или развёрнутые пептиды с неполярными остатками или липидными якорями также могут проникать через межфазную область мембраны и достигать углеводородного ядра, особенно когда такие пептиды являются катионными и взаимодействуют с отрицательно заряженными мембранами[35][36][37].
Категории белков
Ферменты
Периферические ферменты участвуют в метаболизме различных компонентов мембраны, таких как липиды (фосфолипазы и холестериноксидазы), олигосахариды клеточной стенки (гликозилтрансферазы и трансгликозидазы) или белки (сигнальные пептидазы и тиоэстеразы пальмитоиловых белков). Липазы также могут расщеплять липиды, образующие мицеллы или неполярные капли в воде.
| Класс | Выполняемые функции | Физиология | Структура |
|---|---|---|---|
| Суперсемейство Альфа/бета гидролаз | Катализируют гидролиз химических связей[38]. | Включает бактериальные, грибковые, желудочные и панкреатические липазы, тиоэстеразы пальмитилированных белков, кутиназы и холинэстеразы. | Центральный бета-лист вставлен между двумя слоями альфа-спиралей[39]. |
| Фосфолипаза A2 (секретируемая и цитозольная) | Гидролиз связи sn-2 положения ацильного остатка жирной кислоты в фосфолипиде[40]. | Расщепление липидов, разрушение мембран и липидная сигнализация. | Содержит каталитическую триаду аминокислот: аспартат, серин и гистидин[41]. |
| Фосфолипаза C | Гидролизует PIP2, фосфатидилинозитол, на два вторичных мессенджера (посредника) — инозитолтрифосфат и диацилглицерол[42]. | Липидная сигнализация. | Сердцевинная структура (кор), состоящая из расщепленного цилиндра триозофосфат-изомеразы (ТИМ), который имеет активный участок, каталитические остатки и участок связывания Ca2+[43]. |
| Холестерол оксидазы | Окисляют и изомеризуют холестерин в холест-4-ен-3-он[44]. | Истощают содержание холестерина в клеточные мембранах, который используется в бактериальном патогенезе. | Две петли остатка, которые действуют как крышка на активном сайте[45]. |
| Каротиноидная оксигеназа | Расщепляет каротиноиды[46]. | Каротиноиды используются в растениях и животных как гормоны (в том числе витамин А в организме человека), пигменты, вкусовые и цветочные ароматические вещества, а также защитные соединения. | Состоит из множества ферментов (мультиферментный комплекс), соединённых между собой и образующих ветвистые структуры[47]. |
| Липоксигеназы | Железосодержащие ферменты, катализирующие диоксигенацию полиненасыщенных жирных кислот[48]. | У животных липоксигеназы участвуют в синтезе медиаторов воспаления, известных как лейкотриены. | Сотни аминокислот, составляющих белок, организованы в два домена: N-концевой — бета лист и спиральный C-концевой[49]. |
| Альфа токсины | Расщепляют фосфолипиды в клеточной мембране, подобно фосфолипазе С[50]. | Являются факторами вирулентности Clostridium perfringens. Бактериальный патогенез. | Растворимый мономер с олигомерными «предпоровыми» комплексами[51]. |
| Сфингомиелиназа С | Фосфодиэстераза, расщепляет фосфодиэфирные связи[52]. | Деградация липидов, таких как сфингомиелин. | Домен сапозина и соединительные области с металлофосфатным каталитическим доменом[53]. |
| Гликозилтрансферазы: MurG (эссенциальная бактериальная гликозилтрансфераза, или муреиновая гликозилтрансфераза) и трансгликозидазы. | Катализируют перенос углеводных соединений от активированных донорных молекул к специфическим акцепторным молекулам, образуя гликозидные связи[54]. | Биосинтез дисахаридов, олигосахаридов и полисахаридов (гликоконъюгатов), MurG (эссенциальная бактериальная гликозилтрансфераза, или муреиновая гликозилтрансфераза) участвует в биосинтезе бактериального пептидогликана. | Три петли, богатые глицином: одна в С-конце и две в N-конце[55]. |
| Феррохелатаза | Катализирует процесс превращения протопорфирина IX в гем[56]. | Протопорфирины участвуют в метаболизме порфиринов (гема, хлорофилла итд.) и используются для укрепления яичной скорлупы. | Полипептид, сложенный из двух доменов, каждый из которых имеет четырёхцепочечный параллельный бета-лист, фланкированный альфа-спиралями[57]. |
| Семейство белков, связанных с миотубулярином | Липидфосфатаза, дефосфорилирующая фосфатидилинозитол-3-фосфат и фосфатидилинозитол-3,5-бисфосфат[58]. | Необходим для дифференцировки мышечных клеток. | Содержит GRAM-домен, SET-взаимодействующий домен и PDZ-связывающий домен[59]. |
| Дигидрооротатдегидрогеназы | Окисление дигидрооротата (DHO) до оротата[60]. | Биосинтез пиримидиновых нуклеотидов в прокариотических и эукариотических клетках. | Состоит из двух доменов: альфа/бета бочкообразного домена, содержащего активный сайт, и альфа-спирального домена, образующего туннель для открытия активного сайта[61]. |
| Оксидаза гликолевой кислоты | Катализирует окисление α-гидроксикарбоновых кислот до соответствующих α-кетокислот[62]. | У зелёных растений фермент участвует в фотодыхании. У животных фермент участвует в синтезе оксалата. | β8/α8 фолд, содержащий альфа-спирали, бета-спирали, петли и повороты[63]. |
Мембранно-нацеленные домены («липидные зажимы»)
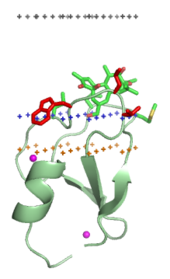
Мембранно-нацеленные домены специфически связываются с головными группами своих липидных лигандов, встроенных в мембрану. Эти липидные лиганды присутствуют в разных концентрациях в различных типах биологических мембран (например, фосфатидилинозитол-3-фосфат можно обнаружить в основном в мембранах ранних эндосом, фосфатидилинозитол-3,5-бисфосфат — в поздних эндосомах, а фосфатидилинозитол-4-фосфат — в мембране комплекса Гольджи)[19]. Таким образом, каждый домен нацелен на определённую мембрану.
- Домены C1 связывают эфиры форбола.
- Домены C2 связывают фосфатидилсерин, фосфатидилхолин, фосфатидилинозитол-3,4-бисфосфат или фосфатидилинозитол-4,5-бисфосфат.
- Домены плекстриновой гомологии (PH-домены), домены PX и домены Tubby связывают разные фосфоинозитиды.
- Домены FYVE более специфичны для фосфатидилинозитол-3-фосфата.
- Домены ENTH связывают фосфатидилинозитол-3,4-бисфосфат или фосфатидилинозитол-4,5-бисфосфат.
- Домен ANTH связывает фосфатидилинозитол-4,5-бисфосфат.
- Белки семейства ERM (ezrin/radixin/moesin) связывают фосфатидилинозитол-4,5-бисфосфат.
- Другие фосфоинозитид-связывающие белки включают фосфотирозин-связывающий домен и некоторые PDZ-домены. Они связывают фосфатидилинозитол-4,5-бисфосфат.
- Дискоидиновые домены факторов свёртывания крови
- Домены ENTH, VHS и ANTH.
Структурные домены
Структурные домены опосредуют прикрепление других белков к мембранам. Их связывание с мембранами может быть опосредовано ионами кальция (Ca2+), которые образуют мостики между кислыми остатками белка и фосфатными группами липидов, как в аннексинах или GLA-доменах.
| Класс | Выполняемые функции | Физиология | Структура |
|---|---|---|---|
| Аннексины | Кальций-зависимое связывание внутриклеточной мембраны с фосфолипидами[64]. | Функции включают транспортировку везикул, слияние мембран и формирование ионных каналов. | Сердцевина (кор) аннексинов включает в себя 310 аминокислот, который состоит из четырёх аннексиновых повторов, каждый из которых состоит из 5 альфа-спиралей[65]. |
| Синапсин I | Покрывает синаптические везикулы и связывается с несколькими цитоскелетными элементами[66]. | Функции регуляции высвобождения нейромедиаторов. | |
| Синуклеин | Неизвестная клеточная функция[67]. | Предполагается, что он играет роль в регулировании стабильности и/или вращение плазматической мембраны. Связан как с болезнью Паркинсона, так и с болезнью Альцгеймера. | Имеет структуру, схожую с тубулин/актин-связывающими белками. |
| GLA-домены белков системы свёртывания крови | Гамма-карбоксиглутаматные (GLA)-домены отвечают за высокоаффинное связывание ионов кальция[68]. | Участвуют в функционировании факторов свёртывания в каскаде свёртывания крови. | |
| Спектрин и α-актинин-2 | Обнаружен в нескольких цитоскелетных и микрофиламентных белках[69]. | Поддержание целостности плазматической мембраны и структуры цитоскелета. |
Переносчики небольших гидрофобных молекул
Эти периферические белки выполняют функцию переносчиков неполярных соединений между различными типами клеточных мембран или между мембранами и цитозольными белковыми комплексами. К транспортируемым веществам относятся фосфатидилинозитол, токоферолы, ганглиозиды, гликолипиды, производные стеролов, ретинол, жирные кислоты, вода, макромолекулы, эритроциты, фосфолипиды и нуклеотиды.
- Белки-переносчики гликолипидов[70][71]
- Липокалины, включая ретинол-связывающие белки и белки, связывающие жирные кислоты
- Полиизопреноид-связывающий белок, например, белковый домен YceI
- Белки-активаторы ганглиозида GM2
- Домен CRAL-TRIO (белки-переносчики α-токоферола и фосфатидилинозитола sec14p)
- Белки-переносчики стеринов
- Белки переноса фосфатидилинозитола и домены STAR
- Оксистерол-связывающий белок.
Переносчики электронов
Эти белки участвуют в цепях переноса электронов. К ним относятся цитохром c, купредоксины, высокопотенциальный железо-серный белок, адренодоксин-редуктаза, некоторые флавопротеины и другие.
Полипептидные гормоны, токсины и антимикробные пептиды
Многие гормоны, токсины, ингибиторы или антимикробные пептиды специфически взаимодействуют с трансмембранными белковыми комплексами. Они также могут накапливаться на поверхности липидного бислоя до связывания своих белков-мишеней. Такие полипептидные лиганды часто положительно заряжены и электростатически взаимодействуют с анионными мембранами.
Некоторые водорастворимые белки и пептиды также могут образовывать трансмембранные каналы. Обычно они подвергаются олигомеризации, значительным конформационным изменениям и необратимо связываются с мембранами. 3D-структура одного из таких трансмембранных каналов, α-гемолизина, уже определена. В других случаях экспериментальная структура представляет собой водорастворимую конформацию, взаимодействующую с липидным бислоем периферически, хотя некоторые каналообразующие пептиды достаточно гидрофобны и поэтому изучались методом ЯМР-спектроскопии в органических растворителях или в присутствии мицелл.
| Класс | Белки | Физиология |
|---|---|---|
| Токсины и яды |
| Хорошо известные типы биологических токсинов включают нейротоксины, цитотоксины, гемотоксины и некротоксины. Биологические токсины выполняют две основные функции: хищническую (токсины змей, скорпионов и улиток конусов) и защитную (токсины медоносных пчёл и муравьёв)[72]. |
| Токсины морских анемон |
| Ингибирование натриевых и калиевых каналов и образование мембранных пор — основные воздействия, оказываемые более 40 известных пептидных токсинов морских анемон. Морские анемоны — плотоядные животные и используют токсины для охоты и защиты; токсин анемоны по токсичности схож с наиболее токсичными фосфорорганическими боевыми отравляющими веществами[73]. |
| Бактериальные токсины |
| Микробные токсины являются основными факторами вирулентности различных патогенных бактерий. Некоторые токсины образуют поры, которые лизируют клеточные мембраны. Другие токсины ингибируют синтез белка или активируют пути вторичного мессенджера, вызывая резкие изменения в путях передачи сигналов, критически важных для поддержания различных клеточных функций. Некоторые бактериальные токсины могут воздействовать непосредственно на иммунную систему, выступая в роли суперантигенов и вызывая массовую пролиферацию Т-клеток, что приводит к перенапряжению иммунной системы. Ботулотоксин — это сильнейший нейротоксин, который не позволяет нейросекреторным везикулам стыковаться с плазматической мембраной нервного синапса, подавляя высвобождение нейротрансмиттеров[74]. |
| Токсины грибов |
| Эти пептиды характеризуются наличием необычной аминокислоты, α-аминоизомасляной кислоты, и проявляют антибиотические и противогрибковые свойства благодаря своей мембранной каналообразующей активности[75]. |
| Антимикробные пептиды |
| Способы действия антимикробных пептидов, убивающих бактерии, разнообразны и включают в себя разрушение мембран, вмешательство в метаболизм и воздействие на цитоплазматические компоненты. В отличие от многих обычных антибиотиков, эти пептиды, по-видимому, являются бактериоцидными, а не бактериостатическими. |
| Дефензины |
| Дефензины представляют собой разновидность антимикробного пептида; и являются важным компонентом практически всех врождённых защитных механизмов хозяина против микробной инвазии. Дефензины проникают в мембраны микробных клеток посредством электрического притяжения и образуют поры в мембране, обеспечивающие эффлюкс (отток), что в конечном итоге приводит к лизису микроорганизмов[76]. |
| Нейрональные пептиды |
| Данные белки вызывают возбуждение нейронов, вызывают поведенческие реакции, являются мощными вазодилататорами и отвечают за сокращение многих типов гладких мышц[77]. |
| Регуляторы апоптоза | Члены семейства Bcl-2 регулируют проницаемость внешней мембраны митохондрий. Bcl-2 сам по себе подавляет апоптоз различных типов клеток, включая лимфоциты и нейрональные клетки. |
Примечания
- ↑ extrinsic protein | biology | Britannica (англ.). www.britannica.com. Дата обращения: 4 июля 2022. Архивировано 12 декабря 2023 года.
- ↑ Д.Нельсон, М.Кокс. Основы биохимии Ленинджера. — Москва: БИНОМ. Лаборатория знаний, 2011. — Т. 1. — С. 530-531. — 694 с. — ISBN 978-5-94774-365-4.
- ↑ Structure and interactions of C2 domains at membrane surfaces // Protein-Lipid Interactions: From Membrane Domains to Cellular Networks. — Chichester : John Wiley & Sons, 2005. — P. 403–22. — ISBN 3-527-31151-3.
- ↑ Ghosh M, Tucker DE, Burchett SA, Leslie CC (November 2006). "Properties of the Group IV phospholipase A2 family". Progress in Lipid Research. 45 (6): 487—510. doi:10.1016/j.plipres.2006.05.003. PMID 16814865.
- ↑ 1 2 Johnson JE, Cornell RB (2002). "Amphitropic proteins: regulation by reversible membrane interactions (review)". Molecular Membrane Biology. 16 (3): 217—235. doi:10.1080/096876899294544. PMID 10503244.
- ↑ Thuduppathy GR, Craig JW, Kholodenko V, Schon A, Hill RB (June 2006). "Evidence that membrane insertion of the cytosolic domain of Bcl-xL is governed by an electrostatic mechanism". Journal of Molecular Biology. 359 (4): 1045—1058. doi:10.1016/j.jmb.2006.03.052. PMC 1785297. PMID 16650855.
- ↑ Takida S, Wedegaertner PB (June 2004). "Exocytic pathway-independent plasma membrane targeting of heterotrimeric G proteins". FEBS Letters. 567 (2—3): 209—213. doi:10.1016/j.febslet.2004.04.062. PMID 15178324. S2CID 36940600.
- ↑ The energetics of peptide-lipid interactions: modification by interfacial dipoles and cholesterol // Current Topics in Membranes. — Academic Press, 2003. — Vol. 52. — P. 205–253. — ISBN 978-0-12-643871-0.
- ↑ Mitra K, Ubarretxena-Belandia I, Taguchi T, Warren G, Engelman DM (March 2004). "Modulation of the bilayer thickness of exocytic pathway membranes by membrane proteins rather than cholesterol". Proceedings of the National Academy of Sciences of the United States of America. 101 (12): 4083—4088. Bibcode:2004PNAS..101.4083M. doi:10.1073/pnas.0307332101. PMC 384699. PMID 15016920.
- ↑ Marsh D (July 2001). "Polarity and permeation profiles in lipid membranes". Proceedings of the National Academy of Sciences of the United States of America. 98 (14): 7777—7782. Bibcode:2001PNAS...98.7777M. doi:10.1073/pnas.131023798. PMC 35418. PMID 11438731.
- ↑ Marsh D (December 2002). "Membrane water-penetration profiles from spin labels". European Biophysics Journal. 31 (7): 559—562. doi:10.1007/s00249-002-0245-z. PMID 12602343. S2CID 36212541.
- ↑ Nagle JF, Tristram-Nagle S (November 2000). "Structure of lipid bilayers". Biochimica et Biophysica Acta (BBA) - Reviews on Biomembranes. 1469 (3): 159—195. doi:10.1016/S0304-4157(00)00016-2. PMC 2747654. PMID 11063882.
- ↑ Goñi FM (2002). "Non-permanent proteins in membranes: when proteins come as visitors (Review)". Molecular Membrane Biology. 19 (4): 237—245. doi:10.1080/0968768021000035078. PMID 12512770. S2CID 20892603.
- ↑ Goforth RL, Chi AK, Greathouse DV, Providence LL, Koeppe RE, Andersen OS (May 2003). "Hydrophobic coupling of lipid bilayer energetics to channel function". The Journal of General Physiology. 121 (5): 477—493. doi:10.1085/jgp.200308797. PMC 2217378. PMID 12719487.
- ↑ McIntosh TJ, Simon SA (2006). "Roles of bilayer material properties in function and distribution of membrane proteins". Annual Review of Biophysics and Biomolecular Structure. 35 (1): 177—198. doi:10.1146/annurev.biophys.35.040405.102022. PMID 16689633.
- ↑ 1 2 Hanakam F, Gerisch G, Lotz S, Alt T, Seelig A (August 1996). "Binding of hisactophilin I and II to lipid membranes is controlled by a pH-dependent myristoyl-histidine switch". Biochemistry. 35 (34): 11036—11044. doi:10.1021/bi960789j. PMID 8780505.
- ↑ Lipidated peptides as tools for understanding the membrane interactions of lipid-modified proteins // Current Topics in Membranes. — Academic Press, 2003. — Vol. 52. — P. 371–395. — ISBN 978-0-12-643871-0.
- ↑ Lipid modifications of proteins // Biochemistry of Lipids, Lipoproteins and Membranes. — 4th. — Elsevier Science, 2002. — P. 37–54. — ISBN 978-0-444-51139-3.
- ↑ 1 2 Cho W, Stahelin RV (June 2005). "Membrane-protein interactions in cell signaling and membrane trafficking". Annual Review of Biophysics and Biomolecular Structure. 34: 119—151. doi:10.1146/annurev.biophys.33.110502.133337. PMID 15869386.
- ↑ Ben-Tal N, Honig B, Miller C, McLaughlin S (October 1997). "Electrostatic binding of proteins to membranes. Theoretical predictions and experimental results with charybdotoxin and phospholipid vesicles". Biophysical Journal. 73 (4): 1717—1727. Bibcode:1997BpJ....73.1717B. doi:10.1016/S0006-3495(97)78203-1. PMC 1181073. PMID 9336168.
- ↑ Protein-lipid interactions with peripheral membrane proteins // Protein-lipid interactions. — Elsevier, 1993. — P. 127–162. — ISBN 0-444-81575-9.
- ↑ Malmberg NJ, Falke JJ (2005). "Use of EPR power saturation to analyze the membrane-docking geometries of peripheral proteins: applications to C2 domains". Annual Review of Biophysics and Biomolecular Structure. 34 (1): 71—90. doi:10.1146/annurev.biophys.34.040204.144534. PMC 3637887. PMID 15869384.
- ↑ Spencer AG, Thuresson E, Otto JC, Song I, Smith T, DeWitt DL, et al. (November 1999). "The membrane binding domains of prostaglandin endoperoxide H synthases 1 and 2. Peptide mapping and mutational analysis". The Journal of Biological Chemistry. 274 (46): 32936—32942. doi:10.1074/jbc.274.46.32936. PMID 10551860.
- ↑ Lathrop B, Gadd M, Biltonen RL, Rule GS (March 2001). "Changes in Ca2+ affinity upon activation of Agkistrodon piscivorus piscivorus phospholipase A2". Biochemistry. 40 (11): 3264—3272. doi:10.1021/bi001901n. PMID 11258945.
- ↑ Kutateladze T, Overduin M (March 2001). "Structural mechanism of endosome docking by the FYVE domain". Science. 291 (5509): 1793—1796. Bibcode:2001Sci...291.1793K. doi:10.1126/science.291.5509.1793. PMID 11230696.
- ↑ Tatulian SA, Qin S, Pande AH, He X (September 2005). "Positioning membrane proteins by novel protein engineering and biophysical approaches". Journal of Molecular Biology. 351 (5): 939—947. doi:10.1016/j.jmb.2005.06.080. PMID 16055150.
- ↑ 1 2 Hristova K, Wimley WC, Mishra VK, Anantharamiah GM, Segrest JP, White SH (July 1999). "An amphipathic alpha-helix at a membrane interface: a structural study using a novel X-ray diffraction method". Journal of Molecular Biology. 290 (1): 99—117. doi:10.1006/jmbi.1999.2840. PMID 10388560.
- ↑ Murray D, Honig B (January 2002). "Electrostatic control of the membrane targeting of C2 domains". Molecular Cell. 9 (1): 145—154. doi:10.1016/S1097-2765(01)00426-9. PMID 11804593.
- ↑ Efremov RG, Nolde DE, Konshina AG, Syrtcev NP, Arseniev AS (September 2004). "Peptides and proteins in membranes: what can we learn via computer simulations?". Current Medicinal Chemistry. 11 (18): 2421—2442. doi:10.2174/0929867043364496. PMID 15379706.
- ↑ Lomize AL, Pogozheva ID, Lomize MA, Mosberg HI (June 2006). "Positioning of proteins in membranes: a computational approach". Protein Science. 15 (6): 1318—1333. doi:10.1110/ps.062126106. PMC 2242528. PMID 16731967.
- ↑ Comparison with experimental data. Orientations of Proteins in Membranes. University of Michigan. Дата обращения: 8 февраля 2007. Архивировано 25 февраля 2007 года.
- ↑ Papahadjopoulos D, Moscarello M, Eylar EH, Isac T (September 1975). "Effects of proteins on thermotropic phase transitions of phospholipid membranes". Biochimica et Biophysica Acta (BBA) - Biomembranes. 401 (3): 317—335. doi:10.1016/0005-2736(75)90233-3. PMID 52374.
- ↑ Seelig J (November 2004). "Thermodynamics of lipid-peptide interactions". Biochimica et Biophysica Acta (BBA) - Biomembranes. 1666 (1—2): 40—50. doi:10.1016/j.bbamem.2004.08.004. PMID 15519307.
- ↑ Darkes MJ, Davies SM, Bradshaw JP (1997). "Interaction of tachykinins with phospholipid membranes: A neutron diffraction study". Physica B. 241: 1144—1147. Bibcode:1997PhyB..241.1144D. doi:10.1016/S0921-4526(97)00811-9.
- ↑ Ellena JF, Moulthrop J, Wu J, Rauch M, Jaysinghne S, Castle JD, Cafiso DS (November 2004). "Membrane position of a basic aromatic peptide that sequesters phosphatidylinositol 4,5 bisphosphate determined by site-directed spin labeling and high-resolution NMR". Biophysical Journal. 87 (5): 3221—3233. Bibcode:2004BpJ....87.3221E. doi:10.1529/biophysj.104.046748. PMC 1304792. PMID 15315949.
- ↑ Marcotte I, Dufourc EJ, Ouellet M, Auger M (July 2003). "Interaction of the neuropeptide met-enkephalin with zwitterionic and negatively charged bicelles as viewed by 31P and 2H solid-state NMR". Biophysical Journal. 85 (1): 328—339. Bibcode:2003BpJ....85..328M. doi:10.1016/S0006-3495(03)74477-4. PMC 1303088. PMID 12829487.
- ↑ Zhang W, Crocker E, McLaughlin S, Smith SO (June 2003). "Binding of peptides with basic and aromatic residues to bilayer membranes: phenylalanine in the myristoylated alanine-rich C kinase substrate effector domain penetrates into the hydrophobic core of the bilayer". The Journal of Biological Chemistry. 278 (24): 21459—21466. doi:10.1074/jbc.M301652200. PMID 12670959.
- ↑ Pfam entry Abhydrolase 1. Дата обращения: 25 января 2007. Архивировано из оригинала 29 сентября 2007 года.
- ↑ Bauer, Tabea L.; Buchholz, Patrick C. F.; Pleiss, Jürgen (March 2020). "The modular structure of α/β‐hydrolases". The FEBS Journal (англ.). 287 (5): 1035—1053. doi:10.1111/febs.15071. ISSN 1742-464X. Архивировано 13 декабря 2023. Дата обращения: 13 декабря 2023.
- ↑ Pfam entry: Phospholipase A2. Дата обращения: 25 января 2007. Архивировано из оригинала 29 сентября 2007 года.
- ↑ Casale, Jarett; Kacimi, Salah Eddine O.; Varacallo, Matthew (2023), "Biochemistry, Phospholipase A2", StatPearls, Treasure Island (FL): StatPearls Publishing, PMID 30521272, Архивировано 18 июня 2022, Дата обращения: 29 ноября 2023
- ↑ Pfam entry: Phosphatidylinositol-specific phospholipase C, X domain. Дата обращения: 25 января 2007. Архивировано из оригинала 29 сентября 2007 года.
- ↑ "Phospholipase C", Wikipedia (англ.), 2023-08-16, Дата обращения: 29 ноября 2023
- ↑ Pfam entry: Cholesterol oxidase. Дата обращения: 25 января 2007. Архивировано из оригинала 29 сентября 2007 года.
- ↑ Yue, Q. Kimberley; Kass, Ignatius J.; Sampson, Nicole S.; Vrielink, Alice (1999-04-01). "Crystal Structure Determination of Cholesterol Oxidase from Streptomyces and Structural Characterization of Key Active Site Mutants ,". Biochemistry (англ.). 38 (14): 4277—4286. doi:10.1021/bi982497j. ISSN 0006-2960. Архивировано 13 декабря 2023. Дата обращения: 13 декабря 2023.
- ↑ Pfam entry: Retinal pigment epithelial membrane protein. Дата обращения: 25 января 2007. Архивировано из оригинала 29 сентября 2007 года.
- ↑ "Carotenoid oxygenase", Wikipedia (англ.), 2023-11-29, Дата обращения: 29 ноября 2023
- ↑ Pfam entry: Lipoxygenase. Дата обращения: 25 января 2007. Архивировано из оригинала 29 сентября 2007 года.
- ↑ Prigge, S. T.; Boyington, J. C.; Faig, M.; Doctor, K. S.; Gaffney, B. J.; Amzel, L. M. (1997-11-01). "Structure and mechanism of lipoxygenases". Biochimie. 79 (11): 629—636. doi:10.1016/S0300-9084(97)83495-5. ISSN 0300-9084. Архивировано 13 декабря 2023. Дата обращения: 13 декабря 2023.
- ↑ PDBsum entry: Alpha Toxin. Дата обращения: 13 декабря 2023. Архивировано 18 мая 2023 года.
- ↑ Alpha Toxin - an overview | ScienceDirect Topics. www.sciencedirect.com. Дата обращения: 29 ноября 2023.
- ↑ Pfam entry: Type I phosphodiesterase. Дата обращения: 25 января 2007. Архивировано из оригинала 29 сентября 2007 года.
- ↑ Xiong, Zi-Jian; Huang, Jingjing; Poda, Gennady; Pomès, Régis; Privé, Gilbert G. (2016-07-31). "Structure of Human Acid Sphingomyelinase Reveals the Role of the Saposin Domain in Activating Substrate Hydrolysis". Journal of Molecular Biology. 428 (15): 3026—3042. doi:10.1016/j.jmb.2016.06.012. ISSN 0022-2836.
- ↑ Pfam entry: Glycosyl transferases group 1. Дата обращения: 25 января 2007. Архивировано из оригинала 29 сентября 2007 года.
- ↑ Ünligil, Uluğ M; Rini, James M (2000-10-01). "Glycosyltransferase structure and mechanism". Current Opinion in Structural Biology. 10 (5): 510—517. doi:10.1016/S0959-440X(00)00124-X. ISSN 0959-440X.
- ↑ Pfam entry: Ferrochelatase. Дата обращения: 25 января 2007. Архивировано из оригинала 29 сентября 2007 года.
- ↑ Al-Karadaghi, Salam; Hansson, Mats; Nikonov, Stanislav; Jönsson, Bodil; Hederstedt, Lars (November 1997). "Crystal structure of ferrochelatase: the terminal enzyme in heme biosynthesis". Structure. 5 (11): 1501—1510. doi:10.1016/s0969-2126(97)00299-2. ISSN 0969-2126.
- ↑ Pfam entry:Myotubularin-related. Дата обращения: 25 января 2007. Архивировано из оригинала 26 сентября 2007 года.
- ↑ Emery and Rimoin's Principles and Practice of Medical Genetics (англ.). ScienceDirect. Дата обращения: 29 ноября 2023. Архивировано 13 декабря 2023 года.
- ↑ Pfam entry:Dihydroorotate dehydrogenase. Дата обращения: 25 января 2007. Архивировано из оригинала 26 сентября 2007 года.
- ↑ Liu, Shenping; Neidhardt, Edie A; Grossman, Trudy H; Ocain, Tim; Clardy, Jon (January 2000). "Structures of human dihydroorotate dehydrogenase in complex with antiproliferative agents". Structure. 8 (1): 25—33. doi:10.1016/s0969-2126(00)00077-0. ISSN 0969-2126.
- ↑ Pfam entry: FMN-dependent dehydrogenase. Дата обращения: 25 января 2007. Архивировано из оригинала 29 сентября 2007 года.
- ↑ Glycolate oxidase - Proteopedia, life in 3D (англ.). proteopedia.org. Дата обращения: 28 ноября 2023. Архивировано 13 декабря 2023 года.
- ↑ Pfam entry: Annexin. Дата обращения: 25 января 2007. Архивировано из оригинала 29 сентября 2007 года.
- ↑ Gerke V, Creutz CE, Moss SE (June 2005). "Annexins: linking Ca2+ signalling to membrane dynamics". Nat. Rev. Mol. Cell Biol. 6 (6): 449—61. doi:10.1038/nrm1661. PMID 15928709. S2CID 37526262.
- ↑ Pfam entry Synapsin N. Дата обращения: 25 января 2007. Архивировано из оригинала 26 сентября 2007 года.
- ↑ Pfam entry Synuclein. Дата обращения: 25 января 2007. Архивировано из оригинала 26 сентября 2007 года.
- ↑ Pfam entry: Gla. Дата обращения: 25 января 2007. Архивировано из оригинала 29 сентября 2007 года.
- ↑ Pfam entry Spectrin. Дата обращения: 25 января 2007. Архивировано из оригинала 26 сентября 2007 года.
- ↑ Rao CS, Lin X, Pike HM, Molotkovsky JG, Brown RE (November 2004). "Glycolipid transfer protein mediated transfer of glycosphingolipids between membranes: a model for action based on kinetic and thermodynamic analyses". Biochemistry. 43 (43): 13805—15. doi:10.1021/bi0492197. PMC 2596630. PMID 15504043.
- ↑ Airenne TT, Kidron H, Nymalm Y, Nylund M, West G, Mattjus P, Salminen TA (January 2006). "Structural evidence for adaptive ligand binding of glycolipid transfer protein". Journal of Molecular Biology. 355 (2): 224—36. doi:10.1016/j.jmb.2005.10.031. PMID 16309699.
- ↑ Animal toxins: facts and protocols. — Basel : Birkhũser Verlag, 2000. — ISBN 3-7643-6020-8.
- ↑ Sea Anemone Toxins. The ASA Newsletter (1999). Архивировано 15 июня 2013 года.
- ↑ Schmitt CK, Meysick KC, O'Brien AD (1999). "Bacterial toxins: friends or foes?". Emerging Infectious Diseases. 5 (2): 224—234. doi:10.3201/eid0502.990206. PMC 2640701. PMID 10221874.
- ↑ Chugh JK, Wallace BA (August 2001). "Peptaibols: models for ion channels". Biochemical Society Transactions. 29 (Pt 4): 565—570. doi:10.1042/BST0290565. PMID 11498029.
- ↑ Oppenheim JJ, Biragyn A, Kwak LW, Yang D (November 2003). "Roles of antimicrobial peptides such as defensins in innate and adaptive immunity". Annals of the Rheumatic Diseases. 62 (Suppl 2): ii17—ii21. doi:10.1136/ard.62.suppl_2.ii17. PMC 1766745. PMID 14532141.
- ↑ Pfam entry Tachykinin. Дата обращения: 25 января 2007. Архивировано из оригинала 26 сентября 2007 года.
Внешние ссылки
- Peripheral membrane proteins in OPM database
- DOLOP Геномно-ориентированная база данных бактериальных липопротеинов
- Peptaibol database
- Antimicrobial Peptide Database Архивировано 20 июля 2011 года.
Структуры клеточной мембраны | |
|---|---|
| Мембранные липиды[англ.] | |
| Мембранные белки | |
| Другое | |