
Цитохром — небольшой гем-содержащий белок, относится к классу цитохромов, содержит в структуре гем типа c. Выполняет в клетке две функции. С одной стороны, он является одноэлектронным переносчиком, свободно связанным с внутренней мембраной митохондрий, и необходимым компонентом дыхательной цепи. Он способен окисляться и восстанавливаться, но не связывает при этом кислород. С другой стороны, при определённых условиях он может отсоединяться от мембраны, переходить в раствор в межмембранном пространстве и активировать апоптоз. Такая двойственность связана со специфичными свойствами молекулы цитохрома c.
Дыхательная цепь переноса электронов, также электрон-транспортная цепь (сокр. ЭТЦ, англ. ETC, Electron transport chain) — система трансмембранных белков и переносчиков электронов, необходимых для поддержания энергетического баланса. ЭТЦ поддерживает баланс за счёт переноса электронов и протонов из НАД∙Н и ФАДН2 в акцептор электронов. В случае аэробного дыхания акцептором может быть молекулярный кислород (О2). В случае анаэробного дыхания акцептором могут быть NO3−, NO2−, Fe3+, фумарат, диметилсульфоксид, сера, SO42−, CO2 и т. д. ЭТЦ у прокариот локализована в ЦПМ, у эукариот — на внутренней мембране митохондрий. Переносчики электронов расположены в порядке уменьшения сродства к электрону, то есть по своему окислительно-восстановительному потенциалу, где у акцептора самое сильное сродство к электрону. Поэтому транспорт электрона на всём протяжении цепи протекает самопроизвольно с выделением энергии. Выделение энергии в межмембранное пространство при переносе электронов происходит ступенчато, в виде протона (H+). Протоны из межмембранного пространства попадают в протонную помпу, где наводят протонный потенциал. Протонный потенциал преобразуется АТФ-синтазой в энергию химических связей АТФ. Сопряжённая работа ЭТЦ и АТФ-синтазы носит название окислительного фосфорилирования.

Цитохром-c-оксида́за (цитохромоксидаза) или цитохром-c-кислород:оксидоредуктаза, также известная как цитохром aa3 и комплекс IV — терминальная оксидаза аэробной дыхательной цепи переноса электронов, которая катализирует перенос электронов с цитохрома с на кислород с образованием воды. Цитохромоксидаза присутствует во внутренней мембране митохондрий всех эукариот, где её принято называть комплекс IV, а также в клеточной мембране многих аэробных бактерий.

Тилакоиды — ограниченные мембраной компартменты внутри хлоропластов и цианобактерий. В тилакоидах происходят светозависимые реакции фотосинтеза. Слово «тилакоид» происходит от греческого слова θύλακος, означающего «мешочек». Тилакоиды состоят из мембраны, окружающей просвет тилакоида. Тилакоиды хлоропластов часто имеют структуру, напоминающую стопку дисков. Эти стопки называют гранами. Граны соединены межграновыми или строматическими тилакоидами (ламеллами) в единое функциональное пространство.

Хлорофи́лл a — особая форма хлорофилла, используемая для оксигенного фотосинтеза. Сильнее всего поглощает свет в фиолетово-голубой и оранжево-красной части спектра. Этот пигмент жизненно необходим для фотосинтеза в клетках эукариот, цианобактерий и прохлорофитов из-за своей способности отдавать возбуждённые электроны в электрон-транспортную цепь. Хлорофилл a также является частью антенного комплекса и передаёт резонансную энергию, которая затем поступает в реакционный центр, где расположены специальные хлорофиллы P680 и P700.
Электрон-транспортная цепь фотосинтеза — последовательность переносчиков электронов, расположенных на белках фотосинтетических мембран и осуществляющих фотоиндуцированный транспорт электронов, сопряжённый с трансмембранным переносом протонов против электрохимического градиента.
Ферредокси́ны — группа небольших растворимых белков, содержащих железосерные кластеры и являющихся подвижными переносчиками электронов в ряде метаболических процессов. Обычно они переносят один или два электрона за счёт изменения окисленности атомов железа.

Фотосисте́ма I, или пластоциани́н-ферредокси́н-оксидоредукта́за — второй функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он принимает электрон от пластоцианина и, поглощая световую энергию, формирует сильный восстановитель П700, способный через цепь переносчиков электронов осуществить восстановление НАДФ+. Таким образом, при участии ФСI синтезируется источник электронов (НАДФН) для последующих реакций восстановления углерода в хлоропластах в цикле Кальвина. Кроме того, ФСI может осуществлять циклический транспорт электронов, сопряжённый с синтезом АТФ, обеспечивая дополнительный синтез АТФ в хлоропластах.

Цитохром f — самая большая субъединица цитохром-b6/f комплекса шифр КФ 1.10.99.1. По своей структуре и функциям цитохром-b6/f комплекс аналогичен цитохром-b/c1 комплексу митохондрий и пурпурных бактерий. Цитохром f играет роль аналогичную цитохрому c1, несмотря на их различия по вторичной структуре.
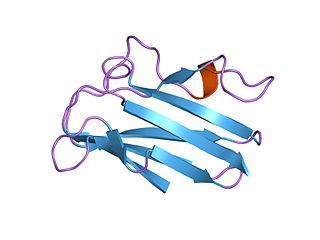
Пластоцианиновое/азуриновое семейство медьсвязывающих белков — семейство маленьких белков способных связывать один атом меди и характеризуемых сильным поглощением с пиком в области 600 нм, благодаря чему в восстановленном состоянии обычно окрашены в ярко-голубой цвет. Самые известные из членов этого класса белков — растительные пластоцианины, маленькие водорастворимые белки хлоропластов, переносящие электрон с цитохрома f, и состоящие в дальнем родстве с бактериальными белками азуринами, которые обмениваются электронами с цитохромом c551. В это же семейство включают белок амицианин из таких бактерий как Methylobacterium extorquens или Paracoccus versutus, способных расти на метиламине; аурацианины A и B из Chloroflexus aurantiacus; голубой медьсодержащий протеин из Alcaligenes faecalis; купредоксин из очистков Cucumis sativus (огурца); кузацианин из огурца; галоцианин, мембраносвязанный белок из Natronomonas pharaonis, псевдоазурин из Pseudomonas; рустицианин из Thiobacillus ferrooxidans; стеллацианин из Rhus vernicifera ; умецианин из корней Armoracia rusticana (хрен); и аллерген Ra3 из амброзии. Этот содержащийся в пыльце белок эволюционно близок перечисленным выше полипептидам, но по-видимому потерял способность связывать медь. Хотя в первичной структуре всех этих белков наблюдается заметное расхождение, участок связывания меди остаётся крайне консервативным.
Терминальная оксидаза, или альтернативная оксидаза пластид, — фермент, который находится на стромальной стороне тилакоидной мембраны в хлоропластах растений и водорослей и внутриклеточных впячиваниях цитоплазматической мембраны цианобактерий. Впервые гипотеза о его существовании была высказан в 1982 году, и подтвердилась годами позже, после открытия в мембране тилакоида фермента, аминокислотная последовательность которого была сходной с таковой у альтернативной оксидазы митохондрий. Обе оксидазы произошли от общего предшественника — белка прокариот, и настолько схожи в структурном и функциональном плане, что помещённая в тилакоид альтернативная оксидаза может заменить выключенную терминальную оксидазу.

Реакционный центр — комплекс белков, пигментов и других кофакторов, взаимодействие которых обеспечивает реакцию превращения энергии света в химическую при фотосинтезе. Реакционный центр получает энергию или через непосредственное возбуждение одной из своих молекул или через перенос энергии от светособирающих комплексов, что даёт начало цепочке химических реакций, происходящей на связанных белками кофакторах. Эти кофакторы — светопоглощающие молекулы такие как хлорофилл, феофитин и хиноны. Энергия фотона используется для поднятия электрона на более высокий энергетический уровень. Запасённая таким образом свободная энергия идёт на восстановление цепочки акцепторов электрона с более высоким редокс-потенциалом.

Фотосисте́ма II (втора́я фотосисте́ма, фотосисте́ма два, ФСII), или H2O-пластохиноноксидоредуктаза — первый функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он расположен в мембранах тилакоидов всех растений, водорослей и цианобактерий. Поглощая энергию света в ходе первичных фотохимических реакций, он формирует сильный окислитель — димер хлорофилла a (П680+), который через цепь окислительно-восстановительных реакций способен вызвать окисление воды.

Цитохро́м-b6f-ко́мплекс, или пластохинолпластоцианинредукта́за — мультибелковый комплекс, который осуществляет окисление пластохинолов и восстановление белка пластоцианина, обеспечивая, таким образом, транспорт электронов между реакционными центрами фотосистемы I (ФСI) и фотосистемы II (ФСII). Он восстанавливает маленький водорастворимый белок пластоцианин, который переносит электрон к ФСII. Аналогичную реакцию катализирует цитохром-bc1-комплекс электрон-транспортной цепи митохондрий. Цитохром-b6f-комплекс присутствует в тилакоидной мембране хлоропластов растений, водорослей и цианобактерий. Он функционально объединяет две фотосистемы в единую цепь переноса электронов от воды к НАДФ+, то есть является участником нециклического потока электронов. Кроме того, цитохромный комплекс вовлечён в циклический транспорт электронов, осуществляемый фотосистемой I.

Цитохро́м-bс1-ко́мплекс или убихинол-цитохром с-оксидоредуктаза, или комплекс III — мультибелковый комплекс дыхательной цепи переноса электронов и важнейший биохимический генератор протонного градиента на мембране митохондрий. Этот мультибелковый трансмембранный комплекс кодируется митохондриальным и ядерным геномами.

Фотофосфорили́рование — процесс синтеза АТФ из АДФ за счёт энергии света. Как и в случае окислительного фосфорилирования, энергия света расходуется на создание протонного градиента на мембране тилакоидов или клеточной мембране бактерии, который затем используется АТФ-синтазой. Фотофосфорилирование — очень древняя форма фотосинтеза, которая есть у всех фототрофных эукариот, бактерий и архей. Различают два типа фосфорилирования — циклическое, сопряжённое с циклическим потоком электронов в электрон-транспортной цепи, и нециклическое, сопряжённое с прямым потоком электронов от H2O к НАДФ+ в случае эукариот или другого донора электрона в случае бактерий, например, H2S. Как разновидность нециклического типа выделяют псевдоциклическое фотофосфорилирование, при котором акцептором электронов служит кислород.

Фотосистема представляет собой функциональную и структурную единицу белковых комплексов, которые осуществляют первичные фотохимические реакции фотосинтеза: поглощение света, преобразование энергии и перенос электронов. Фотосистемы находятся в тилакоидной мембраны растений, водорослей и цианобактерий, либо в цитоплазматической мембране фотосинтезирующих бактерий. В общем случае все фотосистемы подразделяют на два типа — подобные фотосистеме II и подобные фотосистеме I.
П680 (P680, пигмент 680) или первичный донор фотосистемы II — димер из двух молекул хлорофилла а, П1 и П2, которые также называют специальной парой. Вместе две эти молекулы образуют экситонный димер, то есть они функционально представляют собой единую систему и при возбуждении ведут себя как одна молекула. Максимум поглощения энергии света такой специальной пары приходится на длину волны λ = 680 нм. Первичный донор возбуждается, поглощая фотоны с подходящей длиной волны или через перенос энергии возбуждения от других хлорофиллов фотосистемы II. П680 поглощает квант света и переходит в фотовозбуждённое состояние, в результате чего один из его электронов переходит на более высокий энергетический уровень — с основного подуровня S0 на первый синглетный подуровень S1. Этот электрон отрывается от специальной пары и захватывается первичным акцептором электрона, феофитином, который расположен внутри фотосистемы II рядом с П680. Процесс отщепления электрона от специальной пары и переходе его на феофитин с образованием радикальной пары носит название разделения зарядов. Окисленный П680+ восстанавливается, захватывая электрон от водоокисляющего комплекса фотосистемы II.
Цитохром b559 — гетеродимерный белок, состоящий из одной альфа (PsbE) и одной бета (PsbF) субъединицы, между которыми расположен гем; важный компонент фотосистемы II.
НАДН-дегидрогена́зный ко́мплекс хлоропла́стов — мультибелковый комплекс электронотранспортной цепи фотосинтеза, расположенный в тилакоидной мембране пластид высших растений и водорослей. Комплекс окисляет ферредоксин и восстанавливает молекулы пластохинона, которые высвобождаются в мембрану. При этом энергия окисленного восстановительного эквивалента расходуется на перенос протонов из стромы хлоропласта в люмен тилакоида с образованием протонного градиента. Показано большее сходство НАДН-дегидрогеназного комплекса хлоропластов с цианобактериальным НАДН дегидрогеназным комплексом (NDH-1), чем с митохондриальным комплексом I.

