Позвоночные
| Позвоночные | |
|---|---|
 Сверху: американский атлантический осётр (костные рыбы), саванный слон (четвероногие); Снизу: тигровая акула (хрящевые рыбы), речная минога (бесчелюстные). | |
| Научная классификация | |
Домен: Эукариоты Царство: Животные Подцарство: Эуметазои Без ранга: Двусторонне-симметричные Без ранга: Вторичноротые Тип: Хордовые Подтип: Позвоночные | |
| Международное научное название | |
| Vertebrata Lamarck, 1801[1] | |
| Синонимы | |
| |
| Инфратипы | |
| |
Позвоно́чные (лат. Vertebrata) — подтип хордовых животных. Доминирующая (наряду с насекомыми) на земле и в воздушной среде группа животных. Обилие и разнообразие позвоночных делают их важным экологическим фактором.
Роль позвоночных в современном животном мире настолько велика, что всех остальных животных объединяют в группу (несистематическую) беспозвоночные, несмотря на то что они могут иметь очень мало схожестей друг с другом - например, иглокожие относятся к вторичноротым, а членистоногие и моллюски - к первичноротым.
Позвоночные — высший подтип хордовых. По сравнению с низшими хордовыми — бесчерепными и оболочниками — они характеризуются значительно более высоким уровнем организации, что наглядно выражено как в их строении, так и в физиологии. Если низшие хордовые ведут сидячий (прикреплённый) или малоподвижный образ жизни, то поведение позвоночных несравненно более сложно. Среди них нет видов с сидячим образом жизни; они перемещаются в широких пределах, активно разыскивая и захватывая пищу, спасаясь от преследования врагов, разыскивая для размножения особей другого пола[2]. Активному образу жизни представителей подтипа отвечает высокий уровень развития их нервной системы[3].
Строение
Особенности строения
Активные перемещения обеспечивают позвоночным животным возможность смены мест обитания в зависимости от изменений условий существования и потребностей на разных этапах их жизненного цикла, например при развитии, половом созревании, размножении, зимовках и т. д. Указанные общебиологические черты позвоночных прямо связаны с особенностями их морфологической организации и с физиологией.
Нервная система значительно более дифференцирована, чем у низших хордовых. У всех позвоночных имеется достаточно сложно устроенный головной мозг, развивающийся из трёх пузыревидных вздутий переднего конца нервной трубки и защищённый черепом[3]. Функционирование головного мозга обусловливает высшую нервную деятельность — основу приспособительного поведения. Для позвоночных характерно наличие разнообразных и сложно устроенных органов чувств, служащих основной связью между живым организмом и внешней средой.
С развитием головного мозга и органов чувств связано[3] возникновение черепа, служащего надёжным футляром для этих крайне нежных и важных органов. В качестве осевого скелета взамен хорды у подавляющего большинства животных функционирует более совершенное и прочное образование — позвоночный столб, который выполняет роль не только опорного стержня тела, но и футляра, заключающего в себе спинной мозг. У четвероногих позвоночник, грудина и рёбра образуют грудную клетку - структуру, поддерживающую и защищающую сердце и лёгкие, происходит дифференцировка на отделы, развиваются крестец и таз (у некоторых позвоночных, например у китообразных, таз редуцирован).
В области переднего отдела кишечной трубки возникают подвижные части скелета, из которых формируется ротовой, а у огромного большинства видов — челюстной аппарат, обеспечивающие схватывание и удержание пищи, а у высших позвоночных — и измельчение её[2].
Эволюционная продвинутость позвоночных по сравнению с низшими хордовыми определяется признаками, связанными с тремя типами тканей: вентральным гипомером, плакодами и клетками нервного гребня. Вентральный гипомер образует мускулатуру рта, глотки, сердца и кишечника. Плакоды участвуют в формировании парных сенсорных органов головы и боковой линии. Клетки нервного гребня дают начало меланоцитам, висцеральным дугам, черепу и прочим элементам скелета, а также ганглиям черепных и спинальных нервов[4].
Общий обмен веществ у позвоночных несравненно более высокий, чем у низших хордовых. В этой связи надо указать на характерные черты организации: наличие сердца обусловливает быстрый кровоток; в выделительной системе почки надёжно обеспечивают выведение из организма возросшего количества продуктов обмена.
Позвоночные объединены общностью морфофизиологической организации. Во всех системах органов этих животных можно проследить черты преемственных изменений в связи с эволюционным преобразованием органов. Ниже излагается общий план строения, функционирования и закладки в онтогенезе отдельных систем органов.
Кожные покровы
Кожные покровы — весьма важная в функциональном отношении система. Кожа и мускулы покрывают тело животного, придают ему форму и удерживают внутренние органы. Кожные покровы защищают тело от внешних механических, физических и химических повреждений, воздействия температуры, иссушения, проникновения микробов. Кожа принимает участие в теплорегуляции, газообмене и выведении продуктов распада. Производные кожи могут принимать участие в формировании органов передвижения (копыта), служить для хватания (когти), нападения и защиты (рога, иглы и др.), полёта (складки, крылья), плавания (перепонки). Кожа содержит осязательные рецепторы, в ней много желёз разного назначения (слизистые, жировые, пахучие, потовые и пр.).
Для кожи позвоночных характерна двуслойность (у более примитивного ланцетника, относящегося к бесчерепным хордовым, наружный покров тела ещё однослойный[5]). Её наружный слой — эпидермис — имеет эктодермальное происхождение и всегда многослоен: нижний слой его пожизненно остаётся живым и деятельным, продуцируя новые слои клеток, а верхние слои эпидермиса составлены обычно уплощёнными клетками (которые у наземных позвоночных ороговевают, отмирают и постоянно слущиваются). Эпидермис даёт начало роговым производным — роговым чешуям, перьям, волосам, когтям, копытам. В эпидермисе развиваются разнообразные кожные железы[6].
Внутренний волокнистый слой кожи — кориум — развивается из мезодермального зачатка. Он толст и составляет основную часть кожи, обладающую большой прочностью. В кориуме развиваются разнообразные окостенения в виде чешуи рыб, покровных костей, образующих так называемый кожный скелет (в отличие от хондрального скелета); в нижней части кориума накапливается подкожный слой жира[6].
Мышечная система
Располагающийся под кожей слой мышц составляет основную массу мускулатуры, именуемую мускулатурой тела, или соматической. Она обеспечивает животным возможность движения в среде и состоит из поперечно-полосатой мышечной ткани. У низших позвоночных, как и у бесчерепных, мускулатура сегментирована. У высших позвоночных в связи с общим усложнением телодвижений, с развитием конечностей сегментация нарушается, и туловищная мускулатура группируется, оформляя такие части тела, как торс, голову, органы движения.
Кроме соматической мускулатуры у позвоночных есть мускулатура кишечника и некоторых других внутренних органов (сосудов, каналов). Эта мускулатура носит название висцеральной. Она сложена гладкой мышечной тканью и обеспечивает, в частности, движение пищи в кишечнике, сокращение стенок кровеносных сосудов.
Туловищная мускулатура эмбрионально возникает из внутреннего листка миотома, то есть спинного отдела мезодермы. Висцеральная мускулатура — это производное боковой пластинки, то есть брюшного отдела мезодермы.
Скелет
Внутренний скелет — это опорная основа тела позвоночного животного. Скелет участвует в движении тела, осуществляет защиту внутренних органов. К скелету прикрепляется мускулатура. В костях скелета располагаются кроветворные ткани — в частности, красный костный мозг. Также скелет служит как депо веществ — в нём хранятся запасы кальция и др. веществ.
Топографически скелет позвоночных может быть разделен на осевой, висцеральный, скелет поясов конечностей и свободных конечностей.
Осевой скелет в первоначальном виде представлен хордой, окружённой толстой соединительнотканной оболочкой; последняя охватывает не только хорду, но и лежащую над ней нервную трубку. Хорда развивается из зачатка спинной стороны первичной кишки, то есть имеет энтодермальное происхождение. В осевом скелете различают позвоночный столб и мозговой череп.
У большинства представителей подтипа хорда замещается позвоночником, слагающимся сначала из хрящевых, а потом и костных позвонков. Низшие позвоночные сохраняют хорду всю жизнь, а у высших от неё остаются лишь упругие межпозвоночные диски, придающие позвоночному столбу прочность и упругость[7]. Хрящевой и костный скелеты развиваются как производные вышеупомянутой соединительнотканной (мезодермальной по происхождению) оболочки (таким образом, данная оболочка является скелетогенной).
От каждого позвонка вверх и вниз отходят парные отростки — верхние и нижние дуги. Верхние дуги, смыкаясь концами, образуют канал, в котором располагается спинной мозг; к нижним в туловищной части причленяются рёбра, а в хвостовой части они тоже смыкаются в канал, по которому проходят хвостовые артерии и вена[5].
Конечности бывают двух типов: непарные и парные. Скелет непарных конечностей — спинного, хвостового, анального плавников — состоит из ряда хрящевых или костных лучей, не связанных с другими частями скелета. Скелет парных конечностей подразделяется на скелет поясов конечностей (всегда расположенных внутри тела животного) и на скелет свободной конечности, который бывает двух типов: плавник рыб и (обычно) пятипалая конечность наземных позвоночных (у акантостеги — восьмипалая, у ихтиостеги — семипалая, у птиц обычно четырёхпалая; зачастую редуцируется число пальцев и у копытных млекопитающих)[8]. При этом скелет плавника представлен несколькими рядами хрящиков или косточек, которые перемещаются относительно пояса как единый рычаг, а скелет пятипалой конечности состоит из ряда рычагов (способных перемещаться и совместно относительно пояса конечностей, и отдельно — один относительно другого). Закладка скелета конечностей происходит в соединительнотканном слое кожи.
Органы пищеварения
Система пищеварительных органов представлена трубкой, начинающейся ротовым отверстием и заканчивающейся анальным отверстием. Эпителий пищеварительного тракта является энтодермальным. Только в области ротового и анального отверстий энтодермальный эпителий незаметно переходит в эктодермальный.
Пищеварительный тракт подразделяется на следующие основные отделы:
- ротовая полость, служащая для принятия пищи;
- глотка — отдел, всегда связанный с органами дыхания: у рыб в глотку открываются жаберные щели, у наземных позвоночных в глотке располагается гортанная щель; глотку справедливо называют дыхательным отделом пищеварительной трубки;
- пищевод;
- желудок — расширение кишечного тракта, имеющее в некоторых случаях весьма сложное устройство;
- кишечник, в типичном случае подразделяющийся на переднюю, или тонкую, среднюю, или толстую, и заднюю, или прямую, кишку.
Морфологическое усложнение кишечного тракта в ряду позвоночных идёт по пути его удлинения и дифференцировки на отделы. В пищеварительную трубку открываются протоки трёх видов пищеварительных желёз: слюнных, печени, поджелудочной.
Слюнные железы — приобретение наземных позвоночных. В них преобразуются слизистые железы ротовой полости. Секрет их смачивает пищу и способствует расщеплению углеводов.
Печень и поджелудочная железа развиваются путём выпячивания переднего отдела эмбриональной кишки. Функции обеих желёз шире, чем только пищеварительные. Так, печень кроме выделения желчи, эмульгирующей жиры и активизирующей действие других пищеварительных ферментов, служит важным органом обмена веществ. Здесь нейтрализуются некоторые вредные продукты распада, накапливается гликоген. Ферменты поджелудочной железы расщепляют белки, жиры и углеводы. Одновременно поджелудочная железа служит органом внутренней секреции. Расстройство этой функции приводит к потере способности организма использовать сахар. В результате возникает тяжёлое заболевание — диабет.
Пороками развития пищеварительной системы у человека является сужение и атрезия (отсутствие просвета) пищевода, микро- и макроэзофагус, гастроптоз (опущение желудка), дивертикул Меккеля, обратное расположение внутренних органов, короткая брыжейка. Иногда при незарастании рудиментов жаберных щелей сохраняются врождённые свищи шеи.
Органы дыхания
Органы дыхания позвоночных бывают двух типов — жабры и лёгкие, и у значительной части позвоночных существенное значение в дыхании имеет кожа.
Жаберный аппарат представляет собой систему парных, обычно симметрично расположенных, щелей, служащих для сообщения глотки с наружной средой. Передние и задние стенки жаберных щелей выстланы слизистой оболочкой, образующей пластинчатые выросты; выросты поделены на лепестки, носящие название жаберных. Каждая жаберная пластинка выше лепестков носит название полужабры. В промежутках между жаберными щелями (в жаберных перегородках) располагаются висцеральные жаберные дуги. Таким образом, каждая жаберная дуга связана с двумя полужабрами двух разных жаберных щелей.
Органы дыхания наземных позвоночных — лёгкие — в схеме представляют собой пару мешков, открывающихся в глотку через гортанную щель. Эмбрионально лёгкие возникают в виде выпячивания брюшной стенки глотки в задней части жаберного аппарата, то есть имеют энтодермальное происхождение. На ранних стадиях развития эмбрионов зачатки лёгких напоминают пару внутренних (энтодермальных) жаберных щелей. Эти обстоятельства, а также общие для лёгких и жабр черты кровоснабжения и иннервации заставляют считать лёгкие гомологами задней пары жаберных мешков.
Кожа участвует в дыхании в случаях, когда в ней отсутствуют плотные роговые или костные чешуи, например у земноводных, голокожих рыб.
Функционально дыхательная система участвует в обогащении крови кислородом и в удалении углекислого газа. Через дыхательную систему у низших водных животных происходит сбрасывание аммиака. У теплокровных животных она участвует в процессах терморегуляции. Принцип работы дыхательной системы — обмен CO2 и O2 между потоками газа и крови, направленными противотоком навстречу друг другу.
У человека в процессе нарушения эмбриогенеза возможны сохранение жаберных щелей, атрезия трахеи, трахейно-пищеводные свищи, агенезия (отсутствие) или гипоплазия (недоразвитие) доли или целого лёгкого, недоразвитие бронхов, добавочные доли или целое лёгкое, кисты лёгких.
Органы кровообращения
У всех позвоночных кровеносная система замкнутая — как и у бесчерепных, но (в отличие от бесчерепных) у позвоночных имеется сердце — мускулистый орган, проталкивающий своими сокращениями кровь по сосудам[5]. Кровеносная система позвоночных состоит из сообщающихся между собой кровеносных сосудов, которые (в грубой схеме) могут быть сведены к двум стволам: спинному, где кровь течёт от головы к хвосту, и брюшному, по которому она движется в обратном направлении.
Сердце позвоночных представляет собой толстостенный мускульный мешок, разделённый на несколько отделов — камер. Основными отделами сердца являются предсердие, принимающее кровь, и желудочек, направляющий её по телу. Число камер сердца различно у разных классов позвоночных.
Эмбрионально сердце возникает как расширение задней части брюшной аорты, которая в этом месте свёртывается в изогнутую петлю. Передний отдел петли даёт начало желудочку сердца, задний — предсердию.
Сердце имеет поперечно-полосатую мускулатуру, работающую в автоматическом режиме, и сокращение его не подчинено волевым импульсам. Размеры сердца связаны с интенсивностью его работы, и его размеры относительно размеров тела увеличиваются в ряду позвоночных.
Кровеносные сосуды подразделяются на две системы: артериальную, в которой кровь течёт от сердца, и венозную, по которой кровь возвращается к сердцу. В процессе усложнения позвоночных наблюдается переход от животных, имеющих один круг кровообращения, к обладателям двух кругов кровообращения.
Кровь позвоночных по своей природе относится к соединительной ткани, проникающей в кровеносное русло из межклеточного пространства. Состоит кровь из бесцветной жидкости — плазмы, в которой находятся форменные элементы крови: красные кровяные тельца, или эритроциты, содержащие красящее вещество — гемоглобин, и белые кровяные тельца — лейкоциты. Эритроциты переносят кислород. Лейкоциты участвуют в уничтожении попавших в тело микроорганизмов. Кроме того, в крови есть тромбоциты, играющие важную роль в свёртывании крови, а также прочие клетки. Масса крови в эволюционном ряду позвоночных увеличивается.
Кровеносная система многофункциональна. Она участвует в получении органами, тканями, клетками кислорода, органических и минеральных веществ, жидкостей и выносе продуктов распада, углекислого газа, в переносе гормонов желёз внутренней секреции и т. д.
Механизм транспорта CO2 в крови
В тканях в процессе жизнедеятельности клеток образуется углекислый газ, откуда он попадает в тканевую жидкость и плазму крови. Однако из плазмы крови большая часть CO2 диффундирует в эритроциты. Там часть молекул CO2 реагирует с водой под действием фермента карбоангидразы, давая угольную кислоту H2CO3, которая диссоциирует на ионы H+ и HCO3-. После этого гидрокарбонат-ионы уходят в плазму, и вместо них в эритроцит входят ионы Cl- (этот анионный обмен называется сдвигом Хамбургера[англ.]). Тем временем в эритроцитах оксигемоглобин отдаёт кислород, так как при высоком парциальном давлении CO2 сродство гемоглобина к кислороду снижается (эффект Холдена). Освободившийся гемоглобин связывает часть углекислого газа, диффундировавшего в эритроцит (так образуется карбоксигемоглобин), а другие молекулы гемоглобина связывают два протона, предотвращая закисление крови и появление эффекта Бора (при понижении pH (то есть повышении концентрации CO2) сродство гемоглобина к кислороду падает, что препятствует эффективному захвату кислорода в лёгких). В лёгочных капиллярах перечисленные реакции протекают в обратном направлении. Так как парциальное давление углекислого газа в альвеолах меньше, чем в венозной крови, CO2 уходит в альвеолы из карбоксигемоглобина в эритроцитах; кроме того карбоксигемоглобина, высвобождается также гемоглобин, связавший два протона. При этом парциальное давление углекислого газа в альвеолах растёт, и из-за этого увеличивается сродство гемоглобина к кислороду, который гемоглобин начинает связывать (эффект Бора). В обмен на ионы хлора ионы HCO3- входят в эритроцит, под действием карбоангидразы связываются с высвобожденными гемоглобином протонами, давая угольную кислоту. Последняя распадается на воду и углекислый газ, который также выходит из эритроцитов в альвеолы[9][10].
Лимфатическая система
Наряду с кровеносной системой у позвоночных есть другая, связанная с ней, сосудистая система — лимфатическая. Она состоит из лимфатических сосудов и лимфатических желёз. Лимфатическая система незамкнута. Только крупные её сосуды имеют самостоятельные стенки, в то время как их разветвления открываются в межклеточные пространства различных органов. Лимфатические сосуды содержат бесцветную жидкость — лимфу, в которой плавают лимфоциты, образующиеся в лимфатических железах. Движение лимфы обусловливается сокращением стенок некоторых участков крупных сосудов (так называемых лимфатических сердец) и периодически изменяющимся давлением на сосуды различных органов. Лимфатическая система служит посредницей в обмене веществ между кровью и тканями.
Нервная система
Функции нервной системы — восприятие внешних раздражений и передача возникающих возбуждений к клеткам, органам, тканям, а также объединение и согласование деятельности отдельных систем органов и организма в целом в единую функционирующую живую систему. Нервная система позвоночных в эмбриональный период возникает, так же как и у бесчерепных, в виде закладывающейся в эктодерме на спинной стороне зародыша полой трубки. В последующем происходит её дифференцировка, приводящая к образованию: а) центральной нервной системы, представленной головным и спинным мозгом, б) периферической нервной системы, состоящей из нервов, отходящих от головного и спинного мозга, и в) симпатической нервной системы, состоящей в основе из нервных узлов, расположенных около позвоночного столба и связанных продольными тяжами.
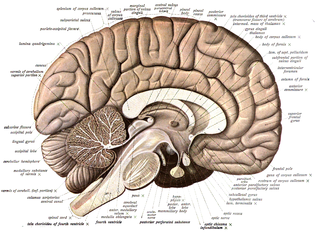
Головной мозг представлен у позвоночных животных пятью отделами: передним, промежуточным, средним, мозжечком и продолговатым мозгом. Он закладывается эмбрионально в виде вздутия переднего отдела нервной трубки, которое вскоре делится на три первичных мозговых пузыря. В дальнейшем передняя часть первого мозгового пузыря даёт начало переднему мозгу; задняя его часть преобразуется в промежуточный мозг. Из второго мозгового пузыря формируется средний мозг. Путём выпячивания крыши третьего мозгового пузыря формируется мозжечок, под которым располагается продолговатый мозг. Передний мозг, кроме того, подразделяется на левую и правую половины.
Одновременно с разрастанием и дифференцировкой головного отдела нервной трубки происходит соответственное преобразование невроцеля. Два его расширения в полушариях переднего мозга известны под названием боковых желудочков мозга. Расширенная часть невроцеля в промежуточном отделе мозга обозначается как третий желудочек, полость среднего мозга — как сильвиев водопровод, полость продолговатого мозга — как четвёртый желудочек, или ромбовидная ямка. От головного мозга отходят 10 или 12 пар черепных нервов.
Передний мозг имеет впереди два симметрично расположенных выступа, от которых отходит первая пара головных нервов — обонятельные. От дна промежуточного мозга отходят зрительные нервы. На крыше промежуточного мозга развиваются два сидящих на ножках выступа: передний — теменной орган и задний — эпифиз.
От дна промежуточного мозга отрастает непарный выступ — воронка, к которой прилегает сложное по строению и функции образование — гипофиз. Передний отдел гипофиза развивается из эпителия ротовой полости, задний — из мозгового вещества. Там же расположен гипоталамус.
Крыша среднего мозга образует парные вздутия — зрительные доли (бугры). От среднего мозга отходит третья пара головных нервов (глазодвигательные). Четвёртая пара головных нервов (блоковые) отходит на границе между средним и продолговатым мозгом, все остальные головные нервы отходят от продолговатого мозга.
Спинной мозг не отграничен резко от продолговатого мозга. В центре спинного мозга (по главной оси органа) сохраняется невроцель, известный у позвоночных под названием спинномозгового канала.
От спинного мозга метамерно (по числу сегментов) отходят спинномозговые нервы. Они начинаются двумя корешками: спинным — чувствующим и брюшным — двигательным. Эти корешки вскоре по выходе из спинного мозга сливаются, образуя спинномозговые нервы, которые затем вновь делятся на спинную и брюшную ветви.
Органы чувств
Эта группа органов возникает как производные разных частей зародыша и на разных этапах его развития. Это органы обоняния, зрения, слуха, вестибулярный аппарат, органы боковой линии, органы вкуса, осязания, специфические органы, воспринимающие магнитное поле Земли, электрические поля, тепловые излучения и пр.
Данные сравнительной эмбриологии позволяют предполагать, что обоняние — одна из самых древних функций мозга. Органы обоняния закладываются в эмбрионе как утолщение эктодермы одновременно с нервной пластинкой. Параллельно формируется скелет обонятельных капсул, которые входят в состав мозгового черепа. Поначалу обонятельные капсулы сообщаются только с внешней средой и имеют наружные ноздри. Впоследствии в связи с наземным существованием ноздри становятся сквозными.
Органы зрения тоже принадлежат к древним органам чувств. У всех позвоночных есть глаза с фокусирующей линзой — хрусталиком, дающим перевёрнутое изображение на сетчатке (у низших позвоночных иногда встречается теменной глаз, затянутый кожей и лишь отличающий свет от темноты)[3]. Фоторецепция возникает на очень раннем этапе эволюции хордовых и формируется в раннем эмбриогенезе.
Органы зрения позвоночных подразделяют на парные и непарные. И те и другие есть выросты промежуточного мозга. Парные глаза закладываются как выросты боковых частей промежуточного мозга, непарные — как последовательно расположенные в крыше промежуточного мозга (эпифиз и теменной орган). Закладка парных глаз сопровождается формированием около них зрительных капсул, входящих в состав мозгового черепа.
Органы слуха имеют у позвоночных животных сложное происхождение. Наиболее рано в эволюции формируется внутреннее ухо, которое закладывается в эктодерме зародыша, углубляется в виде ямки и оформляется как слуховой пузырёк, лежащий в слуховой капсуле. Слуховой пузырёк делится перетяжкой на две части. Верхний отдел превращается в вестибулярный аппарат. Это орган равновесия. Он позволяет ощущать положение тела в трёхмерном пространстве. Этот орган — 3 полукружных канала во внутреннем ухе. Нижний отдел слухового пузырька представляет собой собственно внутреннее ухо — слуховой мешочек. Среднее и наружное ухо формируются на поздних этапах возникновения позвоночных животных в связи с выходом на сушу.
Органы боковой линии свойственны только первичноводным позвоночным, закладываются также в эктодерме. Это — кожные органы чувств, представляющие собой желобки, которые тянутся по бокам головы и вдоль тела; они могут быть прикрыты (или нет — у голокожих) костной чешуёй. Органы боковой линии представляют собой сейсмосенсорную систему; они воспринимают лёгкие движения и колебания воды вблизи от источников колебаний: скорость и направление течений, движений собственного тела и наличие предметов на пути движения животного в воде[3].
Органы вкуса закладываются в энтодерме и воспринимают вкус потребляемой пищи в диапазоне: сладкая, горькая, кислая, солёная. Они располагаются на вкусовых сосочках в пределах ротовой полости.
Органы осязания чувствующих клеток не имеют, а представляют собой разветвлённые в коже нервные окончания, которые и воспринимают предметы среды на ощупь.
Органы выделения
У всех позвоночных выделительные органы представлены почками, предназначенными для выведения из тела излишков воды, минеральных солей и продуктов распада азотистого обмена в виде мочевины или мочевой кислоты и аллантоина. Они имеют мезодермальное происхождение, закладываясь на внешней стенке сомитов. Однако строение и механизм функционирования почек у разных групп позвоночных не одинаковы. В процессе эволюции позвоночных животных происходит смена трёх типов почек: головная, или предпочка (пронефрос), туловищная, или первичная, почка (мезонефрос) и тазовая, или вторичная, почка (метанефрос). Разные типы почек имеют разные принципы процесса выделения: выделение из полости тела, смешанное выделение (из полости тела и из крови) и, наконец, только из крови. Одновременно происходят изменения в механизме реабсорбции воды. Выделение воды и растворённых в ней продуктов белкового обмена из полости тела оказывается возможным благодаря множеству воронковидных нефростом, открывающихся в полость тела. Выделение из крови происходит через мальпигиевы тельца почек. Первичные выводные протоки называются вольфовы каналы, в тазовой почке их сменяют вторичные мочеточники. У большинства позвоночных есть мочевой пузырь. У первичноводных возможно сбрасывание аммиака через жабры.
Половые органы
Половые железы позвоночных — яичники у самок и семенники у самцов — как правило, парные. Эмбрионально они развиваются из отдела мезодермы в месте подразделения этого зачатка на сомит и боковую пластинку.
Первоначально (у бесчелюстных) половые железы не имели выводных протоков и половые продукты выпадали через разрывы стенок гонад в полость тела, откуда выводились в наружную среду через специальные поры. Впоследствии возникли половые пути, которые у самцов связаны с выделительными органами (вольфов канал). А у самок в качестве яйцевода функционирует мюллеров канал, который сохраняет связь целома с внешней средой.
Гермафродитизм у позвоночных встречается редко и присущ лишь немногим видам низших позвоночных; все высшие представители позвоночных раздельнополы[11].
Эмбриология
Для позвоночных (как и для других вторичноротых) характерны радиальное дробление оплодотворённого яйца, недетерминированное развитие бластомеров и энтероцельный способ формирования целома, тогда как у прогрессивных сегментированных беспозвоночных дробление спиральное, развитие детерминированное, а целом образуется схизоцельным путём[4].
Генетика
Сравнение геномов позвоночных с геномами оболочников и ланцетника Branchiostoma floridae показало, что у позвоночных имеются следы полногеномных дупликаций, то есть удвоений генома. У бесчелюстных (миног и миксин) выявлена лишь одна дупликация, а у челюстноротых произошла и вторая. В некоторых группах (у костистых рыб, части лягушек) имели место дополнительные полногеномные дупликации. В результате дупликаций увеличилось количество генетического материала, так например, в геноме мыши выявлено 4 кластера Hox-генов вместо одного. И хотя часть новообретённых генов-паралогов впоследствии была утрачена, оставшиеся в процессе эволюции изменились и приобрели новые функции. В основном эти функции связаны с регуляцией активности других генов, регуляцией эмбрионального развития, деятельностью нервной системы и передачей сигналов. Таким образом, полногеномные дупликации обеспечили возможность усложнения организации позвоночных[12][13].
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Родственные связи базальных позвоночных согласно Miyashita et al., 2019[14] |
Происхождение и эволюция

Позвоночные появились в нижнем кембрии[15], а в юре существовали уже представители всех известных ныне их традиционных классов. Общее число современных видов около 40 тыс.
О начальных этапах эволюции позвоночных известно мало. И первые их формы, и их непосредственные предки почти наверняка были мягкотелыми организмами, похожими на современных ланцетников и практически неспособными сохраняться в ископаемом состоянии. Такая возможность возникла только после появления костей, чешуй и зубов. По всей видимости, предпосылки для фоссилизации костей сложились значительно позже первоначальной радиации позвоночных[16].
В 2014 году в силурийских отложениях древностью не менее 423 млн лет найдена Megamastax amblyodus — костная рыба длиной до метра; на 2014 год она считается первым позвоночным хищником, специализирующимся на поедании других позвоночных[17].
Классификация
Эволюционная таксономия

В рамках эволюционной таксономии обычно признаётся от 7 до 9 современных классов позвоночных. Подобные системы классификации часто используются в научных справочниках, а также в школьных и университетских учебниках.
| Традиционная классификация (по А. Ш. Ромеру)[19][20] | Классификация по МСОП[21] |
|---|---|
|
|
Филогенетическая систематика
Проблематично, что эволюционная таксономия основывается прежде всего на морфологических признаках, а не на непосредственном эволюционном родстве. Многие из традиционных классов являются парафилетическими, то есть включают только часть потомков общего предка, тогда как остальные потомки выделяются в более «продвинутые» классы. В настоящее время многие специалисты, особенно палеонтологи, считают, что описанный подход является эволюционно некорректным и поэтому стремятся избавиться от каких бы то ни было парафилетических таксонов[22]. Филогенетические взаимоотношения между современными позвоночными можно отобразить следующей кладограммой[23][24][25]:
| Позвоночные |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| (Vertebrata) |

Избавление от парафилетических таксонов часто сопровождается отказом от понятия таксономического ранга как такового, поскольку ранг таксона зависит от субъективного мнения автора. С другой стороны, в некоторых источниках были предприняты попытки совместить принципы филогенетической и традиционной таксономии. Так, сайт The Taxonomicon придерживается следующей классификации позвоночных[26]:
- Круглоротые (Cyclostomata): миксины и миноги
- † Конодонты (Conodonta). По строению тела близки к круглоротым, однако имели более сложное строение ротовой полости.
- † Парноноздрёвые (Pteraspidomorpha)
- † Анаспиды (Anaspida)
- † Телодонты (Thelodonti)
- † Непарноноздрёвые (Cephalaspidomorphi)
- Надкласс Челюстноротые (Gnathostomata)
- † Плакодермы (Placodermi)
- Хрящевые рыбы (Chondrichthyes)
- Клада Костные позвоночные (Euteleostomi)
- Лучепёрые рыбы (Actinopterygii)
- Клада Мясистолопастные (Sarcopterygii): латимерии, двоякодышащие, четвероногие и их вымершие родственники
- Отряд Целакантообразные (Coelacanthiformes)
- Отряд † Onychodontiformes
- Клада Рипидистии (Rhipidistia)
- Клада Дипноморфы (Dipnomorpha): двоякодышащие и их вымершие родственники
- Клада Тетраподоморфы (Tetrapodomorpha)
- Клада Четвероногие (Tetrapoda)
- Земноводные (Amphibia)
- Клада Рептилиоморфы (Reptiliomorpha): амниоты и их вымершие родственники
- Клада Амниоты (Amniota)
- Клада Синапсиды (Synapsida): млекопитающие и их вымершие родственники
- Млекопитающие (Mammalia)
- Рептилии (Reptilia): черепахи, крокодилы, птицы, клювоголовые, чешуйчатые и их вымершие родственники
- Клада Синапсиды (Synapsida): млекопитающие и их вымершие родственники
- Клада Амниоты (Amniota)
- Клада Четвероногие (Tetrapoda)
Примечания
- ↑ 1 2 Наумов, 1982, с. 24.
- ↑ 1 2 3 4 5 Медников, 1994, с. 257.
- ↑ 1 2 Кэрролл, т. 1, 1992, с. 34.
- ↑ 1 2 3 Медников, 1994, с. 258.
- ↑ 1 2 Наумов, 1982, с. 26.
- ↑ Медников, 1994, с. 257—258.
- ↑ Наумов, 1982, с. 28—29.
- ↑ Зильбернагль, Деспопулос, 2013, с. 130—131.
- ↑ Campbell, 2011, p. 925.
- ↑ Наумов, 1982, с. 41.
- ↑ Марков А. В., 2008.
- ↑ Марков А. В., 2013.
- ↑ Miyashita T., Coates M. I., Farrar R., Larson P.[англ.], Manning P. L., Wogelius R. A., Edwards N. P., Anné J., Bergmann U., Palmer A. R., Currie P. J. Hagfish from the Cretaceous Tethys Sea and a reconciliation of the morphological–molecular conflict in early vertebrate phylogeny (англ.) // PNAS : journal. — 2019. — Vol. 116, iss. 6. — P. 2146—2151. — ISSN 1091-6490. — doi:10.1073/pnas.1814794116. Архивировано 31 декабря 2021 года. — .
- ↑ Shu D.-G., Luo H.-L., Conway Morris S., Zhang X.-L., Hu S.-X. Lower Cambrian vertebrates from south China (англ.) // Nature : journal. — 1999. — Vol. 402, iss. 6757. — P. 42—46. — ISSN 1476-4687. — doi:10.1038/46965. Архивировано 4 августа 2021 года. — .
- ↑ Кэрролл, т. 1, 1992, с. 25.
- ↑ В силурийских отложениях Китая найден древнейший позвоночный хищник. Дата обращения: 10 июля 2014. Архивировано 26 июня 2014 года.
- ↑ Evolutionary systematics: Spindle Diagrams (англ.). Palaeos[англ.]. Дата обращения: 5 июля 2021. Архивировано 25 февраля 2021 года.
- ↑ Romer A. S.[англ.]*. The Vertebrate Body. — Philadelphia: W.B. Saunders[англ.], 1949. (2nd ed. 1955; 3rd ed. 1962; 4th ed. 1970).
- ↑ Позвоночные : [арх. 21 октября 2022] / Ананьева Н. Б. // Большая российская энциклопедия : [в 35 т.] / гл. ред. Ю. С. Осипов. — М. : Большая российская энциклопедия, 2004—2017.
- ↑ Chordata - Phylum (англ.). IUCN Red List of Threatened Species. Дата обращения: 4 августа 2021. Архивировано 4 августа 2021 года.
- ↑ Обширный список литературы см. на сайте Международного общества филогенетической номенклатуры[англ.][1] Архивная копия от 3 августа 2021 на Wayback Machine (англ.).
- ↑ Marjanović D. The Making of Calibration Sausage Exemplified by Recalibrating the Transcriptomic Timetree of Jawed Vertebrates (англ.) // Frontiers in Genetics : journal. — 2021. — Vol. 12. — ISSN 1664-8021. — doi:10.3389/fgene.2021.521693. — PMID 34054911. Архивировано 10 июля 2021 года. — .
- ↑ Betancur-R R., Wiley E. O., Arratia G., Acero A., Bailly N., Miya M., Lecointre G., Ortí G. Phylogenetic classification of bony fishes (англ.) // BMC Evolutionary Biology[англ.] : journal. — 2017. — Vol. 17, iss. 1. — P. 162. — ISSN 1471-2148. — doi:10.1186/s12862-017-0958-3. — PMID 28683774. Архивировано 19 марта 2021 года. — .
- ↑ Chakra M. A., Hall B. K., Stone J. R. Using information in taxonomists’ heads to resolve hagfish and lamprey relationships and recapitulate craniate–vertebrate phylogenetic history (англ.) // Historical Biology[англ.] : journal. — 2014. — Vol. 26, iss. 5. — P. 652—660. — ISSN 0891-2963. — doi:10.1080/08912963.2013.825792. Архивировано 4 августа 2021 года. — .
- ↑ Subphylum Vertebrata (англ.) на сайте Taxonomicon.
Литература
- Биологический энциклопедический словарь / Под ред. М. С. Гилярова и др. 2-е изд. — М.: Сов. энциклопедия, 1989. — 864 с. — ISBN 5-85270-002-9.
- Зильбернагль С., Деспопулос А. . Наглядная физиология. — М.: БИНОМ. Лаборатория знаний, 2013. — 408 с. — ISBN 978-5-94774-385-2.
- Константинов В. М., Наумов С. П., Шаталова С. П. Зоология позвоночных. — М.: Академия, 2000. — 496 с. — ISBN 5-7695-0319-X.
- Кэрролл Р. Палеонтология и эволюция позвоночных: В 3 т. Т. 1. — М.: Мир, 1992. — 280 с. — ISBN 5-03-001819-0.
- Кэрролл Р. Палеонтология и эволюция позвоночных: В 3 т. Т. 3. — М.: Мир, 1993. — 312 с. — ISBN 5-03-001819-0.
- Медников Б. М. Биология: формы и уровни жизни. — М.: Просвещение, 1994. — 415 с. — ISBN 5-09-004384-1.
- Наумов С. П. Зоология позвоночных. — М.: Просвещение, 1982. — 464 с.
- Campbell N. A., Reece J. B., Urry L. A. e. a. . Biology. 9th ed. — Benjamin Cummings, 2011. — 1263 p. — ISBN 978-0-321-55823-7.
- Nelson J. S. Fishes of the World. 4th ed. — New York: John Wiley & Sons, 2006. — xx + 601 p. — ISBN 978-0-471-25031-9.
Ссылки
- Позвоночные — статья из Большой советской энциклопедии.
- Позвоночные животные России // Институт Проблем Экологии и Эволюции имени А. Н. Северцова — Информационная поисковая система по биоразнообразию позвоночных России
- Александр Марков. Геном ланцетника помог раскрыть секрет эволюционного успеха позвоночных. Элементы (23 июня 2008). Дата обращения: 24 июня 2013. Архивировано 29 июня 2013 года.
- Александр Марков. Своим сердцем позвоночные обязаны полногеномной дупликации. Элементы (7 июня 2013). Дата обращения: 24 июня 2013. Архивировано 29 июня 2013 года.
| Таксономия | ||||
|---|---|---|---|---|